沙棘GPD1和DGAT1双基因载体的构建及表达验证1)
2023-07-10赵思阳阮成江丁健卢顺光温秀凤胡建忠
赵思阳 阮成江 丁健 卢顺光 温秀凤 胡建忠
(大连民族大学,大连,116600) (水利部沙棘开发管理中心)
沙棘(Hippophaerhamnoides)为胡颓子科(Elaeagnaceae)沙棘属多年生落叶灌木或小乔木,耐寒抗旱的木本粮油树种。沙棘种子油中富含不饱和脂肪酸(如油酸(15.8%~32.8%)、亚油酸(21.7%~45.9%)、亚麻酸(14.1%~23.4%)、黄酮、甾醇等活性物质),具有预防和治疗癌症、抗氧化、抗衰老等功能[1]。甘油三酯(TAG)是木本油料树种种子储存脂质的主要形式,沙棘种子含油率仅为7%~11%,远不能满足全球日益扩大的沙棘种子油的需求。甘油-3-磷酸(G3P)是TAG合成的前体物质[2-3],甘油-3-磷酸脱氢酶(GPD1)将二羟基丙酮还原为甘油-3-磷酸,是糖酵解途径中的重要部分,也是促进油脂合成前体G3P合成的限速酶,直接影响3-磷酸甘油和甘油三酯水平[4-5]。Vigeolas et al.[6]在油菜中过表达甘油-3-磷酸脱氢酶(GPD1)酵母基因,发现发育中的种子中甘油-3-磷酸的水平增加了3~4倍,种子脂质质量分数增加40%。Liu et al.[7]在烟草中过表达油菜BnGPDH基因,与野生型相比,转基因植株的烟草种子含油量显著增加了4%。Kennedy通路是酰基辅酶A依赖性反应,也是是大多数植物合成TAG的主要途径[8]。在种子发育过程中,二酰甘油酰基转移酶(DGAT)通过将脂肪酰链从酰基辅酶a转移到sn-1,2-甘油二酯的sn-3位置,催化肯尼迪途径TAG合成的最后一步,因此,DGAT被认为是种子贮藏过程中油脂积累的重要酶[9-10]。Banilas et al.[11]研究发现橄榄DGAT1基因有助于种子油脂的合成,而基因DGAT1与DGAT2共表达有助于中果皮油脂的积累;Zhao et al.[12]揭示大豆基因GmDGAT1A和GmDGAT1B的组成性表达导致种子油含量增加;在微绿球藻中过表达拟南芥DGAT导致总脂质和TAG含量显著增加,且C16(C16:0+C16:1)急剧升高,C18(C18:0+C18:1)略有下降[13]。随着分子生物学技术的发展,将多个基因构建于一个植物表达载体,是实现多基因组装与转化的策略之一[14-15]。Chhikara et al.[16]将拟南芥二酰甘油酰基转移酶1(DGAT1)和酵母甘油-3-磷酸脱氢酶(GPD1)基因在亚麻荠种子中共表达,与野生型相比,转基因植株种子含油率升高了8%~13%,油酸质量分数降低26%,亚油酸质量分数升高23%。
沙棘种子发育期油脂合成积累源于源基因GPD1和汇基因DGAT的协同表达[17]。因而,调控GPD1和DGAT基因在提高种子含油量方面有潜在效果。本研究旨在成功构建沙棘油脂合成关键基因GPD1和DGAT1的双价载体pCAMBIA1301-HrGPD1-HrDGAT1,通过拟南芥异源表达对转基因植株脂肪酸不同组份的比例和含油率进行测定,验证共表达HrGPD1和HrDGAT1对沙棘种子品质的影响,为培育高品质沙棘提供科学依据。
1 材料与方法
1.1 试验材料
选取大连民族大学校园内的沙棘‘实优1号’的种子为研究对象,选用野生型拟南芥(Col-0)作为目的基因的转化受体。
1.2 沙棘GPD1和DGAT1基因的克隆
Trizol法提取沙棘‘实优1号’种子的RNA,反转录合成cDNA模板,RT-PCR扩增GPD1、DGAT1基因的CDS区,扩增GPD1基因的引物添加KpnI酶切位点,扩增DGAT1基因的引物分别添加BglII和BstEII酶切位点(见表1)。聚合酶链式反应(PCR扩增)程序:95 ℃(30 s),39个循环:95 ℃(15 s)、51 ℃(15 s)、72 ℃(1 min),72 ℃(5 min)。PCR产物琼脂糖凝胶电泳(见图1),切胶回收纯化目的扩增产物条带后进行测序。

表1 目的基因引物序列
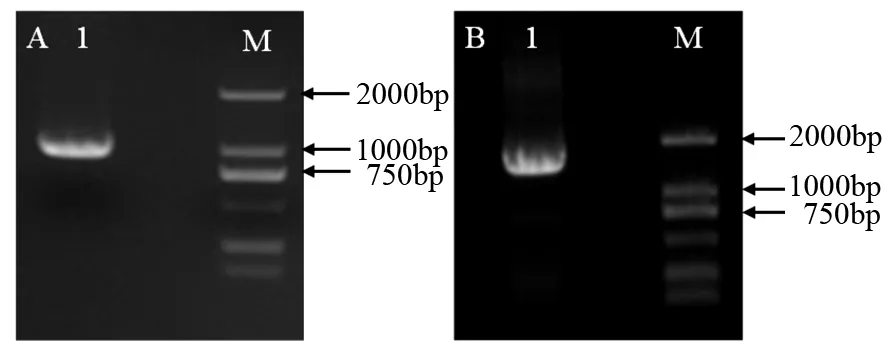
M为DL2000 DNA标识;A1为GPD1 PCR产物;B1为DGAT1 PCR产物。
1.3 双价表达载体的构建及转化
用限制性内切酶KpnI单酶切载体pCAMBIA1301-KY使其线性化,酶切产物纯化后用ClonExpress-II One Step Cloning Kit(ClonExpress-II一步克隆试剂盒,诺维赞生物)与HrGPD1基因的PCR纯化产物进行同源重组,取In-Fusion重组产物热转化至大肠杆菌DH5α细胞,涂布平板,37 ℃过夜培养。挑选PCR阳性的转化子摇菌培养提取质粒,同时扩增产物送测序,测序比对结果正确后提取质粒pCAMBIA1301-HrGPD1。将pCAMBIA1301-HrGPD1质粒用BglII和BstEII双酶切使其线性化,酶切产物纯化后与HrDGAT1基因的PCR产物进行重组反应,重组反应与HrGPD1基因连接相同的转化和验证步骤,测序比对正确后得到质粒pCAMBIA1301-HrGPD1-HrDGAT1。利用农杆菌介导法,将植物表达载体转化野生型拟南芥(Col-0),取适量野生拟南芥种子(T0)经潮霉素(质量浓度20 mg/L)筛选,选取长出真叶并能生根的抗性苗移栽,移栽苗正常开花后,取适量叶片提取DNA,以野生型拟南芥(Col-0)的cDNA作为对照(WT),检测引物为插入目的基因两侧的载体序列(见表1),上游统一用pCAMBIA1301-KY载体中的启动子片段35S-F。PCR扩增HrGPD1、HrDGAT1基因,电泳后按正确条带确定阳性植株。
1.4 目的基因荧光定量PCR分析方法
取适量转基因一代(T1)拟南芥植株的叶片提取RNA并反转录为cDNA,以野生型拟南芥的cDNA作为对照,以拟南芥actin(AT-actin,表1)为内参基因进行qPCR扩增检测,确定HrGPD1和HrDGAT1同时在转基因拟南芥中过表达。所用的荧光试剂为AceQ® qPCR SYBR® Green Master Mix(Vazyme # Q111-02/AA)。PCR反应体系为10 μL。每个植株样品3个技术重复。扩增的反应条件为:95 ℃ 5 min;95 ℃ 10 s,58 ℃ 30 s,40个循环;95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s;40 ℃ 5 min。目的基因相对表达量用2-ΔΔCT法计算。采用SPSS 20.0软件对数据进行统计分析,用邓肯检验分析组间差异性。
1.5 转基因T2拟南芥种子含油率测定
收取pCGD-11拟南芥株系的T2代纯合株系拟南芥的种子,采用氯仿甲醇法[18]测定种子含油率。以野生型拟南芥种子作为对照,每份种子为0.2 g,设置3次重复。种子干燥后研磨,称取粉末质量为m,加入2 mL甲醇和4 mL氯仿涡旋混匀2 min,超声30 min后离心,取上清加入1/4体积质量分数为0.88%氯化钾溶液,混匀静置30 min,收集下层溶液加入到已知质量(m0)的样品瓶中,挥发至恒质量后,称量油的质量(m1),含油率=(m1-m0)/m。
1.6 转基因二代(T2)拟南芥种子脂肪酸组分及比例测定
采用先皂化后甲酯化的方法对脂肪酸组成进行测定分析。甲酯化根据丁健等[19]的方法,适当改进。以野生型拟南芥种子作为对照,设置3个重复,取1.5节提取的转基因拟南芥种子油1 μL于4 mL贮样瓶,加200 μL正己烷,振荡溶解样品,加入500 μL浓度1 mol/L氢氧化钾甲醇溶液(现配),充分振荡;密封后,60 ℃水浴30 min,期间振荡2~3次,冷却;加入1 mL三氟化硼甲醇溶液,密封后,再于60 ℃水浴30 min,期间振荡2~3次,冷却;加入1 mL饱和氯化钠溶液和2 mL正己烷,剧烈振荡1 min,静置,分层后,收集上层溶液,加入新离心管;取适量无水硫酸钠加入离心管振荡干燥,静置;抽取上层清液过0.22 μm有机膜,用于气相色谱质谱联用进行脂肪酸组分分析。
GC-MS条件:采用DB-23色谱柱(60.00 m×0.25 mm,模厚0.25 μm),进样口温度230 ℃,载气流速1 mL/min,进样量1 μL,分流比40∶1。柱箱升温程序:温度50 ℃以15 ℃/min升温至200 ℃,保持15 min;以3 ℃/min升温至215 ℃,保持10 min;以3 ℃/min升温至230 ℃,保持5 min。离子源(EI)温度230 ℃,传输线温度245 ℃,检测电压1 700 V,溶剂延迟5 min,质量扫描范围(m/z)45~400 u。根据37种脂肪酸甲酯标准品建立的质谱库对样品定性分析,采用峰面积归一化法[20]对脂肪酸进行定量分析。
2 结果与分析
2.1 沙棘GPD1和DGAT1双基因载体的构建和转化
以沙棘种子发育20 d的种子cDNA为模板,PCR扩增GPD1和DGAT1基因,电泳结果显示扩增出975和1 608 bp的条带(见图1)。将构建成功的重组质粒分别转入农杆菌GV3101中,通过拟南芥蘸花法侵染野生型拟南芥,取适量T0代种子经潮霉素筛选(见图2A),选取长出真叶并能生根的抗性苗移栽(见图2B)。拟南芥移栽苗正常开花后,取T1代转基因植株的叶片提取DNA,以野生型拟南芥(WT)的cDNA作为对照,PCR分别扩增HrGPD1、HrDGAT1基因,共获得18株转基因阳性拟南芥植株(见图3),经测序结果比对,表明HrGPD1和HrDGAT1克隆到植物表达载体pCAMBIA1301-ky中,成功构建双价共表达载体pCMABIA1301-HrGPD1-HrDGAT1(pCGD)(见图4)。

图2 拟南芥转基因pCAMBIA-GPD1-DGAT1抗性筛选及抗性苗移栽

A为“A1~A18”为抗性植株GPD1基因PCR检测电泳;B为B1~B18”为抗性植株DGAT1基因PCR检测电泳;WT为对照野生型植株;“-”为纯水空白对照;“+”为质粒对照;“M”为DL2000标识。
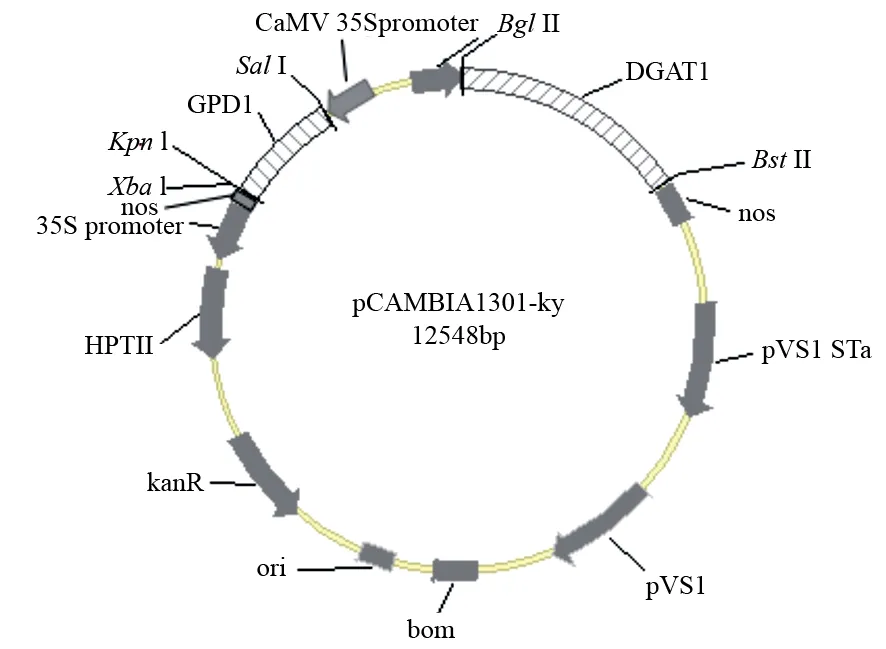
图4 pCMABIA1301-HrGPD1-HrDGAT1载体图谱
2.2 转基因拟南芥HrGPD1和HrDGAT1的表达水平
利用农杆菌介导法获得T1代转基因株系(pCGD-1、pCGD-10、pCGD-11)的叶片,以野生型(WT)为对照,通过qRT-PCR检测目的基因的表达。由表2可知,沙棘HrGPD1和HrDGAT在pCGD-1和pCGD-11转基因拟南芥植株中的显著高于对照表达,表明HrGPD1和HrDGAT共同转入拟南芥植株。在不同的转基因株系中,基因表达量差异比较大,这与基因的插入位点以及插入的拷贝数有关系。
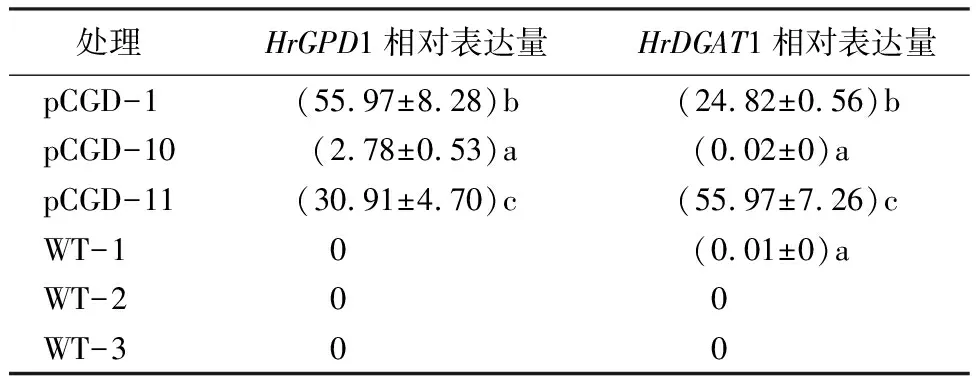
表2 转基因拟南芥株系中HrGPD1(A)和HrDGAT1(B)基因的相对表达量
2.3 过表达pCGD拟南芥的含油率和脂肪酸组分
由表3可知,野生型拟南芥种子油脂中油酸(C18:1)和亚油酸(C18:2)的比例分别为12.57%和33.78%,而转pCGD双价基因T2代拟南芥种子中油酸(C18:1)和亚油酸(C18:2)的比例分别为18.44%和36.94%,在拟南芥中共表达沙棘HrGPD1和HrDGAT1基因,导致油酸和亚油酸分别升高了46.70%和9.35%。野生型拟南芥种子含油率为222.75 mg/g,过表达pCGD的拟南芥种子含油率则为172.94 mg/g,转双价HrGPD1和HrDGAT1基因的拟南芥种子含油率降低了22.36%。

表3 过表达pCGD拟南芥种子的脂肪酸和含油率
3 讨论
重组DNA技术和植物转化系统的进步对将外源基因导入植物以产生具有改良性状的转基因植物非常有益。DGAT1通常是种子和果实含油量和脂肪酸组成的主要决定因素[21-22],控制其基因表达是增加含油量和改变脂肪组成的有效方法[23]。增加油酸等单不饱和脂肪酸的含量是一个优势,因为油酸具有氧化稳定性和低温流动性等化学性质[24]。大豆GmGPD1的过表达使大豆油酸显著增加了31.0%[25],麻风树中过表达AtDGAT1使种子油酸增加了9%~25%[26],向日葵HaDGAT1在酵母中的表达导致棕榈油酸和油酸分别增加了86.6%和81.6%[27]。在莱茵衣藻[28]、油莎草[29]、大豆[30]、亚麻荠[31]、破囊壶菌[32]中过表达DGAT2提高了油酸比例,说明分别过表达GPD1、DGAT可以提高种子不饱和脂肪酸的比例。本研究中,成功构建了沙棘pCAMBIA1301-GPD1-DGAT1双基因表达载体,在拟南芥中异源表达使拟南芥种子油酸和亚油酸分别提高了46.70%和9.35%,说明GPD1和DGAT1促进了不饱和脂肪酸的积累。4个参与甘油三酯合成和保护的重要基因(AtDGAT1、AtPDAT、AtWRI1和AtOLE)在水稻的组成性启动子下过表达,油酸和棕榈酸分别增加了28%和27%[33];Vanherck et al.[34]在烟草中过表达DGAT1、WRI1和Ole基因,油酸含量显著增加;在野生型拟南芥中共表达文冠果XsDGAT1和XsDGAT2,在发育中期,文冠果转基因植株种子的含油率达到最大值,且油酸和亚油酸的比例显著增加[35]。
用RNAi介导的基因沉默和过表达方法分别生成微绿球藻NoDGAT1a敲除和过表达转基因株系,发现TAG sn-1/sn-3位置的C16:0、C18:0的降低,以及TAG sn-2位置的C18:1的降低;NoDGAT1A的过表达导致三酰甘油(TAG)的比例增加了39%,伴随TAG sn1/sn-3位置的C16:0和C18:0增加,以及TAG sn-2位置的C18:1增加[36]。甘油的sn位置导致脂肪酸种类的差异,种子油存在不同种类的TAG。利用贮存在胞质中的酰基CoA池,在内质网上通过3种不同酰基转移酶的作用,合成三酰甘油[37]。饱和脂肪酸通常位于甘油分子的sn-1和sn-3位置,不饱和脂肪酸则大多发生在sn-2位置,二酰甘油酰基转移酶通常作用于sn-3位置[38]。说明DGAT1有助于将饱和及单不饱和脂肪酸转移到真核生物以合成TAG,且过表达DGAT基因可以引起TAG在sn-1/sn-3和sn-2位置的脂肪酸的比例增加。
DGAT1的高表达会影响底物、酶和产物间的关系,从而导致TAG代谢途径中的某些底物受到限制从而影响转基因植株种子含油率[39]。本研究中,共表达沙棘HrGPD1和HrDGAT1使转基因植株种子含油率降低了22.36%。
