新源野果林3种野生果树春播成苗特性分析
2023-07-28刘立强尤璐瑶曼苏尔那斯尔
程 功,李 悦,刘立强,尤璐瑶,曼苏尔·那斯尔,廖 康
(1.新疆农业大学林学与园艺学院,乌鲁木齐 830052;2.邯郸市农业科学院,河北邯郸 056000)
0 引 言
【研究意义】新疆天山野果林是中亚野果林的重要组成部分,是野苹果(Malussieversii)、野杏(Armeniacavulgaris)、野樱桃李(Prunusdivaricata)等多种野生果树的起源地[1, 2],为改良培育抗逆性强的果树新品种提供丰富的遗传基因资源。近年来病虫害蔓延,严重威胁到野果林的生存环境,致使野果林树木死亡,野生果树自然繁衍受阻,植物基因库受损严重。野苹果、野杏和野生樱桃李是野果林的重要组成树种,其繁殖更新能力对维持野果林的种群可持续发展有重要作用。研究野生果树的自然繁殖特性,对探索其实生更能力和研究促进野生果树的更新措施有重要意义。【前人研究进展】野苹果主要通过种子萌发和根蘖苗繁殖更新种群[3]。野杏主要依靠种子繁殖更新,开花期和其它果树相比较早,极易受到晚霜和春雪伤害,在一定程度上造成更新的种源限制,天然更新能力降低。对野苹果[4]和野杏[5]的调查发现林下幼苗数量少,抗性差,生长量小,容易枯死,天然更新困难。野生樱桃李可通过种子萌发实现种群的更新,但种子在原生境萌发具有较大的风险,萌生苗比实生苗对于更新的作用更大[6]。对野生樱桃李的天然更新调查发现,果实成熟离开母株后,由于土壤枯落物层的存在,阻碍了种子与土壤的有效接触,动物取食搬运和风等物理因素同样影响天然更新[7]。对森林的人工更新以飞机播种造林更新技术为主,已经成为森林恢复的主要途径[8],但飞播耗种量较大,林下幼苗影响因素多,成苗困难,造林效果差[9]。【本研究切入点】野杏种子的萌发试验表明,种子萌发过程中有吸水特性,低温层积100 d时种子萌发率可达到90%,是春季播种的最佳催芽方式[10, 11],但层积时间不宜超过130 d,而野苹果种子的最佳层积条件为低温层积75 d,发芽率可达到90%[12],野樱桃李的最佳萌发条件为低温层积100 d,其萌发率可达到72%[13]。另外,去除种壳,种皮和使用一定浓度的GA3处理都可以促进3种果树种子的萌发。野樱桃李的扦插繁殖在蛭石和河沙基质中的生根率可达到40%,而使用一定浓度的NAA处理或NAA和IBA(1∶ 1)可促进生根[14, 15]。上述研究都是关于野生果树在室内控制条件下繁殖特性研究,与原生境的繁殖特性和能力还有很大差别。【拟解决的关键问题】将野杏、野樱桃李和野苹果的种子层积后,春播于新源野果林的平地、阴坡和阳坡3种试验地,分析其出苗规律和成苗特性,研究3种果树的天然繁殖能力,为野生果树人工更新措施的研究提供理论依据。
1 材料与方法
1.1 材 料
试验地位于新疆新源县野果林(E 83°36′4.92″,N 43°22′44.28″),海拔1 402.6 m,属于温带大陆性气候。试验地年均气温9.37℃,极端气温分别为-25.42℃(1月28日)和32.65℃(7月28日),平均光合有效辐射为221.1 W/m2,年降水量556.8 mm,年内降水分布不均,其中4~6月降雨占全年降水的49.1%,土壤pH值为7.38,0~50 cm深度土壤年均含水量为0.227 m3/m3,来自自然降水。播种时土壤表层含水量分别为平地0.356 m3/m3,阴坡0.306 m3/m3,阳坡0.256 m3/m3,播种后30 d内共降水7次合计61.6 mm。播种的平地、阴坡是卫生采伐后的野苹果林地,阳坡没有生长野生果树,以禾本科植被为主。
提前1年秋季果实成熟期,采集原生境的野杏、野苹果和野樱桃李成熟果实,水洗净种后,野苹果种子和野杏种子阴干保存,野樱桃李湿纱布包裹后保湿保存。于第2年播种日期(4月初)前90 d将野杏浸种10 h种子充分吸水后装入纱网袋中,放入4℃冰箱层积保湿层积保存,播种前60 d将野苹果种子和野樱桃李种子水中浸种4 h后装入纱网袋放入4℃冰箱保湿层积保存。部分种子提前萌发时可降低2~3℃保存种子。
1.2 方 法
1.2.1 样地选择与播种
在试验区内选择平地(坡度4°)、阴坡(坡向西北,坡度25°)、阳坡(坡向西南,坡度35°)3种类型试验地,1 m×1 m样方,3次重复,待层积种子萌发后,挑选萌芽种子于春季开沟条播于试验地样方中,播种深度为1~2 cm,3种种子萌发率差异较大,每样方播种量为野杏280粒,野樱桃李122粒,野苹果约792粒。
1.2.2 出苗规律及成苗观测
播种后,每3 d记录1次样方中出苗数量,待无幼苗出土时,计算出苗率;待最后一批出土的幼苗生长出4叶1心时,统计苗木数量,记为成苗数[16];待秋季幼苗停止生长后统计所有样方中存活的苗木数量为保存数量[9]。
出苗率=(出苗数/播种数)×100%;
成苗率=(生长至4叶1心幼苗的数量/播种数)×100%;
保存率=(保存数量/成苗数量)×100%。
1.2.3 幼苗生长测定
播种30 d后开始测量株高、基径和节间长,此后每20 d测量1次。
1.2.4 幼苗生物量测定
于9月20日,幼苗开始落叶前,每样方选取5株有代表性,生长健壮的完整幼苗,整株挖回,测定幼苗的株高、主根长、侧根长、侧根数量、根鲜重、根干重,茎鲜重、茎干重、叶鲜重、叶干重。
1.3 数据处理
试验数据采用Sigmaplot 14和Excel 2013作图,SPSS 20.0软件数据分析。
2 结果与分析
2.1 3种野生果树播种出苗规律
研究表明,播种后13 d,苹果在阴坡首先开始出苗,出苗期集中在4月13日至5月1日,出苗数量占总出苗数量的82.2%;野杏和野樱桃李播种后16 d平地首先开始出苗,在4月19日至5月4日出苗数量较多,野杏出苗数量占总出苗数量81.8%,野樱桃李出苗数量占总出苗数量的86.3%。此后出苗速率减慢。
近年来,在云南省各级森林公安机关的共同努力下,打击破坏野生动物资源犯罪的工作卓有成效。但同时,破坏野生动物资源犯罪的案发数仍然居高不下,野生动物非法贸易现象依然猖獗。
3种试验地出苗率均为野杏>野樱桃李>野苹果,野杏出苗率均高于38%,阳坡最高,平地最低,野苹果均低于7%,阴坡出苗率最高,阳坡最低;野樱桃李在3种试验地出苗率分别为平地13.1%,阴坡17.8%,阳坡10.2%,无显著差异。图1
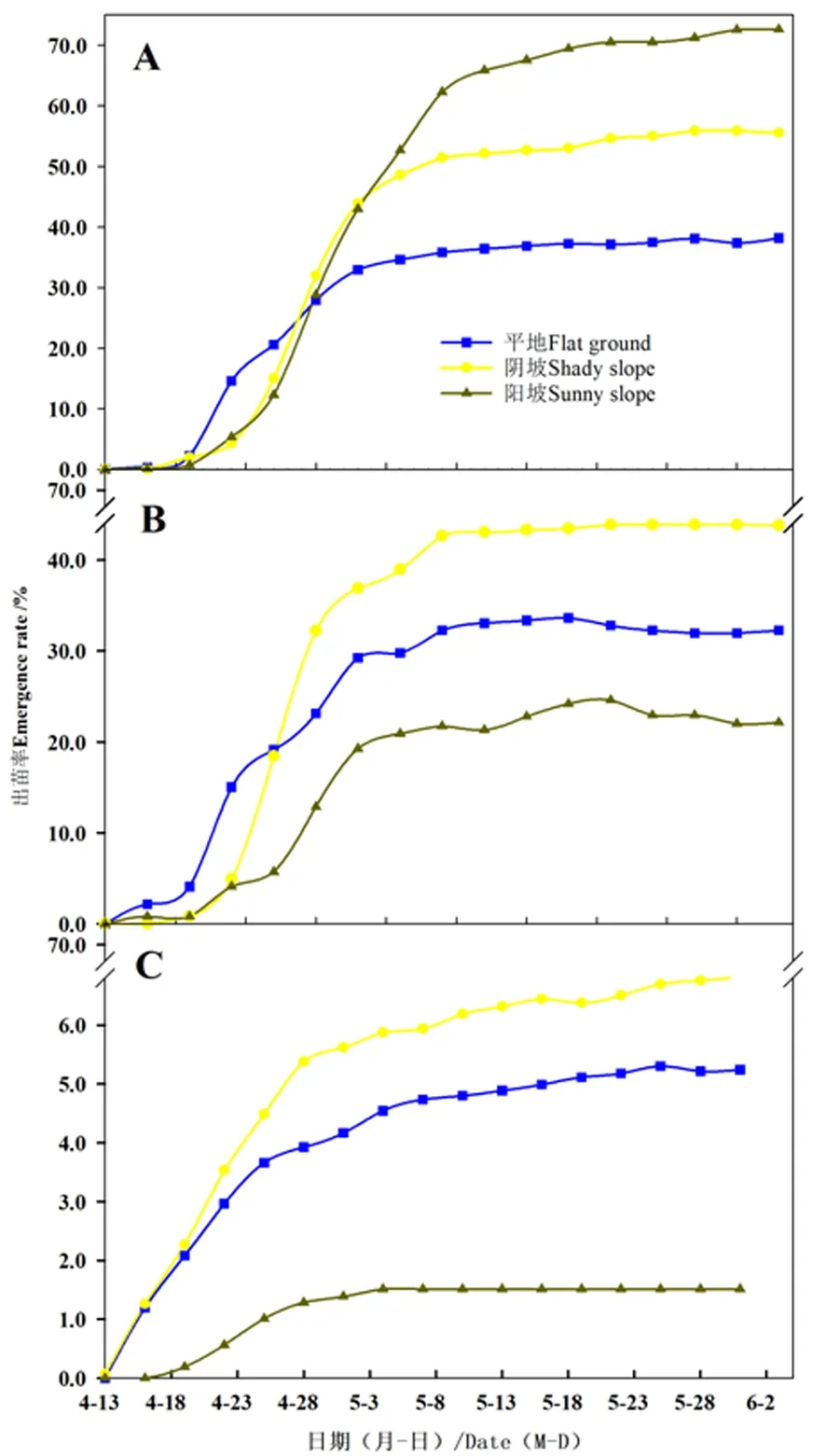
注:A、野杏;B、野樱桃李;C、野苹果
2.2 3种野生果树成苗率及保存率
研究表明,春播后,3种试验地的野杏成苗率均大于38%(阳坡大于72%),保存率均大于93%(阳坡保存率为100%),成苗率和保存率最高;野苹果在阴坡的成苗率最高,为6.8%,在阳坡最低,仅有1.5%,平地和阴坡的保存率为57.4%~60.2%,在阳坡的保存率较高,为83.3%;平地和阴坡的野樱桃李的成苗率可达到37%~48%,而阳坡的成苗率仅有22.1%,而保存率平地和阴坡在95%,阳坡仅有46.3%。
仅有平地和阳坡的野杏成苗数量和成苗率有显著差异,而野樱桃李和野苹果的出苗数量和出苗率均没有显著差异;野杏平地和阳坡的保存数量有显著差异,野樱桃李阴坡的保存率显著高于阳坡,野苹果的保存数量没有显著差异,3种野生果树在三地的保存率均没有显著差异。表1
2.3 3种野生果树实生苗生的年长动态
研究表明,野杏的株高生长最快的时期是在播种后30~130 d,在此期间3种试验地的野杏平均日生长量分别为平地0.80 cm,阴坡为0.50 cm,阳坡为0.38 cm。而3种试验地的主要株高生长从70 d开始差异逐渐增大,至130 d时差异达到最大。在此期间,3种试验地的平均气温分别为平地18.83℃,阴坡为18.66℃,阳坡为20.61℃,0~20 cm土壤含水量分别为平地0.272 m3/m3,阴坡0.285 m3/m3,阳坡0.219 m3/m3,在130~170 d株高生长缓慢;野杏苗的地径在70 d前时,3种试验地间差异较小,在70 d后地径生长量差异变大,平地的地径最大,地径在整个生长季均逐渐增大;不同生境的野杏苗的节间长变化无显著差异。在生长的同时期,野杏的株高和地径平均生长量均为平地最高,阴坡次之,阳坡最小。
春季播种后,野樱桃李的株高生长变化主要集中在30~130 d,3种试验地的野樱桃李平均日生长量分别为平地0.92 cm,阴坡为0.67 cm,阳坡为0.28 cm,其中从第70 d开始,平地和阴坡的生长速度明显高于阳坡,90 d时,阴坡樱桃李生长变缓,其中70 d平地和阴坡开始快速生长,至130 d时,平地的生长量最大,此后,野樱桃李生长均减缓,在此阶段,平地的平均气温为18.83℃,光照为33 685 lx,土壤含水量为0.272 m3/m3;地径在整个生长季均逐渐变大,3种试验地的地径从第50 d开始差异逐渐变大;节间长变化主要集中在30~130 d,其中阴坡的节间长在大约第100 d时超过平地,成为最大。在生长季中,野樱桃李的株高和地径均为平地最高,阴坡次之,阳坡最小。
春季播种后,野苹果的株高变化集中在30~130 d,在此期间3种试验地的野苹果平均日生长量分别为平地0.17 cm,阴坡为0.08 cm,阳坡为0.12 cm,株高快速生长期集中在70~110 d,平地的气候适宜株高的生长;但在第100 d时,阳坡野苹果的株高超过阴坡;地径在整个生长季均逐渐增大,3种试验地差异较小,至170 d时,阳坡的地径最大;节间长的变化集中在70~130 d,随后变化减小;在整个生长过程中,野苹果的株高和节间长始终为平地最高,3种试验地的地径间的差异较小。图2

表1 3种野生果树春播成苗率及保存率
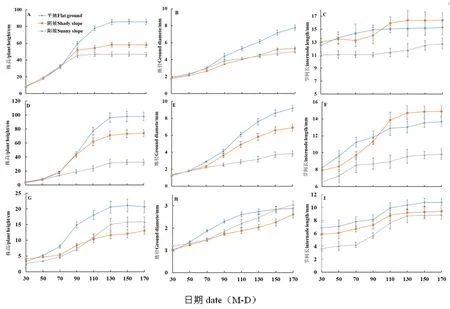
注:A.野杏株高;B.野杏基径;C.野杏节间长;D.野樱桃李株高;E.野樱桃李基径;F.野樱桃李节间长;G.野苹果株高;H.野苹果基径;I.野苹果节间长
2.4 3种野生果树年生物量
研究表明,平地、阴坡和阳坡的1年生野杏苗高分别为85.32、57.93、46.52 cm,3种试验地间均有显著差异。叶片鲜重和干重均为阴坡最大,分别为3.99和1.54 g,3种试验地间差异均不显著。野杏在平地的茎鲜重为17.75 g,干重为9.66 g,分别为3种试验地中最高,平地分别与阴坡和阳坡的茎鲜重与干重有显著差异。野杏的根鲜重平地为15.17 g,干重为7.90 g,均为3种试验地中最重,显著高于阴坡和阳坡。侧根数是阳坡最大,平地次之,阴坡最少,侧根长为平地最大,阳坡次之,阴坡最小,侧根长和侧根数量3种试验地之间没有显著差异。全部生物量鲜重和干重均为平地最大,分别为37.92和18.16 g,均与阴坡和阳坡达到显著差异,但阴坡和阳坡间生物量没有显著差异。图3

野樱桃李在3种试验地的株高分别为97.88、74.04和32.52 cm,平地最高,阳坡最低,平地和阴坡分别与阳坡有显著差异。野樱桃李在平地的叶鲜重和叶干重最大,分别为2.70和0.97 g,叶干重平地与阴坡差异不显著,与阳坡差异显著。樱桃李在平地的茎干鲜重和干重均为3种试验地中最大,为21.41和12.78 g,平地的鲜重与干重同时显著高于阴坡和阳坡,主根长和根鲜重为平地最大,其中主根长3种试验地均为显著差异,根鲜重仅有阴坡与另两地为显著差异,根干重平地最大,为10.03 g,显著高于阴坡和阳坡,侧根数阴坡最大,与平地差异不显著,但与阳坡差异显著,侧根长则为阳坡最大,3种试验地间差异不显著。整株生物量均为平地最大,分别为49.86和25.07 g,同时与阴坡和阳坡有显著差异,但阴坡和阳坡间生物量没有显著差异。
野苹果的株高在3种试验地中分别为20.69、13.14、15.82 cm,平地最高,阴坡最低,平地分别与阴坡和阳坡差异显著。叶片鲜重阳坡最大,为1.35 g,与平地和阴坡有显著差异。叶片干重平地最大,为0.64 g,3种试验地间差异不显著。平地的茎鲜重和干重最大,分别为0.88和0.38 g,但3种试验地间差异不显著。主根长与根鲜重均为阴坡最大,平地最小,3种试验地间差异不显著,根干重为阳坡最大,差异不显著。侧根数量阳坡最大,为52.52条,3种试验地差异不显著,侧根长阴坡最大,为6.62 mm,3种试验地差异不显著。整株生物量的鲜重和干重均为阳坡最大,分别为2.64和1.95 g,但鲜重间没有显著差异,阳坡的干重与平地和阴坡有显著差异。表2
3 讨 论
3.1 3种野生果树的出苗条件
在春季,种子萌发出苗的时间及其气候条件影响成苗质量。野果林的春季气候变化幅度较大,如果比常规物候期提前升温,在满足积温需求条件下种子就会萌发出苗,出苗过早,可能会在后续出现的低温气候下造成幼苗冻害或死亡,同时,春季长时间没有积累够有效的积温,持续低温,则会降低种子的萌发能力,甚至造成种子腐烂降低种子的天然更新能力。因此,保证出苗期适宜的环境条件,种子的集中萌发效应才会凸显,出苗和成苗率才会高,幼苗长势也会较好[17]。原生境下,霍城大西沟的野杏的种子萌发成苗期在3月下旬至5月上旬,而对试验区的环境监测结果表明[18],试验区气温从4月上旬开始上升,地温上升稍有延迟,为种子的萌发提供了适宜的条件,试验中,野杏和野樱桃李的出苗期集中在播种后19~33 d,即4月中旬及下旬,时间跨度小,出苗整齐,长势均较好。野苹果播种量最大,但主要出苗时期超过20 d,出苗少,不整齐,长势也最差。
3.2 3种野生果树的出苗与成苗特征
种子播种后,多种因素共同影响种子和幼苗的命运,包括气温、光照、土壤温湿度、动物取食和病虫侵袭,异质生境的影响、种壳的制约、干扰及繁殖体特征的影响,密度和距离的制约,地形地势的影响[19]等。试验将层积的种子播种至3种试验地后,3种种子成苗率和保存率均不同,3种试验地成苗率和保存率最高的均为野杏,其成苗率均大于38%,在阳坡甚至大于72%,野杏的保存率在3种试验地均大于93%,在阳坡的保存率为100%,这可能与野杏具有较大的种仁,能够为种子萌发生长提供充足的营养物质有关,而野杏播种前具有最高的发芽率(93%),野杏种子具有较高的生活力,适应性强。野苹果的室内育苗试验结果表明,野苹果的种子出苗率为31.8%,而试验3种试验地中野苹果的成苗率仅有7%,可能是因为播种时试验地地温较低,不适宜野苹果种子出苗,而在播种后第2 d(4月2日)的降温可能阻碍了野苹果的出苗。
3.3 3种野生果树一年生苗的生长特征
种子发芽出土后形成幼苗,幼苗期抗性较差,是植物生长的关键时期。对山杏幼苗的研究表明,土壤含水量低于7%时,严重影响山杏的生长,土壤含水量为17%时,山杏的蒸腾速率,光合速率和水分利用效率有最大值[20],而土壤含水量达到最大田间持水量的52%时,才会产生较大的生物量[21]。野樱桃李主要分布于大西沟中常有溪水的支沟中,并与野杏野苹果野山楂等混生成为混交林,而较为干旱的沟中则没有分布[22]。野苹果具有较强的引种适应性[23],能适应较高的土壤含水量[24],但是土壤含水量较低时会影响苹果幼苗的光合速率[25]。试验中,3种野生果树幼苗的生长集中在播种后30~130 d,生长较快,该时期是野果林温度上升,降雨集中的时期,但生长量差异较大,就生长量来看,三者均为平地生长量最大,是因为降雨后平地水分没有流失,有较高的土壤含水量,土质肥沃,为植物生长提供适宜的环境,阴坡次之,阳坡生长量最小,是因为阳坡降雨水分流失,温度高,蒸发量大,土壤含水量低,土质养分少;根据3种试验地类型的幼苗生长量,平地和阴坡适宜春播更新野杏和野樱桃李,而阳坡适宜春播野杏更新,而苹果生长量最小,与杂草竞争力差,春播成苗困难。
3.4 3种野生果树一年实生苗的存活能力
野樱桃李和野苹果在3种试验地的成苗率均为阴坡>平地>阳坡,可能是因为阴坡和平地在种子出苗期有较高的土壤含水量(平地0.353 m3/m3,阴坡0.308 m3/m3),而阳坡更易受风等因素的影响蒸发量较大,土壤含水量低(0.255 m3/m3),平地和阴坡的杂草生长量大,杂草枯落物层厚,也增加了表层土壤的含水量,而阳坡杂草量少,保水效果不明显,但阳坡的土壤含水量适宜野杏的出苗。3种试验地环境的不同导致生长季结束后幼苗保存率不同。在生长过程中,平地和阴坡的野苹果幼苗叶片腐烂严重,幼苗逐渐死亡,可能是因为这两地的杂草生长量大,近地面的空气相对湿度较大,而苹果幼苗生长慢,长期处于较高空气相对湿度中导致腐烂,而这种影响在阳坡则不明显,所以阳坡的野苹果保存率在3种试验地中最高,野杏和野樱桃李生长快,可减小近地表高湿环境的影响。在野果林的人工更新中,野苹果幼苗期应加强锄草防草,降低高湿度环境对野苹果生长的影响。野樱桃李的在3种试验地中的保存率正好相反,阳坡最低,是因为土壤水分含量是影响其生长的关键因素[26],阳坡在降雨后难以存水,蒸发量大,土层含水量低,抑制野樱桃李的生长。
4 结 论
野杏和野樱桃李出苗期集中在播种后19~33 d,野苹果的较早、始于播种后13 d。野杏、野樱桃李和野苹果的成苗率分别为38%~72%、22.1%~41.8%和1.5%~6.8%,存活率分别为93%~100%、46.2%~95%和57.4%~83.3%,野杏的成苗率和存活率均在阳坡最高、后两者的均在阴坡最高。3个树种生长速率和年生长量均在平地最好。原生境中,野杏实生更新适应性最强。
