极端高温环境对骏枣和伏脆蜜枣光合特性的影响
2023-07-28杨晓娟樊丁宇耿文娟
杨晓娟,靳 娟,樊丁宇,郝 庆,杨 磊,耿文娟
(1.新疆农业大学园艺学院,乌鲁木齐 830052;2.新疆农业科学院园艺作物研究所/农业部新疆地区果树科学观测实验站,乌鲁木齐 830091)
0 引 言
【研究意义】枣(ZiziphusjujubaMill.)原产于中国[1],枣树抗逆性强,适应性广[2],近20年,新疆枣树面积增长速度较快[3]。2020年新疆红枣种植面积为4.67×105hm2,新疆红枣产量约占全国总产量的49.09%,产量达3.728×106t[4]。新疆属于温带大陆性气候,光照、昼夜温差等最适红枣栽培[5],目前极端气候时有发生[6]。新疆2015年夏季极端高温导致红枣大量落果,造成大幅减产[7]。研究极端高温环境对枣树光合特性的影响,对新疆红枣抗逆高产高效栽培具有指导意义。【前人研究进展】气温升高伴随干旱胁迫导致净光合速率和水分利用率下降,增温促进叶片蒸腾速率,也增加了气孔导度和胞间CO2浓度[8]。托克逊灰枣在日平均气温23℃时有利于叶片光合作用,灰枣叶片Pn呈“双峰”型曲线;当日平均气温超过35℃时,影响叶片气孔导度和水分利用效率,不利于光合作用[9]。高温环境下吐鲁番伏脆蜜枣叶片Pn为“单峰”型,骏枣为“双峰”型,Tr均呈“双峰”型,但伏脆蜜Tr变化更剧烈且无光合午休现象,伏脆密抗高温能力更强[7]。塔里木盆地8个不同品种枣树净光合速率日变化均呈“双峰”型曲线,在2个峰值之间有明显光合午休现象,经聚类分析鸡心枣、圆脆枣、灰枣属于低Pn、高WUE、低Tr品种,更适宜环塔里木盆地干旱缺水的环境[10]。【本研究切入点】高温主要降低水分利用效率使净光合速率下降。处于高温环境下的骏枣和伏脆蜜在表型上差异明显,但在持续高温胁迫下对二者在光合生理方面的影响是否存在差异,目前还未见文献报道。需研究极端高温环境对骏枣和伏脆蜜枣光合特性的影响。【拟解决的关键问题】测定持续高温胁迫下两枣品种新梢叶片和两年生枝条叶片光合特性日变化及周变化情况,分析其响应高温环境的光合差异,研究高温对枣树光合特性的影响规律,分析枣树对高温生长环境适应特性,以及新疆红枣耐高温品种选育及栽培提供实践指导。
1 材料与方法
1.1 材 料
试验于2021年6月23~30日在新疆农科院园艺作物研究所人工温室进行,室内温度根据吐鲁番夏季高温环境设定。供试材料为2年生的骏枣和伏脆蜜嫁接盆栽苗,栽植容器为10 L塑料花盆,管理水平良好,树高80 cm,冠幅50~75 cm,树干周长6 cm。高温胁迫期间,每天固定20:00浇400 mL水以防止水分胁迫,每隔1 d每盆浇200 mL的1/2霍格兰营养液。温室内安装智能温度控制器连接加热器,将温度控制在(42±2)℃,并在室内每个培养架上连接日光灯和植物培养专用灯。
1.2 方 法
1.2.1 光合作用测定
仪器为Li-6400XT便携式光合作用仪(Li Cor,Lincoln,NE,USA),测定期间以高温前1 d为对照,持续高温胁迫7 d,分别测定CK、高温1、2、3、5和7 d胁迫的光合日变化,测定时间为每天08:00~20:00,每隔3 h测定环境因子和2品种相同部位功能叶片光合参数,共测定5个时间点。2品种各选生长势一致、树势中庸的样树5株,每株样树同时分别选择新梢和两年生枝条上3片位于中部外侧枣吊上第2~5位具有成熟功能且向光的叶片作为样叶,每片样叶测3次瞬时光合特性参数,取其平均值为测定值。利用Li-6400XT便携式光合作用仪(Li Cor,Lincoln,NE,USA),测时叶室PAR设定为枣的光饱和点平均值1 520 μmol/(m2·s)。测定指标包括净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、基于叶温的蒸气压亏缺(VpdL)、基于气温的蒸气压亏缺(VpdA)、空气温度(Ta)和空气湿度(RH)等,并计算水分利用效率(WUE=Pn/Tr)、气孔限制值(Ls=1-Ci/Ca)等。
1.2.2 温室微气候观测
试验期间采用路格L95-6温湿度记录仪实时监测温室内的气温(Ta,℃)、空气相对湿度(RH,%)。每隔10 min测定并存储1次值。
1.3 数据处理
光合速率数据取45个重复的平均值。应用Excel软件整理数据并绘制图表;利用SPSS26.0软件进行单因素多重比较及相关性分析。
2 结果与分析
2.1 高温处理前后培养间温湿度日变化
研究表明,培养间气温Ta(℃)日变化曲线呈逐渐递增趋势;处理前最高温32.5℃,处理后最高温40.1℃,两者均在20:00达到最大值。处理前后培养间空气相对湿度RH(%)日变化曲线完全不同;处理前08:00空气相对湿度最大为33.93%,然后呈急剧下降趋势,11:00达到最小值24.88%,随着光照时间延长空气相对湿度不断增大;加温后空气相对湿度在观测时间段里日变化趋势为先下降后上升,再逐渐下降,14:00达到峰值37%,20:00降到最低值37%。图1

图1 高温处理前后培养间温湿度日变化曲线及枣苗生长环境
2.2 持续高温胁迫下光合速率日变化
2.2.1 骏枣2年生枝条叶片光合各参数不同日变化
研究表明,CK组叶片的Pn、WUE、Ls值全天均为正值,且明显高于高温胁迫后的值,胁迫期间WUE和Ls降低,推测高温使叶片气孔关闭,进入气孔CO2减少,同时水分利用效率降低,两者均不能满足光合作用所需,导致叶片光合速率降低。Ta与Ci之间呈极显著正相关,高温降低气孔导度,增加叶片胞间CO2浓度,最终影响叶片光合作用。高温1 d处理时叶片的Pn、WUE、Ls值全天均最小,随着处理天数增加各参数日均值逐渐增大,在高温7 d处理时仍远低于CK值,骏枣叶片响应高温胁迫期间产生少量抗体机制,但高温胁迫仍极大降低了其光合作用。图2,表1

图2 持续高温胁迫骏枣光合各参数不同日变化曲线
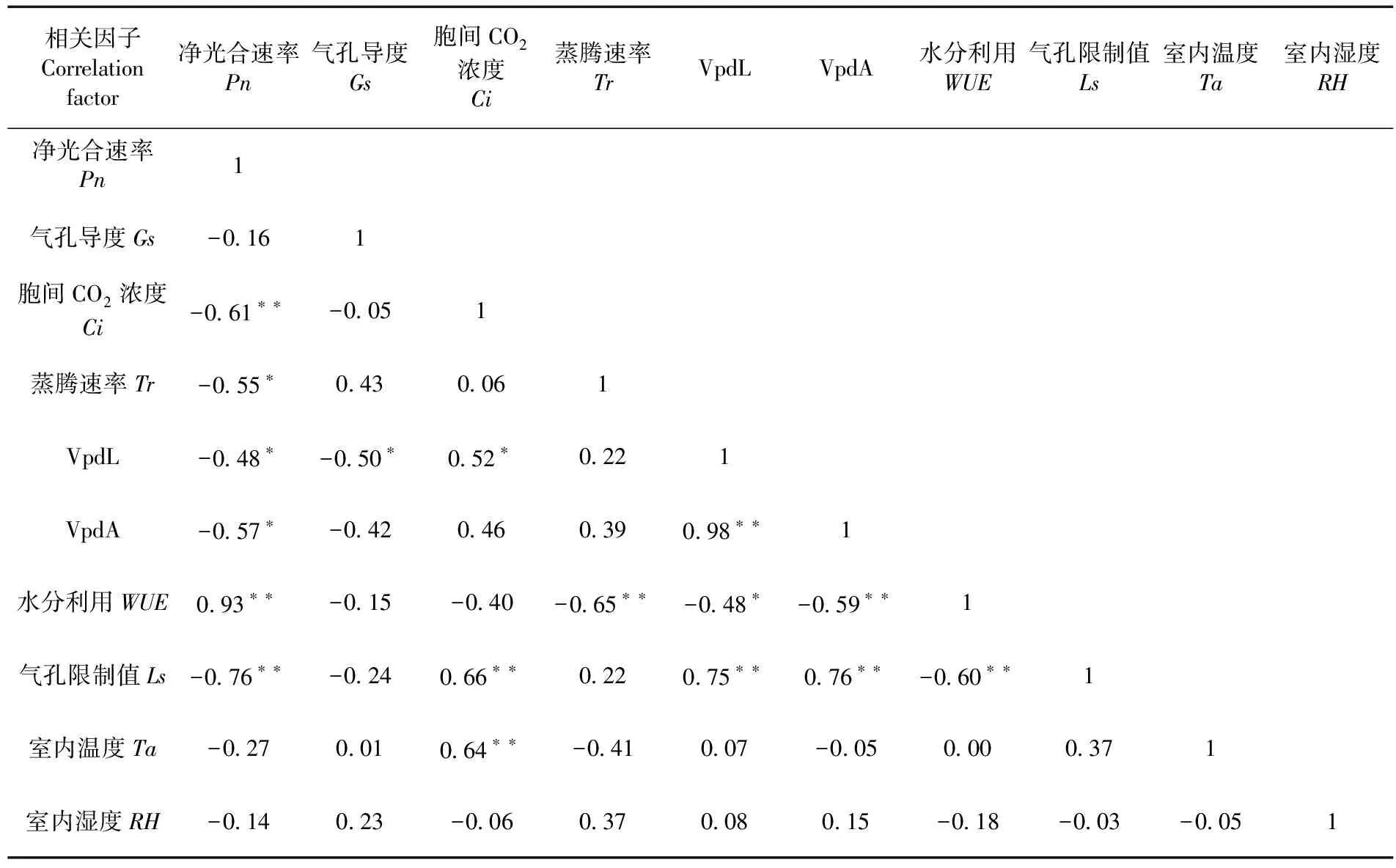
表1 骏枣光合生理因子与生态因子相关性
2.2.2 伏脆蜜枣2年生枝条叶片光合各参数日变化
研究表明,净光合速率是光合和呼吸作用的差值,由于受外部环境以及生理各方面的影响,其在一天内的变化也存在差异。与骏枣相比伏脆蜜叶片Pn在持续7 d高温胁迫期间没有发生明显变化,全天测得CK的Ls均为正值,10:00后高于高温胁迫后的值,持续不同天高温胁迫Ls值全天均呈下降趋势,但下降幅度小、趋势缓慢,与骏枣叶片光合作用降低原因一致,但伏脆蜜气孔没有完全闭合,仍在进行气体交换。同时伏脆蜜树体产生抗体机制更多或缓慢适应高温环境能力更强。各光合参数间相关性表明伏脆蜜Pn与Tr呈极显著正相关(P<0.01),Pn与Ci呈极显著负相关(P<0.01),叶片蒸腾作用越强净光合速率越高,同化CO2能力越强胞间CO2浓度越小,高温环境中伏脆蜜气孔没有完全闭合,叶片通过气孔进行蒸腾作用,带走一部分热量从而降低树体温度,减小了高温对伏脆蜜叶片光合作用的影响。图3,表2

图3 持续高温胁迫伏脆蜜光合各参数日变化曲线
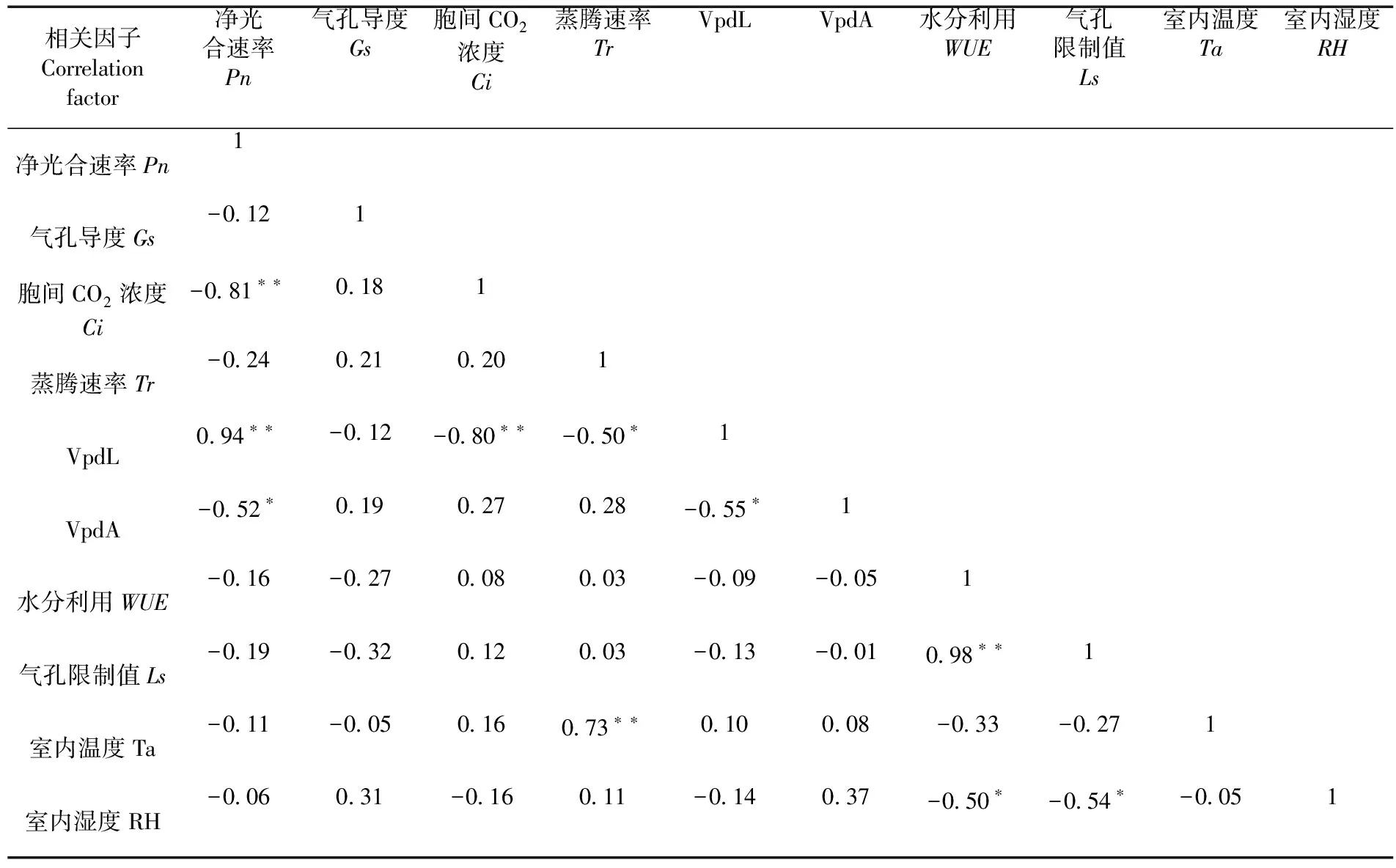
表2 伏脆蜜光合生理因子与生态因子相关性
2.3 持续高温胁迫骏枣和伏脆蜜枣2年生枝条叶片各光合参数周变化对比
研究表明,高温胁迫期间伏脆蜜叶片Ci始终低于骏枣,气孔导度大于骏枣,伏脆蜜叶片气孔没有完全闭合仍在进行气体交换,CO2利用率高,同时水分利用率也相对较高,满足光合所需,使伏脆蜜光合作用强于骏枣。在没有高温胁迫时2品种枣叶片Tr相等且最小,随着高温胁迫天数增加2品种枣变化曲线均呈双峰型,且伏脆蜜始终高于骏枣,相对较高的蒸腾作用带走部分热量使树体温度降低,使伏脆蜜抗高温能力更强,更能适应高温环境。2品种在相同的栽培管理条件下光合作用对其产生的影响大小。连续高温胁迫对2个枣品种Pn、Tr和WUE均有显著影响(P<0.05),对伏脆蜜的Ci有显著影响,但对骏枣的Ci却无显著性影响,高温使伏脆蜜气孔导度相对增大,进入细胞的CO2增多,伏脆蜜光合作用强于骏枣。图4,表3
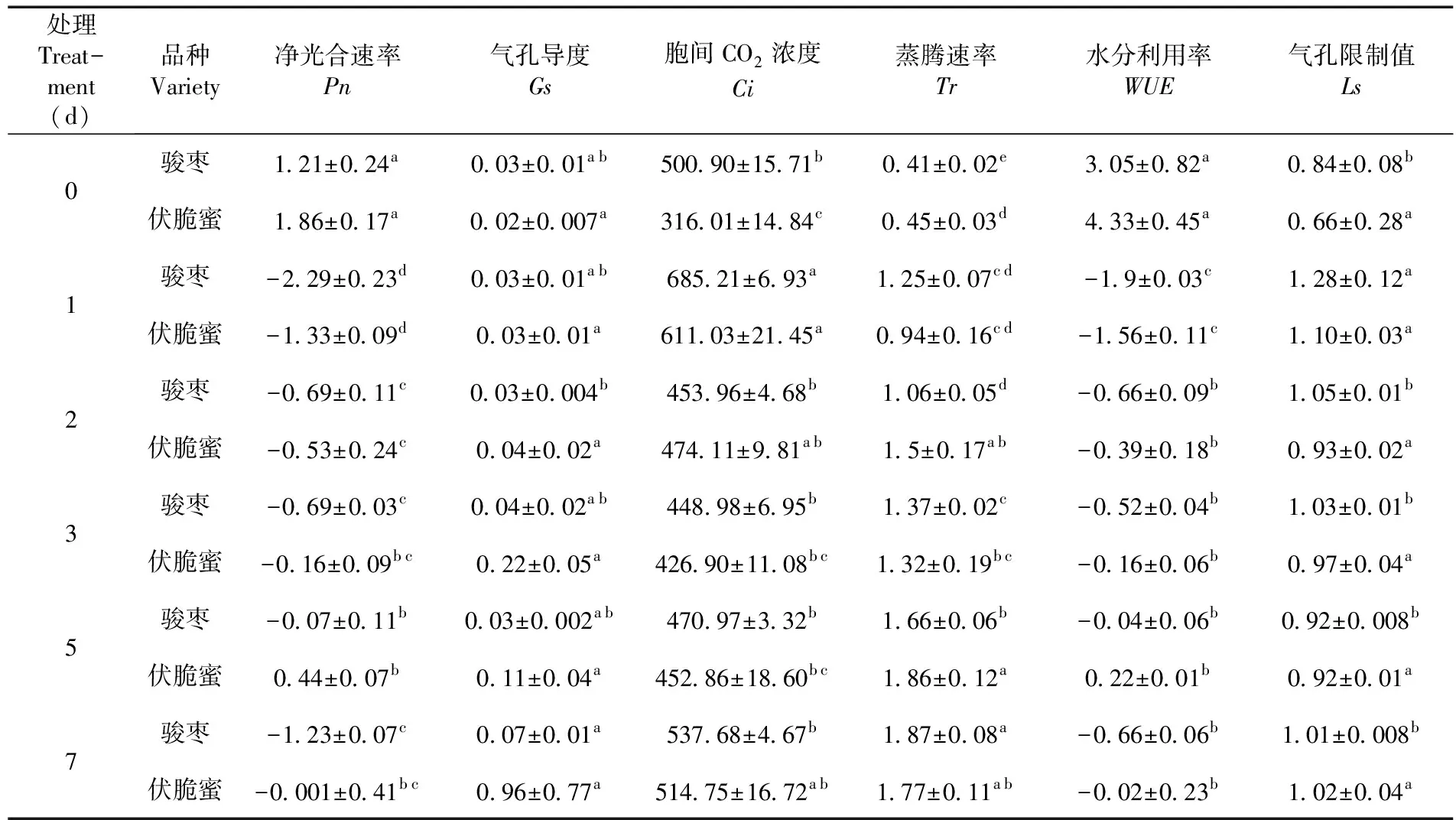
表3 高温胁迫2个枣品种2年生枝条叶片各光合参数日均值

图4 持续高温胁迫两枣品种光合各参数周变化曲线
2.4 持续高温胁迫骏枣和伏脆蜜枣新梢叶片与2年生枝条叶片光合各参数周变化对比
研究表明,高温1 d胁迫骏枣和伏脆蜜不同枝条部位的叶片Pn急剧下降,之后均呈双峰型缓慢上升,在高温7 d胁迫时骏枣不同部位叶片均远低于CK值,而伏脆蜜接近CK值均为正值且新梢叶片高于2年生的,高温胁迫严重影响骏枣叶片的光合作用,且对不同枝条部位叶片的光合速率均有影响,但对伏脆蜜叶片光合作用影响较小,随着高温胁迫天数增加伏脆蜜适应高温胁迫的能力增强,光合作用也增强。高温1 d胁迫时2品种枣叶片WUE均急剧下降,之后呈双峰型缓慢上升,在高温7 d胁迫时仍远低于CK值,高温使水分利用效率降低是引起2品种枣叶片光合作用下降的主要原因之一。伏脆蜜新梢叶片比2年生枝条叶片光合作用强。图5、图6

图5 持续高温胁迫骏枣新梢与2年生枝条光合各参数周变化曲线

图6 持续高温胁迫伏脆蜜新梢与2年生枝条光合各参数周变化曲线
2.5 持续高温胁迫骏枣和伏脆蜜新梢叶片各光合参数周变化对比
研究表明,与老叶相比新叶新陈代谢快,生命力旺盛,高温1 d胁迫骏枣和伏脆蜜叶片Ci、Pn、Tr和WUE值都急剧变化,其中Pn和WUE值都急剧下降,之后Pn、Tr和WUE均呈双峰型缓慢上升,但在整个高温胁迫期间伏脆蜜始终高于骏枣。水分利用效率是光合和蒸腾作用的共同反应,WUE值越大,固定单位数量CO2所需水量越少,水分利用效率越高,高温使叶片水分利用效率迅速降低,不能满足光合作用所需,引起叶片光合作用急剧下降。但在相同水分条件下,WUE值越大,植物抗性越强,伏脆蜜新梢叶片的抗高温能力更强,更能适应高温环境。图7

图7 持续高温胁迫2枣品种新稍光合各参数周变化曲线
3 讨 论
受植物本身的遗传特性影响,同一树种不同品种因其自身的叶片结构、厚度和叶绿素含量不同其光合能力各异[10]。不同枝条部位叶片形成时的环境条件和生理基础不同,光合强度也存在差异[11]。高温降低植物气孔导度、关键光合功能因子,使植物光合产物运输和分配发生变化,叶片光系统PSⅡ受损,最终导致植物体内能量代谢和物质转化过程失衡[12],高温逆境对果树光合作用伤害较显著[13]。当高温、强光照时,空气湿度降低会使植物蒸腾作用加大,根系从土壤中获取水分维持生命活动,若土壤水分充足,Tr与空气蒸发能力呈正比;若土壤水分不足,较大的蒸腾作用导致植物失水严重,空气与叶片之间形成较大水汽压差,保卫细胞失水关闭,因而以气孔限制方式减少蒸腾失水[14]。试验发现,经高温处理都会导致枣树Tr增大,使树体失水加快,在Pn降低的同时,Ci上升或不变,Ls降低,骏枣和伏脆蜜在高温胁迫时出现光合“午休”现象都是由非气孔限制因素导致,与张燕林[15]、徐斌[8]、王龙[16]、刁凯[9]等研究结果一致,但因试验是在人工温室中进行,环境等因素与外界存在差异,所以光合午休现象发生时间有所不同,在连续高温环境中2品种枣树持续处于光合午休状态,净光合速率始终为负数。在塔里木盆地大气温度达到25.6~39.1℃的高温强光条件下,枣树采取了较高的Tr来降低其叶片温度,以避免高温强光对自身的伤害,是红枣对高温干旱环境的适应方式[17]。试验与万素梅等[17]研究结果相同,伏脆蜜在高温处理后Pn、Gs、Tr、WUE和Ls明显高于骏枣,推测伏脆蜜抗高温能力更强。
植物光合作用不仅受生态因子影响,也受其生理因子影响[18],如Ta、RH、PAR等环境因子能引起Tr、Pn、Ci、Gs、Tr等生理因子变化,最终影响植物光合作用[19]。试验结果表明,骏枣的Pn与WUE呈极显著正相关,WUE在骏枣光合作用过程中起了非常重要的作用,可以防止由于失水过多对树体造成损伤。Pn与Ci、Ls呈极显著负相关,Pn与Tr、VpdL、VpdA呈显著负相关,其余等指标的相关性都未达到显著水平,Ta与Ci之间呈极显著正相关,高温影响气孔限制值,从而增加叶片胞间CO2浓度,最终影响叶片光合作用,Ci、WUE和Ls对骏枣Pn影响最明显。伏脆蜜Pn与Tr呈极显著正相关,Tr在其光合作用过程中起了非常重要的作用,可以防止由于叶温过高对叶片造成损伤,而Pn与Ci呈极显著负相关,Pn与VpdA呈显著负相关,其余等指标相关性都未达到显著水平,Ta与Tr之间呈极显著正相关,高温没有使气孔闭合,增大了叶片的蒸腾速率,降低了高温对叶片的伤害,最终影响伏脆蜜叶片光合作用,而Ci和VpdL对伏脆蜜的Pn的影响最为明显,生理因子均对2品种枣叶片Pn影响较大,而生态因子对其影响次之,与王振东[20]、何永波[21]等研究结果稍有出入。试验同时也研究了2个品种枣的新梢叶片和2年生枝条叶片的各光和参数周变化,结果显示在整个测定期间骏枣新梢叶片和2年生枝条叶片的各光和参数周变化区别不大,而伏脆蜜的新梢叶片比2年生枝条叶片光合作用强,且高温胁迫1 d时两者各光和参数都发生了巨大变化。当室内温度过高,引起气孔导度和蒸腾速率变化,最终影响枣树光合速率的变化[22]。叶片衰老是一个遗传程序化的发育过程,其衰老的时间与植株的衰老是不同步的,同一株植物的新叶与老叶在不同的胁迫条件下展现出不同的抵御方式,新叶与老叶在同一植株中,二者之间存在密不可分的联系。同一个体的新叶比老叶有更多的遗传物质,老叶积累化学代谢物,以保护植物抵御逆境,但由于新叶需要生长,将资源分配给生长而不是防御[23],与试验研究结果一致,所以伏脆蜜枣表现出新梢叶片光合作用强于2年生枝条叶片。试验只模拟了高温生长季节时的环境,并对此高温期间枣树光合变化规律进行了研究,而对其前期生长季节的低温环境中的光合特性未做研究,需今后进一步研究。
4 结 论
高温使骏枣叶片气孔关闭,细胞内外气体交换减少,进入细胞CO2减少,同时水分利用效率降低,两者均不能满足光合作用所需,导致叶片光合速率降低;伏脆蜜枣叶片Pn在持续7 d高温胁迫期间没有发生明显变化,且Pn与Tr呈极显著正相关(P<0.01),Pn与Ci呈极显著负相关(P<0.01),气孔没有完全闭合,仍在进行气体交换,叶片通过气孔进行蒸腾作用,降低树体温度,从而减小了高温对伏脆蜜枣叶片光合作用的影响。持续高温胁迫期间伏脆蜜枣不同枝条部位叶片的光合作用周变化始终高于骏枣,伏脆蜜枣适应高温胁迫的能力更强,并伏脆蜜枣的新梢比2年生枝条叶片光合作用强,伏脆蜜枣新梢叶片适应高温胁迫的能力最强。
