转cry2A*/bar基因水稻与杂草稻杂交后代的苗期生长特性
2023-07-18肖乐铨李雷戴伟民强胜宋小玲
肖乐铨 李雷 戴伟民 强胜 宋小玲
转基因水稻与杂草稻杂交后代的苗期生长特性
肖乐铨 李雷 戴伟民 强胜 宋小玲*
(南京农业大学, 南京 210095;*通信联系人, email: sxl@njau.edu.com)
【目的】考查转基因水稻T2A-1与杂草稻的杂交后代苗期特性,为评估T2A-1抗性基因向杂草稻漂移可能引起的生态风险提供参考。【方法】在水培条件下,以T2A-1、受体水稻明恢63(MH63)、来源于江苏泰州、广东茂名、湖南益阳的3种杂草稻(分别命名为WRTZ,WRMM,WRYY)及其杂交F1~F4为试验材料(以T2A-1为父本或母本的携带转基因的为抗性杂交后代,以受体水稻为父本或母本的为非抗性杂交后代),测定供试材料种子发芽力,幼苗在正常氮(40 mg/L)和低氮(20 mg/L)下的株高、整株干物质量以及根长、不定根数、平均直径等8个根系指标,计算根系总体发育指标,评价T2A-1与杂草稻抗性杂交后代的苗期生长特性。【结果】抗性杂交后代的发芽力与非抗性杂交后代相当,但较杂草稻弱。在正常氮条件下,T2A-1与MH63以及抗性杂交后代与非抗性杂交后代的苗期生长指标相当;抗性杂交后代与相应杂草稻相比,WRMM杂交后代的株高显著低于WRMM,但整株干物质量与根系总体发育指标相当;WRYY杂交后代各苗期生长指标均与WRYY相当;WRTZ杂交后代株高显著高于WRTZ,但整株干物质量及根系指标均显著低于WRTZ。在低氮环境下,抗性杂交后代各苗期生长指标均显著低于或相当于非抗性杂交后代;抗性和非抗性杂交后代的相对根系发育指标均显著低于相应的杂草稻。【结论】T2A-1与杂草稻的抗性杂交后代的苗期生长特性在正常氮条件下相当于或低于亲本杂草稻,但在低氮条件下,总体上均显著弱于杂草稻。
复合性状转基因水稻;T2A-1;杂草稻;杂交后代;苗期生长特性;根系发育
水稻(L.)作为我国乃至全世界的主要粮食作物,其转基因技术非常成熟。通过该技术已经培育出抗除草剂、抗虫、品质改良的转基因水稻。其中,华中农业大学培育的转基因抗虫水稻“汕优63”和“华恢1号”两次获得我国农业农村部授予的生产安全证书。除单一性状外,还成功培育出多种复合性状转基因水稻,特别是兼具抗除草剂和抗虫性状的转基因水稻[1-3]。其中,华中农业大学培育的转的T2A-1对二化螟、三化螟和稻纵卷叶螟表现出高抗,同时具有很好的抗草胺膦特性,农艺性状与受体明恢63基本一致,具有产业化前景[4-5]。
种植转基因水稻存在潜在的生态风险,其中最值得关注的是抗性基因通过花粉漂移到同属同种的杂草稻(weedy rice)。杂草稻是广布世界水稻种植区的恶性杂草,具有较强的落粒性、休眠以及早熟特性,能逃脱人工收获进入土壤种子库来年继续为害[6-11]。杂草稻种子萌发特性、苗期光合特性,以及对氮素的利用能力均强于栽培稻,在与栽培稻竞争中占据优势地位。一旦转基因水稻通过花粉介导与杂草稻产生杂交后代,其中的抗性杂交后代因无法用目标除草剂防除而有可能成为田间难以防除的杂草[12-13]。但也有学者认为,对于转基因安全评价需要讲究风险效益平衡的原则。商业化种植转基因抗除草剂和抗虫水稻能提高杂草和靶标害虫的防除效果,提高水稻产量。在合理使用除草剂的基础上,抗除草剂水稻技术能为杂草稻的防除提供简单有效的解决方法,同时也防止了外源基因向杂草稻的漂移,获得的效益要远大于风险。因此需要我们加以客观分析和全面评价[14]。
杂草稻与栽培稻的亲缘关系近,亲和性极高,极易杂交,并能产生杂交后代[15-16]。前期研究发现田间T2A-1向供试江苏泰州和湖南益阳的杂草稻发生基因漂移,漂移率为0.1103%~0.2065%[17]。一旦发生基因漂移,这些抗性杂交后代种子的萌发特性以及苗期生存竞争能力对其在自然界生存定植会有重要影响[18-19]。
种子萌发与幼苗的建立是植物生活史中的关键事件,种子萌发速度越快、越早生根和分蘖的幼苗,其不仅有较长的生长时间,且对环境资源的利用能力越强,因而具有竞争优势[20-23]。具有较高活力的水稻种子在田间中能够快速、均匀发芽,在杂草生长的抑制下具有强大的幼苗发育能力[24-25]。因此,发芽快慢是决定幼苗竞争能力的重要因素。杂草稻比栽培水稻提前2~3 d萌发出苗是杂草稻苗期强竞争力的基础[19, 26]。
植物根系是水分和养分吸收的主要器官,其形态和根系活力对土壤养分吸收有重要作用[27]。已有研究表明,相比栽培稻,杂草稻具有更强的根系,因而具有更强的竞争能力[19]。吴云艳[28]研究了杂草稻与栽培稻在盆栽混种条件下两者的根部吸收养分能力,结果表明杂草稻根部吸收养分能力显著强于栽培稻。Schaedler[29]研究在光竞争下杂草稻与栽培稻根系的初期生长情况,结果发现对光的竞争一定程度上抑制了两者的根系发育,但杂草稻根系发育仍然强于栽培稻。
杂草稻较强的根系使得其能够高效利用氮肥。Burgos等[18]发现无氮肥施用时, 杂草稻与栽培稻的生物量相近,而施用氮肥后杂草稻茎秆的生物量迅速增加, 且显著高于栽培稻。由此推测杂草稻具有更高的氮肥利用效率。吴云艳等[28]的研究表明, 杂草稻对氮肥的利用效率高于栽培稻,有些杂草稻对氮的利用甚至比栽培稻高出3倍。
转基因水稻的基因通过花粉漂移到杂草稻后,杂交后代的种子萌发特性和苗期特性与杂草稻相比是否发生变化,在无选择压条件下,抗性基因对杂交后代的苗期特性是否产生影响,这些问题尚不清楚。因此本研究以复合性状转基因水稻T2A-1、受体水稻明恢63(MH63)以及3种杂草稻的杂交后代为研究对象,研究其种子萌发特性以及在两种氮素浓度条件下的苗期生长特性,为评价T2A-1与杂草稻发生基因漂移后可能产生的生态风险提供参考。
1 材料与方法
1.1 试验材料
复合性状转基因水稻T2A-1及其非转基因受体水稻明恢63(MH63)由华中农业大学提供,3种来源于广东茂名(WRMM)、江苏泰州(WRTZ)和湖南益阳(WRYY)的杂草稻由南京农业大学杂草研究室试验人员采集。T2A-1及MH63与3种杂草稻杂交的12种F1于2018年人工杂交获得,F2、F3和F4由上一代植株自交获得,供试材料均保存在−5℃冷库中备用。
以T2A-1为亲本的携带抗性基因的杂交后代称为抗性杂交后代,表示为F1+、F2+、F3+和F4+;以MH63为亲本的杂交后代称为非抗性杂交后代,表示为F1−、F2−、F3−和F4−。试验材料见表1和表2。
1.2 试验设计
1.2.1 种子发芽力
试验于南京农业大学牌楼温室进行。每个材料选取籽粒饱满的种子每种100粒,放置于50℃烘箱中处理 48 h,打破种子休眠,将破休眠后的种子用75%酒精消毒浸种,之后把种子均匀摆放在垫有2层滤纸(用10 mL蒸馏水浸湿)的培养皿中,置于25℃培养箱中培养(12 h光照)。每隔24 h统计一次种子发芽数,14 d后计算发芽势与发芽指数。重复4次。
发芽势=发芽峰期中发芽种子数/总种子数。发芽峰期指在规定期限的最初1/3期间内种子发芽数之和。
发芽指数=∑t/t=当天的发芽数/发芽日数。待所有可发芽种子均发芽后,统计发芽率(发芽率=发芽种子数/总发芽种子数)。
在种子发芽高峰期(约培养3~4 d),从发芽的种子中选取萌发时间一致且芽长相似(小于1 cm)的种子,以T2A-1为父本的杂交后代的种子置于盛有40 mg/L的草胺膦溶液(18%的草胺膦可溶液剂,拜耳股份公司)的托盘中培养,并以T2A-1为对照,以T2A-1为母本的杂交后代和以MH63为亲本的杂交后代种子置于盛有蒸馏水的托盘中培养,托盘均垫有2层滤纸,种子胚根浸入药液或蒸馏水。培养温度为28℃,相对湿度为70%~80%,光周期为光照12 h/黑暗12 h。
培养一周后,观察以T2A-1为父本的杂交后代种子胚根和胚芽生长是否正常,能正常生长的植株采用基因特异性引物扩增的方法进行分子检测,确认供试材料为携带抗性基因的杂交后代。对于以T2A-1为母本的杂交后代和以MH63为亲本的杂交后代,因真杂交种扩增片段应包括杂草稻红色果皮基因()118 bp和栽培水稻白色果皮基因()104 bp片段,采用和特异性引物扩增的方法对幼苗进行分子检测[13]。
将真杂交种长势一致的幼苗转移至氮素营养液中培养15 d,培养温度为白天35℃±5℃,夜间20℃±5℃,自然光照。水稻培养液设置低氮(20 mg/L)与正常氮(40 mg/L)2种处理,每种试验材料重复4次。水稻培养液采用国际水稻所营养液配方。每种杂交后代均设置父本和母本作为对照处理。
1.2.2 根系发育指标测定
幼苗生长至15 d,使用根系扫描仪(LC-4800,LEGENTSYS-SINTEK,加拿大),测量所有供试材料总根长、种子根长、不定根数、最长三条不定根的平均长度、根系表面积、根平均直径、根系体积、根总分支数。
1.2.3 株高、干物质量测定
幼苗生长至15 d,测量所有供试材料株高,并放入烘箱中在60℃烘干24 h,测量所有供试材料整株干物质量。
1.3 数据统计
以处理为单位,运用SPSS 20.0中的Duncan新复极差测验,比较抗性杂交后代(F+和RF+)与非抗性杂交后代(F−和RF−)或相应杂草稻的发芽率、发芽势和发芽指数、株高、整株干物质量和根系发育各指标的差异显著性。

表1 水稻和杂草稻的来源和主要特征

表2 水稻和杂草稻的杂交后代
=1~4,表示F1、F2、F3和F4。
=1-4,means F1, F2, F3and F4.
以F−、RF−和3种杂草稻的根系发育指标为1,F+与RF+与它们的各项指标比值的平均值定义为相对根系发育指标。运用SPSS 20.0中的独立样本检验比较分析F+和RF+与相应的F−和RF−或杂草稻相对根系发育指标的差异。
2 结果与分析
2.1 杂交后代与杂草稻种子发芽力比较
所有供试材料的发芽率均高于95%。在F1~F4发芽试验中,所有抗性杂交后代与其非抗性杂交后代相比,发芽势与发芽指数均无显著差异(表3~表6)。

表3 杂草稻及与转基因水稻T2A-1杂交F1的发芽力
采用Duncan新复极差测验法分析F1和其相应杂草稻亲本的种子发芽力,不同小写字母表示同种杂草稻的杂交后代以及杂草稻之间的差异显著性(<0.05,Mean±SE,=4)。
The germination ability of F1hybrids and their corresponding weedy rice were analyzed by Duncan's Multiple Range Test. The different lowercase letter means significant difference among F1hybrids of the same weedy rice and their corresponding weedy rice (< 0.05, Mean±SE,=4).

表4 杂草稻及与转基因水稻T2A-1的杂交F2的发芽力
采用Duncan新复极差测验法分析F2及其相应杂草稻亲本的种子发芽力。不同小写字母表示同种杂草稻的杂交后代以及杂草稻间的差异显著性。<0.05,Mean±SE,=4。
The germination ability of F2hybrids and their corresponding weedy rice were analyzed by Duncan's Multiple Range Test. Different lowercase letters mean significant difference among F2hybrids from weedy rice and their corresponding weedy rice.<0.05, Mean±SE,=4.
以上结果说明对供试3种杂草稻的杂交后代,抗性基因的导入没有改变杂交后代的发芽力。WRMM和WRYY杂交后代的发芽势和发芽指数均显著低于相应杂草稻,这说明它们的杂交后代种子发芽整齐度和发芽速率均低于其杂草稻;但WRTZ的杂交后代发芽势与WRTZ相当,发芽指数均显著低于WRTZ,这说明WRTZ杂交后代的发芽整齐度与其杂草稻相当,但发芽速率均低于其杂草稻。同种杂草稻杂交后代的发芽力并没有随代数的增加而发生明显改变。

表5 杂草稻及其与转基因水稻T2A-1的杂交F3的发芽力
采用Duncan新复极差测验法分析F3和其相应杂草稻亲本的种子发芽力。不同小写字母表示同种杂草稻的杂交后代以及杂草稻间的差异显著性。<0.05, Mean±SE,=4。
The germination ability of F3hybrids and their corresponding weedy rice were analyzed by Duncan's Multiple Range Test. Different lowercase letters mean significant difference among F3hybrids derived from the same weedy rice and their corresponding weedy rice.< 0.05, Mean±SE,=4.
表6 杂草稻及与转基因水稻T2A-1的杂交F4的发芽力
采用Duncan新复极差测验法分析F4和其相应杂草稻亲本的种子发芽力,不同小写字母表示同种杂草稻的杂交后代以及杂草稻之间的差异显著性。< 0.05, Mean±SE,=4。
The germination ability of F4hybrids and their corresponding weedy rice were analyzed by Duncan's Multiple Range Test. Different lowercase letter mean significant difference among F4hybrids of the same weedy rice and their corresponding weedy rice.< 0.05, Mean±SE,=4.
2.2 杂交后代与杂草稻株高及整株干物质量
正常氮和低氮环境下,所有抗性杂交后代与其非抗性杂交后代相比,株高与整株干物质量均无显著差异。与相应的杂草稻相比,F1~F4规律一致。正常氮条件下,FMM+和RFMM+的株高显著低于其杂草稻WRMM,而整株干物质量与杂草稻WRMM相当;FYY+和RFYY+的株高及干物质量均与WRYY相当;FTZ+和RFnTZ+的株高显著高于其杂草稻,但整株干物质量显著低于其杂草稻WRTZ(表7)。低氮条件下与相应杂草稻相比,FMM+和RFMM+的株高显著低于WRMM,而整株干物质量与WRMM相当;FYY+和RFYY+的株高显著低于WRYY,但整株干物质量与WRYY相当。FTZ+和RFTZ+的株高显著高于WRTZ,但整株干物质量显著低于其杂草稻WRTZ。
除WRYY的杂交后代正常氮条件下株高与WRYY无显著差异,而在低氮条件下显著低于WRYY外,WRTZ杂交后代在正常氮条件与低氮条件下与相应杂草稻相比的趋势一致,均表现杂交后代苗期整株干物质量低于其杂草稻。相比而言,低氮条件下两者的差异更为明显。
2.3 杂交后代与杂草稻根系发育指标
正常氮条件下,所有抗性后代F+与非抗性后代F−相比,根系发育各指标均无显著差异(图1)。与相应杂草稻相比,F1~F4规律基本一致。FTZ+和RFTZ+的相对根系发育指标显著低于WRTZ(图2),这主要体现在除根平均直径外,FTZ+和RFTZ+的其他根系发育指标包括种子根长、总根长、不定根数、根系表面积及根体积等均显著低于WRTZ。FMM+和RFMM+以及FYY+和RFYY+的相对根系发育指标与其相应杂草稻均无显著差异,但部分指标存在差异,其中FMM+和RFMM+的种子根长均显著低于WRMM,FYY+和RFYY+的总根长低于WRYY。
低氮条件下,抗性后代F+与非抗性后代F−相比,规律并不完全一致。F1MM+、RF1MM+、F1YY+、RF2TZ+、RF2YY+、F3MM+、RF3MM+、F4MM+、RF4MM+以及RF4TZ+,这些抗性后代的相对根系发育指标均显著低于相应的非抗性后代(图1)。其他3种F1+、4种F2+和4种F3+以及3种F4+与相应的非抗性后代相对根系发育指标并无显著差异。与相应的杂草稻相比,3种杂草稻的抗性杂交后代F+相对根系发育指标均显著低于相应的杂草稻(图2)。这主要体现在3种杂草稻的杂交后代的8个根系发育指标总体均显著低于其相应的杂草稻。这说明即使在氮元素不足条件下,3种杂草稻根系发育受抑制情况轻于其抗性杂交后代。

表7 杂草稻及杂交后代的株高及干物质量
采用Duncan新复极差测验法分析杂交Fn和相应杂草稻亲本的株高及干生物量, 不同小写字母表示同种杂草稻的杂交后代以及杂草稻之间的差异显著性。<0.05, Mean±SE,=4。
The plant height and whole plant dry biomass of Fnhybrid and its corresponding weedy rice were analyzed by Duncan's Multiple Range Test. Different lowercase letter mean significant difference among Fnhybrids of the same weedy rice and their corresponding weedy rice.< 0.05, Mean±SE,=4.
3 讨论
3.1 种子发芽力
一年生植物开始于种子发芽,种子活力决定了其萌发率、出苗率、出苗整齐度,以及苗的健壮程度,也是其适应性的重要决定因子。衡量种子活力的指标有发芽率,种子发芽率较高,会给种群带来数量上的优势,除发芽率外,表示种子发芽整齐度和发芽速率的发芽势及发芽指数也是衡量种子活力的重要指标[30-31]。杂草稻具有较高的发芽势是其在各种环境条件下迅速出苗,占据生存空间的主要原因。李振博等[32]研究温度、pH 值、水势及NaCl 胁迫对杂草稻与常规栽培稻发芽势影响,发现在15℃低温、pH值为6.0和8.0、−0.3 MPa水势以及180 nmol/L NaCl等逆境条件下杂草稻的发芽势均强于常规栽培稻。王楠等[33]研究发现杂草稻耐深播能力强,出苗能力好,甚至在地下16 cm也能萌发出土,与栽培稻相比具有明显的优势。刘睿等[26]发现江苏省6种杂草稻均比栽培稻早2 d发芽。本研究表明,T2A-1或受体水稻与杂草稻的杂交F1~F4尽管均具有较高的发芽率,但与相应杂草稻相比,无论是抗性杂交后代还是非抗性杂交后代的发芽势和发芽指数几乎均低于相应的杂草稻,这意味着杂交后代比杂草稻的出苗能力降低,因而杂交后代在苗期占据生存空间上不具有优势。对于以T2A-1为父本的后代在存活植株中,均检测到了429 bp基因和602 bp基因片段,说明这些后代均含有和基因。而以T2A-1为母本和以MH63为亲本的后代,均检测到118 bp和104 bp的目的片段,说明这些后代为真杂交种。

1~7分别代表T2A-1/MH63, FnMM+/FnMM−, RFnMM+/RFnMM−, FnYY+/FnYY−, RFnYY+/RFnYY−, FnTZ+/FnTZ−, RFnTZ+/RFnTZ−。在A图中, n=1; 在B图中, n=2; 如此类推。采用独立样本t检验分析相对根系发育(以F−、RF−和MH63的根系发育指标为1,F+与RF+与它们的各项指标比值的平均值定义为相对根系发育指标)。*表示Fn+与Fn−或RFn+与RFn−差异显著(P<0.05,Mean±SE,n=4)。
对于发芽率与发芽势,T2A-1的抗性杂交后代与其受体水稻的杂交后代均无显著差异。这说明外源基因的导入并未改变杂交后代的发芽率及发芽势。因此杂交后代发芽能力的降低是因为其具有栽培水稻的基因,这说明转基因水稻的基因漂移到杂草稻后产生的杂交后代在没有目标除草剂以及靶标虫压的选择下,杂交后代种子在出苗能力上弱于杂草稻。含有转基因的杂草稻后代相比于不含转基因的杂草稻无显著优势,并且杂草稻中含有的栽培稻基因或外源抗除草剂基因会被逐渐淘汰[34]。同时有研究发现抗虫转基因水稻在虫害发生较低的环境下,其生长、分蘖和产生种子的能力等适合度指标相比非转基因水稻亲本或不含转基因的杂种分离后代均显著降低[35-37],这与本研究结果相类似。
3.2 根系发育
植物根系是水分和养分吸收的主要器官,其形态和根系活力对土壤养分吸收有重要作用[27]。根系数量、生物量以及表面积与植物养分吸收和产量正相关[38-41]。水稻根系形态的总根长、总根表面积、总根体积、分支数与生物量、氮累积量呈正相关[42]。程建峰等[43]研究表明水稻根系总吸收面积和活跃吸收面积与氮素利用效率正相关。因此,如果植物根系较早发育,生长迅速,根系发达,它们就可快速利用水资源和营养物质,在竞争地下资源方面占据优势,因而这类植物具有较强竞争能力[44]。前人研究表明,杂草稻比栽培稻具有更强的根系,使得其具有更强的竞争能力。Schaedler[29]发现杂草稻与栽培稻对光的竞争一定程度上降低了两者的根系发育情况,然而杂草稻的根系发育仍然强于栽培稻。本研究中杂草稻和栽培稻种子发芽后在2种氮素水平下水培15 d后,三种杂草稻的杂交后代在正常氮条件下根系发育均低于或相当于亲本杂草稻,而在低氮环境下,杂交后代的根系发育均显著弱于杂草稻,特别是杂交后代的总根长、根体积、根系表面积具有劣势,说明杂交后代的根系量少,在根系发育和形态上均不具有强竞争力的基础,会影响杂交后代根系对养分的充分吸收。
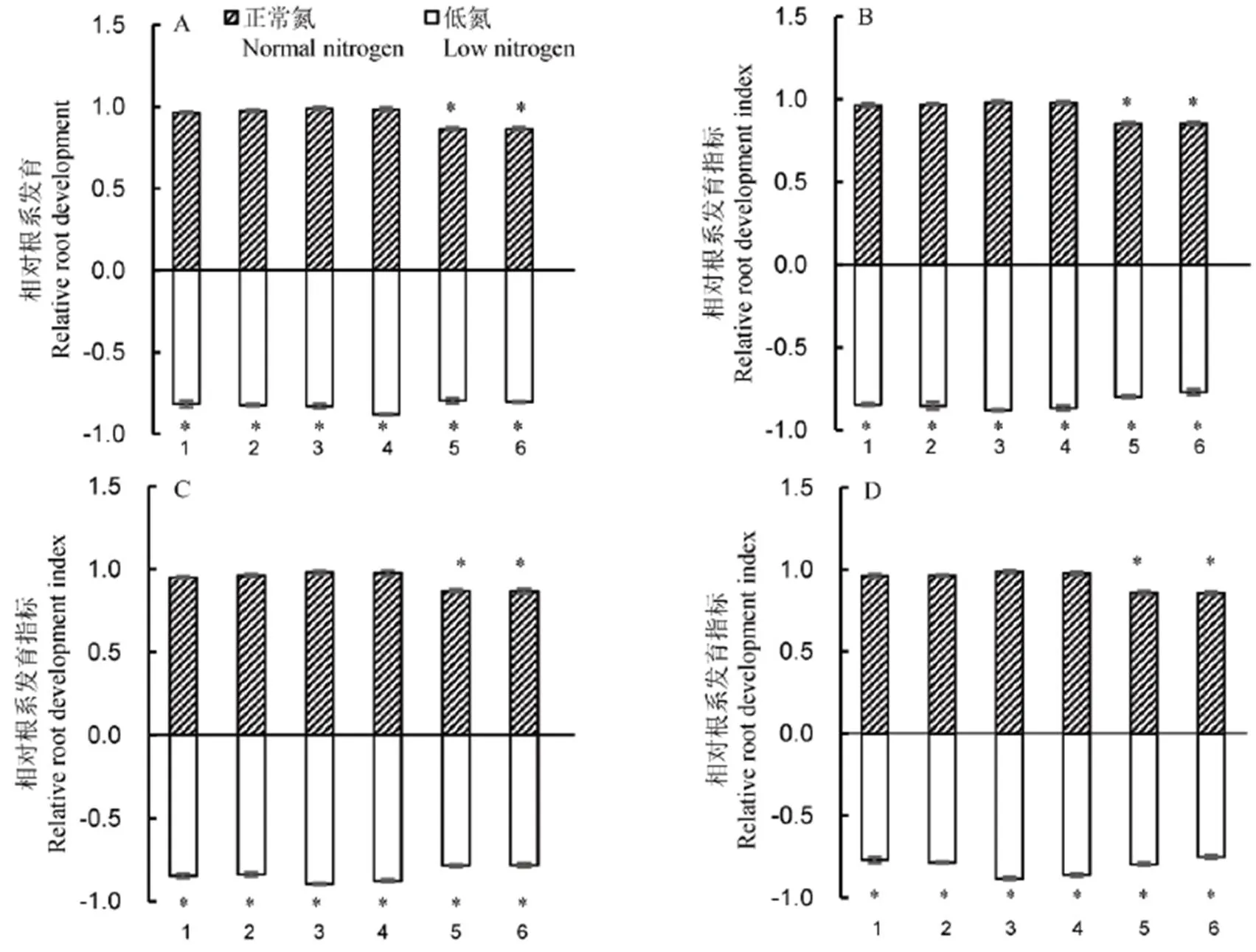
1~6分别代表FnMM+/WRMM, RFnMM+/WRMM, FnYY+/WRYY, RFnYY+/WRYY, FnTZ+/WRTZ, RFnTZ+/WRTZ。在A图中, n=1; 在B图中, n=2; 如此类推。采用独立样本t检验分析相对根系发育(以3种杂草稻的根系发育指标为1,F+与RF+与它们的各项指标比值的平均值定义为相对根系发育指标)。*表示Fn+或RFn+与杂草稻间差异显著(P<0.05, Mean±SE, n=4)。
在低氮条件下,T2A-1与抗性杂交后代的株高、干物质量以及根系发育相当于或低于MH63及其杂交后代,这可能是在养分不足的情况下维持外源基因的表达造成的非期望效应,具体生理机制仍需进一步研究。非期望效应是指将特定的外源基因整合进受体植物基因组时,除产生一些能稳定高效表达的符合原先设计需求的目标性状外,外源基因随机插入受体作物,或可能影响受体植物原有的生理代谢过程,或改变受体植物的原有性状等,这均是转基因设计过程中无法控制并预期的[45]。额外表达外源基因需要消耗物质与能量是引起非期望效应的重要原因之一[46-48]。
氮素作为植物生长发育所必需的第一大营养元素,在植物体内参与构成磷脂、蛋白质、核酸、植物激素、辅酶辅基及叶绿素等主要成分,对器官构建、物质代谢及植物的生长发育有不可替代的作用[49]。杂草稻能够高效利用氮肥, 这是其具有强竞争力的原因之一。Burgos等[18]以在阿肯色州广泛种植的高产水稻和当地最为普遍的杂草稻为试验对象,对比研究它们的氮肥利用效率,结果显示无氮肥施用时, 杂草稻与栽培稻的生物量相近,而施用氮肥后杂草稻茎秆的生物量迅速增加,显著高于栽培稻。由此推测杂草稻具有更高的氮肥利用效率。吴云艳等[28]的研究表明,杂草稻对氮肥的利用效率高于栽培稻,有些杂草稻对氮的利用甚至比栽培稻高出3倍。本研究虽然只设置了2种氮素条件,但基本可以反映杂交后代对土壤养分的吸收弱于杂草稻,从而影响其竞争能力。
在氮同化过程中,硝酸还原酶(NR)是植物体氮素同化和代谢过程的首个关键限速酶,在植物高效利用氮素过程中具有重要的生物学地位,因此NR活性可以反映植株的氮素营养状况和氮代谢水平。硝酸盐还原为氨后,植物须迅速进行氨的同化,植物体内95%以上的NH4+通过谷氨酰胺合成酶/谷氨酸合酶循环同化,两者均为参与氨同化的关键酶[50]。因此有待于进一步研究杂交后代在氮素同化水平上是否与杂草稻相当或处于劣势,从而从生理生化水平上探究杂交后代的氮同化水平。
贾世荣等[34]研究发现转基因水稻与野生稻的杂交后代相比野生稻在生长特性上发生的改变,可能是由于杂交后代受到栽培稻基因的影响所致。在本研究中T2A-1的发芽力和根系的总根长、种子根长以及根系表面积、不定根数在总体上均低于3种杂草稻,推测杂交后代因具有来自栽培水稻的基因,降低了杂交后代种子的发芽力和根系发育能力。从种子发芽力及根系发育来看,栽培水稻的基因消弱了杂交后代的生存竞争能力。但有研究表明栽培向日葵的部分性状,如快速生长能力,增强了栽培种和野生种杂交后代在野外的生存能力[51]。因此,只从根系发育情况不能全面评价栽培水稻的基因对杂交后代可能产生的影响,还需进一步研究栽培水稻其他性状可能带来的潜在影响。
[1] 翁绿水, 蒋利平, 肖国樱. 抗虫抗除草剂转基因水稻恢复系B2A68的培育[J]. 杂交水稻, 2013, 28(1): 63-67.
Weng L S, Jiang L P, Xiao G Y. Development of an insect-resistant and herbicide-resistant transgenic restorer line B2A68 in rice[J]., 2013, 28(1): 63-67. (in Chinese with English abstract)
[2] 陈丽, 邓力华, 陈芬, 肖国樱. 转和基因水稻的获得与性状鉴定[J]. 农业现代化研究, 2012, 33(1): 104-107.
Chen L, Deng L H, Chen F, Xiao G Y. Development and character identification of rice transformed byandgene[J]., 2012, 33(1): 104-107. (in Chinese with English abstract)
[3] 邓力华, 邓晓湘, 魏岁军, 曹正春, 唐俐, 肖国樱. 抗虫抗除草剂转基因水稻B1C893的获得与鉴定[J]. 杂交水稻, 2014, 29(1): 67-71.
Deng L H, Deng X X, Wei S J, Cao Z C, Tang L, Xiao G Y. Development and identification of herbicide and insect resistant transgenic plant B1C893 in rice[J]., 2014, 29(1): 67-71. (in Chinese with English abstract)
[4] Chen H, Tang W, Xu C, Li X, Lin Y, Zhang Q. Transgenic indica rice plants harboring a syntheticgene ofexhibit enhanced resistance against lepidopteran rice pests[J]., 2005, 111(7): 1330-1337.
[5] Yang X, Wang F, Su J, Lu B R. Limited fitness advantages of crop-weed hybrid progeny containing insect-resistant transgenes () in transgenic rice field[J]., 2012, 7(7): e41220.
[6] Gu X Y, Kianian S F, Foley M E. Seed dormancy imposed by covering tissues interrelates to shattering and seed morphological characteristics in weedy rice[J]., 2005, 45(3): 948-955.
[7] Akasaka M, Konishi S, IzawaT, Ushiki J. Histological and genetic characteristics associated with the seed-shattering habit of weedy rice (L.) from Okayama, Japan[J]., 2011, 61: 168-173.
[8] Baek J S, Chung N J. Seed wintering and deterioration characteristics between weedy and cultivated rice[J]., 2012, 5(1): 1-10.
[9] Zhao C, Xu W R, Li H W, Dai W M,Zhang Z, QiangS, SongX L. The rapid cytological process of grain determines early maturity in weedy rice[J]., 2021, 12: 711321-711321.
[10] Zhao C, Xu W R, Meng L C, Qiang S,Dai W M, Zhang Z,Song X L. Rapid endosperm development promotes early maturity in weedy rice (f.)[J]., 2019, 68(2): 168-178.
[11] Zhao C, Xu W R, Song X L, Dai W M, Dai L, Zhang Z, Qiang S. Early flowering and rapid grain filling determine early maturity and escape from harvesting in weedy rice[J]., 2018, 74(2): 465-476.
[12] Zhang F L, Lei S R, Liu Y, Guo L A, Yin Q, Song J, Wang D, Chang L J, Liu W J, Zhou X Q. Differences in ecological fitness between Bt transgenic rice and conventional rice under insect-infestation pressures[J]., 2012, 18(1): 35-41.
[13] Huang Y, Wang Y Y, Qiang S, Song X L, Dai W M. Fitness of F1 hybrids between stacked transgenic rice T1c-19 withgenes and weedy rice[J]., 2019, 18: 2-14.
[14] 肖国樱, 陈芬, 孟秋成, 周浩, 李锦江, 于江辉, 邓力华, 翁绿水.我国转基因抗除草剂水稻的生态风险与控制[J]. 农业生物技术学报, 2015, 23(1): 1-11.
Xiao G Y, Chen F, Meng Q C, Zhou H, Li J J, Yu J H, Deng L H, Weng L S. Ecological risk and management of herbicide-resistant transgenic rice () in China[J]., 2015, 23(1): 1-11. (in Chinese with English abstract)
[15] Song X L, Liu L L, Wang Z, Qiang S. Potential gene flow from transgenic rice(L.) to different weedy rice (f.) accessions based on reproductive compatibility[J]., 2009, 65(8): 862-869.
[16] Zuo J, Zhang L J, Song X L, Dai W M, Qiang S. Innate factors causing differences in gene flow frequency from transgenic rice to different weedy rice biotypes[J]., 2011, 67(6): 677-690.
[17] 王敏. 转基因水稻与杂草稻基因漂移率的检测技术及基因漂移研究[D]. 南京: 南京农业大学, 2019.
Wang M. Study on verifying technology of gene flow frequency and gene flow in transgenic rice and weedy rice[D]. Nanjing: Nanjing Agricultural University, 2019. (in Chinese with English abstract)
[18] Burgos N R, Norman R J, Gealy D R, Blank H. Competitive N uptake between rice and weedy rice[J]., 2006, 99(2-3): 96-105.
[19] Dai L, Song X L, He B Y, Valverde B E, Qiang S. Enhanced photosynthesis endows seedling growth vigour contributing to the competitive dominance of weedy rice over cultivated rice[J]., 2017, 73(7): 1410-1420.
[20] Ross M A, Harper J L. Occupation of biological space during seeding establishment[J]., 1972, 50: 116-122.
[21] Li B, Hara T. On the relative yield of plants in two-species mixture[J]., 1999, 85: 170-176.
[22] Rajjou L, Duval M, Gallardo K, Catusse J, Bally J, Job C, Job D. Seed germination and vigor[J]., 2012, 63(1): 507-533.
[23] Gommers C M M, Monte E. Seedling establishment: A dimmer switch-regulated process between dark and light signaling[J]., 2018, 176(2): 1061-1074.
[24] Foolad M R, Subbiah P, Zhang L P. Common QTL affect the rate of tomato seed germination under different stress and nonstress conditions[J]., 2007, 2007: 97386.
[25] Wang Y, Zhang G, Du J, Liu B, Wang M. Influence of transgenic hybrid rice expressing a fused gene derived fromandon primary insect pests and rice yield[J]., 2010, 29(2): 128-133.
[26] 刘睿, 强胜, 宋小玲, 陈世国, 戴伟民. 杂草稻苗期强竞争性的生理机制[J]. 植物保护学报, 2015, 42(1): 138-144.
Liu R, Qiang S, Song X L, Chen S G, Dai W M. Physiological mechanisms of strong competition of weedy rice at seedling stage[J]., 2015, 42(1): 138-144. (in Chinese with English abstract)
[27] Fitter A H. Lnfluence of mycorrhlzal infection on competition for phosphorus and potassium by two species[J]., 1977, 79: 119-125.
[28] 吴云艳. 杂草稻与栽培稻的地下营养竞争[J]. 西南农业学报, 2016, 29(3): 558-561.
Wu Y Y. Research on nutrient competition between root of weedy rice and that of cultivated rice[J]., 2016, 29(3): 558-561. (in Chinese with English abstract)
[29] Schaedler C E, Taborda C U M, Goulart F A P. Rice root growth and development in competition with weedy rice[J]., 2020, 38: e020216460.
[30] He Y, Chen S S, Liu K X, Chen Y J, Cheng Y H, Zeng P, Zhu P W, Xie T, Chen S L, Zhang H S, Cheng J P. OsHIPL1, a hedgehog-interacting protein-like 1 protein, increases seed vigor in rice[J]., 2022: 1-17.
[31] Li Y X, Zhou J H, Li Z, Qiao J Z, Quan R D, Wang J, Huang R F, Qin H.improves seed germination and salt tolerance by repressing ABA signaling in rice[J]., 2022, 189(2): 1110-1127.
[32] 李振博, 董立尧, 李俊, 张宏军, 杨玉清. 不同环境因子对江苏省杂草稻与常规栽培稻发芽势影响的比较[J]. 杂草科学, 2009(4): 28-31. (in Chinese with English abstract)
Li Z B, Dong L X, Li J, Zhang H J, Yang Y Q. Study on the influence of different environmental factors on the germination potential of weedy rice(L.) and cultivated rice from Jiangsu Province[J]., 2009, 4: 28-31. (in Chinese with English abstract)
[33] 王楠, 马殿荣, 贾德涛, 王莹, 陈温福. 北方杂草稻出苗特性的研究[J]. 华中农业大学学报, 2007, 26(6): 755-758.
Wang N, Ma D R, Jia D T, Wang Y, Chen W F.Germination characteristics of northern weedy rice[J]., 2007, 26(6): 755-758. (in Chinese with English abstract)
[34] Jia S, Yuan Q, Pei X, Wang F, Hu N, Yao K, Wang Z. Rice transgene flow: Its patterns, model and risk management[J]., 2014, 12: 1259-1270.
[35] 卢宝荣. 转基因逃逸及其环境生物安全评价研究进展:抗虫水稻案例分析[J]. 生物安全学报, 2014, 23(4): 217-223.
Lu B R. Assessing transgene escape and its environmental biosafety impacts: A case study in insect- resistant transgenic rice[J]., 2014, 23(4): 217-223.(in Chinese with English abstract)
[36] Xia H, Lu B R, Xu K, Wang W, Yang X, Yang C, Luo J, Lai F X, Ye W L, Fu Q. Enhanced yield performance of Bt rice under target-insect attacks: Implications for field insect management[J]., 2011, 20: 655-664.
[37] Yang X, Wang F, Su J, Lu B R. Limited fitness advantages of crop-weed hybrid progeny containing insect-resistant transgenes () in transgenic rice field[J]., 2012, 7(7): e41220.
[38] Wang Y, Mi G, Chen F, Zhang J, Zhang F. Response of root morphology to nitrate supply and its contribution to nitrogen accumulation in maize[J]., 2004, 27(12): 2189-2202.
[39] 田中伟, 樊永惠, 殷美, 王方瑞, 蔡剑, 姜东, 戴廷波.长江中下游小麦品种根系改良特征及其与产量的关系[J]. 作物学报, 2015, 41(4): 613-622.
Tian Z W, Fan Y H, Yin M, Wang F R, Cai J, Jiang D, Dai T B. Genetic improvement of root growth and its relationship with grain yield of wheat cultivars in the middle-lower Yangtze River[J]., 2015, 41(4): 613-622. (in Chinese with English abstract)
[40] 韩胜芳, 李淑文, 吴立强, 文宏达, 肖凯. 不同小麦品种氮效率与氮吸收对氮素供应的响应及生理机制[J]. 应用生态学报, 2007(4): 807-812.
Han S F, Li S W, Wu L Q, Wen H D, Xiao K. Responses and corresponding physiological mechanisms of different wheat varieties in their nitrogen efficiency and nitrogen uptake to nitrogen supply[J]., 2007(4): 807-812. (in Chinese with English abstract)
[41] Mu X, Chen F, Wu Q, Chen Q, Wang J, Yuan L, Mi G. Genetic improvement of root growth increases maize yield via enhanced post-silking nitrogen uptake[J]., 2015, 63: 55-61
[42] 陈晨, 龚海青, 张敬智, 郜红建. 水稻根系形态与氮素吸收累积的相关性分析[J]. 植物营养与肥料学报, 2017, 23(2): 333-341.
Chen C, Gong H Q, Zhang J Z, Gao H J. Correlation between root morphology and nitrogen uptake of rice[J]., 2017, 23(2): 333-341. (in Chinese with English abstract)
[43] 程建峰, 戴廷波, 荆奇, 姜东, 潘晓云, 曹卫星. 不同水稻基因型的根系形态生理特性与高效氮素吸收[J]. 土壤学报, 2007(2): 266-272.
Cheng J F, Dai T B, Jing Q, Jiang D, Pan X Y, Cao W X. Root morphologlcal and physiological characterisics in relation to nitrogen absorption efficiency in different rice genotypes[J]., 2007(2): 266-272. (in Chinese with English abstract)
[44] Bais H P, Sang-Wook P, Weir T L, Callaway R M, Vivanco J M. How plants communicate using the underground information superhighway[J]., 2004, 9(1): 26-32.
[45] Kotchoni O S, Gachomo E W, Mwangi M. Commercial production of genetically modified crops: A prognosis towards global acceptance[J]., 2005, 7: 681-688.
[46] Filipecki M, Malepszy S. Unintended consequences of plant transformation: A molecular insight[J]., 2006, 47(4): 277-286.
[47] Xia H, Lu B R, Su J, Chen R, Rong J, Song Z P, Wang F. Normal expression of insect-resistant transgene in progenies of common wild rice crossed with genetically modified rice: Its implication in ecological biosafety assessment[J]., 2009, 119: 635-644.
[48] Yang X, Li L, Cai X X, Wang F, Su J, Lu B R. Efficacy of insect-resistancetransgenes in F5-F7generations of rice crop-weed hybrid progeny: implications for assessing ecological impact of transgene flow[J]., 2015, 60(18): 1563-1571.
[49] 梁晓慧, 史树德. 作物氮素快速营养诊断及其在甜菜上的应用前景[J]. 北方农业学报, 2019, 47(1): 49-56.
Liang X H, Shi S D. Rapid diagnosis of crop nitrogen nutrition and prospects for its application in sugar beet[J]., 2019, 47(1): 49-56. (in Chinese with English abstract)
[50] 莫良玉. 高等植物氨基酸态氮营养效应研究[D]. 杭州:浙江大学, 2001.
Mo L Y. Studies on the effects of amino acid nitrogen on different plants under sterile culture[D]. Hangzhou: Zhejiang University, 2001. (in Chinese with English abstract)
[51] Mercer K L, Shaw R G, Wyse D L. Increased germination of diverse crop-wild hybrid sunflower seeds[J]., 2006, 16(3): 845-854.
Seedling Growth Characteristics of Hybrids Between Transgenic Rice with/Genes and Weedy Rice
XIAO Lequan, LI Lei, DAI Weimin, QIANG Sheng, SONG Xiaoling*
(Nanjing Agricultural University, Nanjing 210095, China; Corresponding author, email: sxl@njau.edu.com)
【Objectives】The studyaims to evaluate the seedling growth characteristics of hybrids between transgenic rice T2A-1 withgene and weedy rice, and lay an experimental basis for evaluating the potential ecological risks of gene flow from T2A-1 to weedy rice. 【Methods】F1+-F4+(positive) hybrids (those with transgenic insertion using the transgenic line T2A-1 as the paternal parent) , F1−-F4−(negative) hybrids(those using the non-transgenic receptor Minghui 63 (MH63) as the paternal parent) with three weedy rice (WRMM, WRTZ and WRYY) accessions, were used as experimental materials. Under hydroponic culture, germinability was measured. Eight root development indexeswere measured and relative root development index under normal nitrogen (40 mg/L) and low nitrogen (20 mg/L), and the seedling growth characteristics of hybrids were evaluated. 【Results】Transgenic positive hybrids had similar 个germinability to transgenic negative hybrids, but both showed weeker germination potential and germination index compared with the corresponding weedy rice. There were no significant differences in plant height, whole plant dry biomass, and root development between T2A-1 and MH63, and between transgenic positive and transgenic negative hybrids under normal nitrogen conditions. Compared with the weedy rice, hybrids derived from WRMM displayed significantly lower plant height, but similar whole plant dry biomass and the index of root development. Hybrids of WRYY displayed similar seedling growth characteristics with WRYY. While hybrids derived from WRTZ had significantly greater plant height, but significantly lower whole plant dry biomass and index of root development. Under low nitrogen conditions, transgenic positive hybrids had similar or significantly lower plant height, whole plant dry biomass and index of root development than transgene negative hybrids. The relative root development indexes of all hybrids were significantly lower compared with their corresponding weedy rice. 【Conclution】Compared with their corresponding weedy rice, transgenic positive hybrids displayed equivalent or lower seedling growth momentum under normal nitrogen conditions, but significantly weaker growth under low nitrogen conditions.
dual-transgenic rice; T2A-1; weedy rice; hybrids; seedling growth characteristic; root development
10.16819/j.1001-7216.2023.220612
2022-06-12;
2022-10-20。
国家转基因生物新品种培育重大专项(2016ZX08011-001); 海南省重大科技计划资助项目(ZDKJ202002-2-3)。
