miRNA-148a-3p对山羊卵巢颗粒细胞功能的影响
2023-07-08王钰锟李碧筠张义语丁文飞宋帅飞黄德利徐德军赵中权
王钰锟,李碧筠,张义语,丁文飞,王 磊,唐 雪,宋帅飞,姚 慧,黄德利,徐德军,赵中权*
(1.西南大学 动物科学技术学院,重庆 400715;2.重庆市大足区农业技术服务中心,重庆 402360)
卵巢的生长发育与颗粒细胞的增殖和凋亡有密切关系。颗粒细胞增殖和凋亡的紊乱会导致卵泡功能无法正常运作,从而降低哺乳动物繁殖性能[1]。自噬过程不仅维持细胞正常运作,同时也决定颗粒细胞的存活与死亡。颗粒细胞自噬过程可以调控细胞自身活力,参与卵泡生长发育,越来越多的证据表明颗粒细胞自噬与卵泡闭锁有关[2]。CHOI等[3]发现自噬通过调节Bcl-2/Bax比值调控黄体细胞凋亡,Bcl-2下调通过调节自噬小体积累诱导颗粒细胞凋亡。还有研究发现,卵泡闭锁期间自噬被激活,促进颗粒细胞凋亡[4]。雌二醇(E2)和孕激素(P4)等类固醇激素在牛、羊和猪卵泡生长发育中起不可忽视的作用。其中E2主要参与卵泡闭锁和颗粒细胞凋亡[5],P4会减缓卵泡发育、抑制颗粒细胞的有丝分裂、抑制细胞凋亡,能维持颗粒细胞状态、平衡卵泡数量以及提高卵巢储备[6]。
miR-148a-3p是miR-148/152家族的成员,该家族包含miR-148a、miR-148b和miR-152,这一家族的成员含有21~22个核苷酸[7]。miR-148/152家族成员在许多组织不同的生长发育阶段中差异表达,也在肿瘤的生长发育及发生过程中差异表达,其生理功能是多方面的,包括控制细胞增殖、分化及凋亡等[8]。相关报道表明miR-148/152家族成员可能会抑制癌基因表达,并且抑制癌细胞的增殖与迁移活动。同时,越来越多的研究发现,miR-148/152家族成员不仅在肿瘤疾病中发挥作用,在例如IgA肾病[9]、动脉粥样硬化病变[10]等的疾病中也起重要作用。
XU等[11]研究发现,miR-148a-3p通过靶向ROCK-1抑制膀胱癌细胞的增殖和迁移。BAO等[12]发现,miR-148a-3p可抑制癌症发展进程,是一种新型的胃癌诊断生物标志物,其在胃癌患者的组织和血浆样本中的表达量均明显下调,在胃癌细胞中过表达miR-148a-3p可以抑制细胞的增殖和转移。邓艳等[13]发现,miR-148a-3p在鹅卵巢颗粒细胞中可特异性靶向结合PPARγ,进而抑制孕酮合成的关键基因3β-HSD表达从而降低颗粒细胞类固醇激素的分泌。以上研究证明miR-148a-3p在调控细胞生命活力中发挥重要作用,但其在山羊卵巢颗粒细胞中的具体功能尚不清楚,因此本研究通过过表达或抑制miRNA-148a-3p,探究其对卵巢颗粒细胞增殖、凋亡、自噬及类固醇激素分泌的影响,为深入揭示miRNA调节卵泡发育与闭锁的机理提供理论基础。
1 材料与方法
1.1 主要试剂DMEM/F-12、FBS、胰酶购自美国Hyclone公司;miR-148a-3p mimics、miR-148a-3p inhibitor购自生工生物工程(上海)股份有限公司;EndoFectinTMMax转染试剂购自莱博斯生物科技有限公司;Annexin V-FITC/PI细胞凋亡检测试剂盒购自索莱宝科技有限公司;AdPlus-mcherry-GFP-LC3B、BeyoClickTMEdU-594细胞增殖检测试剂盒购自碧云天生物技术有限公司;CCK-8 试剂盒、BCA蛋白浓度测定试剂盒购自葆光生物技术有限公司;RNAiso、PrimeScriptTMRT reagent Kit with gDNA Eraser、TB GreenTMPremix Ex TaqTMⅢ、MiR-XTMmiRNA FirstStrand Synthesis购自TaKaRa公司;GAPDH Rabbit pAb、Bax Rabbit pAb、Bcl-2 Rabbit pAb、Anti-Rabbit IgG(H+L)购自Proteintech公司;PCNA Rabbit pAb、STAR Rabbit pAb、CYP11A1 Rabbit pAb、CYP19A1 Rabbit pAb购自Bioworld Technology公司;HSD3B1 Rabbit pAb、SQSTM1/p62 Rabbit pAb、LC3B Rabbit pAb购自ABclonal公司;山羊雌二醇ELISA检测试剂盒、山羊孕酮ELISA检测试剂盒购自博诺恒生物科技有限公司。
1.2 试验动物和细胞培养试验动物选用西南大学动物科学技术学院试验羊场的3~4月龄健康大足黑山羊,所有试验均得到西南大学动物实验伦理委员会批准。待山羊屠宰后,将卵巢取出用75%酒精喷洗,用无菌PBS缓冲液冲洗3遍,放入37℃无菌生理盐水中,带回实验室进行后续操作。山羊卵泡颗粒细胞的培养同文献[21]报道。
1.3 miRNA-148a-3p模拟物与抑制剂转染待细胞在6孔板密度达到70%~80%时按照转染试剂说明进行操作。5 μL miR-148a-3p mimics/NC、inhibitor/NC和5 μL EndoFectinTMMax转染试剂用DMEM/F-12稀释至150 μL,室温静置5 min。随后将上述稀释的溶液与EndoFectinTMMax溶液混合,轻柔吹打混匀后静置20 min,等待复合物充分形成。向6孔板中均匀滴加RNA-EndoFectinTM复合物,使其在细胞培养板中均匀扩散。在含有5% CO2的培养箱中于37℃孵育24~72 h后提取细胞总RNA或蛋白进行后续试验。
1.4 AdPlus-mCherry-GFP-LC3B转染在24孔板接种细胞后置于培养箱中培养24 h以上,待细胞密度在50%左右时对细胞进行过表达或者抑制miR-148a-3p处理,24 h后吸弃旧培养液,每孔加入300 μL 新鲜培养液,并分别加入40 μL病毒母液。感染24 h后吸弃培养板中含有病毒的培养液,每孔加入500 μL新鲜的完全培养液继续培养24 h,在适当条件诱导自噬1~2 h后在荧光显微镜下观察LC3B的荧光变化情况。
1.5 CCK8及流式细胞术转染miR-148a-3p模拟物或抑制剂处理72 h后每孔加入10 μL CCK8溶液,置于细胞培养箱中孵育2 h,用酶标测定仪检测450 nm处转染的吸光度值。用胰酶消化收集转染后的卵巢颗粒细胞,用预冷的PBS清洗细胞2次,1 700 r/min 离心5 min,弃上清,用Binding buffer缓冲液重悬细胞,调整细胞密度为107个/mL。在流式管中加入100 μL细胞悬液,随后分别加入5 μL Annexin V-PE和5 μL 7-AAD轻柔混匀,室温避光孵育15 min后用流式细胞仪检测细胞状态。
1.6 实时荧光定量PCR使用RNAiso试剂(TaKaRa,中国)从颗粒细胞中提取总RNA。使用分光光度计测定RNA质量浓度和纯度。根据PrimeScriptTMRT reagent Kit with gDNA Eraser说明书的要求首先去除基因组DNA,之后进行反转录反应。所有基因序列均源自NCBI(https://www.ncbi.nlm.nih.gov/),引物由生工生物工程(上海)股份有限公司合成,合成引物及序列见表1。根据TaKaRa公司的TB Green®Premix Ex TaqTMⅢ(Tli RNaseH Plus)试剂盒说明书,进行荧光定量PCR试验。反应体系为15 μL:TB GreenTMPremix Ex TaqTMⅡ 7.5 μL、Forward primer(10 μmol/L)0.6 μL、Reverse primer(10 μmol/L)0.6 μL、模板(cDNA)1.2 μL、RNase Free dH2O 5.1 μL。经过95℃反应30 s、95℃反应5 s、60℃反应30 s的步骤进行40次循环,在63.5℃时检测荧光信号,溶解曲线依据具体引物可在65℃~95℃范围内进行调整,每0.5℃时读取1次反应Ct 值。

表1 PCR引物序列
1.7 总蛋白提取和Western blot依据动物全蛋白提取试剂盒的说明书提取经不同处理的细胞的总蛋白,提取的总蛋白在-80℃冷冻保存。采取BCA法测定蛋白质量浓度,将蛋白样品与蛋白上样缓冲液混合后沸水浴8~10 min,经电泳转膜等操作,将PVDF膜放入装有封闭液的培养皿中封闭2 h,结束后过夜孵育对应的一抗,随后二抗孵育2 h,利用显影液在照胶仪下观察蛋白相对表达量。
1.8 ELISA检测处理结束后收集细胞培养液至新的离心管,3 000 r/min离心10 min后收集上清液。制备50,25,12.5,6.25,3.125,1.562 5,0 mg/L 的标准品,将样品用标准稀释液按1∶4稀释后在96孔中加入50 μL的样品或不同质量浓度标准品至预包被板孔中。每孔加入辣根过氧化物酶标记的检测抗体100 μL,用封板膜封闭反应孔,恒温箱室温孵育60 min。用300 μL稀释为1×的洗涤液对96孔板洗涤,每次15~30 s,重复5次,在最后1次洗板结束后将板倒扣于厚吸水纸上拍打使其干燥。各加入50 μL A液和B液,在37℃条件下避光孵育15 min后将终止液50 μL加入96孔板中,立刻用酶标仪在450 nm波长条件下测定光密度值。
1.9 统计分析本试验使用GraphPad Prism软件进行统计分析,用独立t检验对两组数据进行比较分析。试验重复3次以上,数据结果采用平均值±标准误表示,P<0.05表示差异显著,P>0.05表示差异不显著。
2 结果
2.1 miR-148a-3p转染效率验证将miR-148a-3p模拟物与抑制剂及NC转染到卵巢颗粒细胞中,分别在24,48,72 h观察细胞转染情况并用qRT-PCR验证其转染效率。由图1可知,在转染后72 h,观察到细胞转染效率最高。利用qRT-PCR进行mRNA水平检测,发现转染miR-148a-3p mimics后72 h 时其在卵巢颗粒细胞中的表达量最高(P<0.01),因此后续试验选用转染后72 h的细胞。由于miRNA inhibitor的干扰原理,未检测到miR-148a-3p表达量的变化。
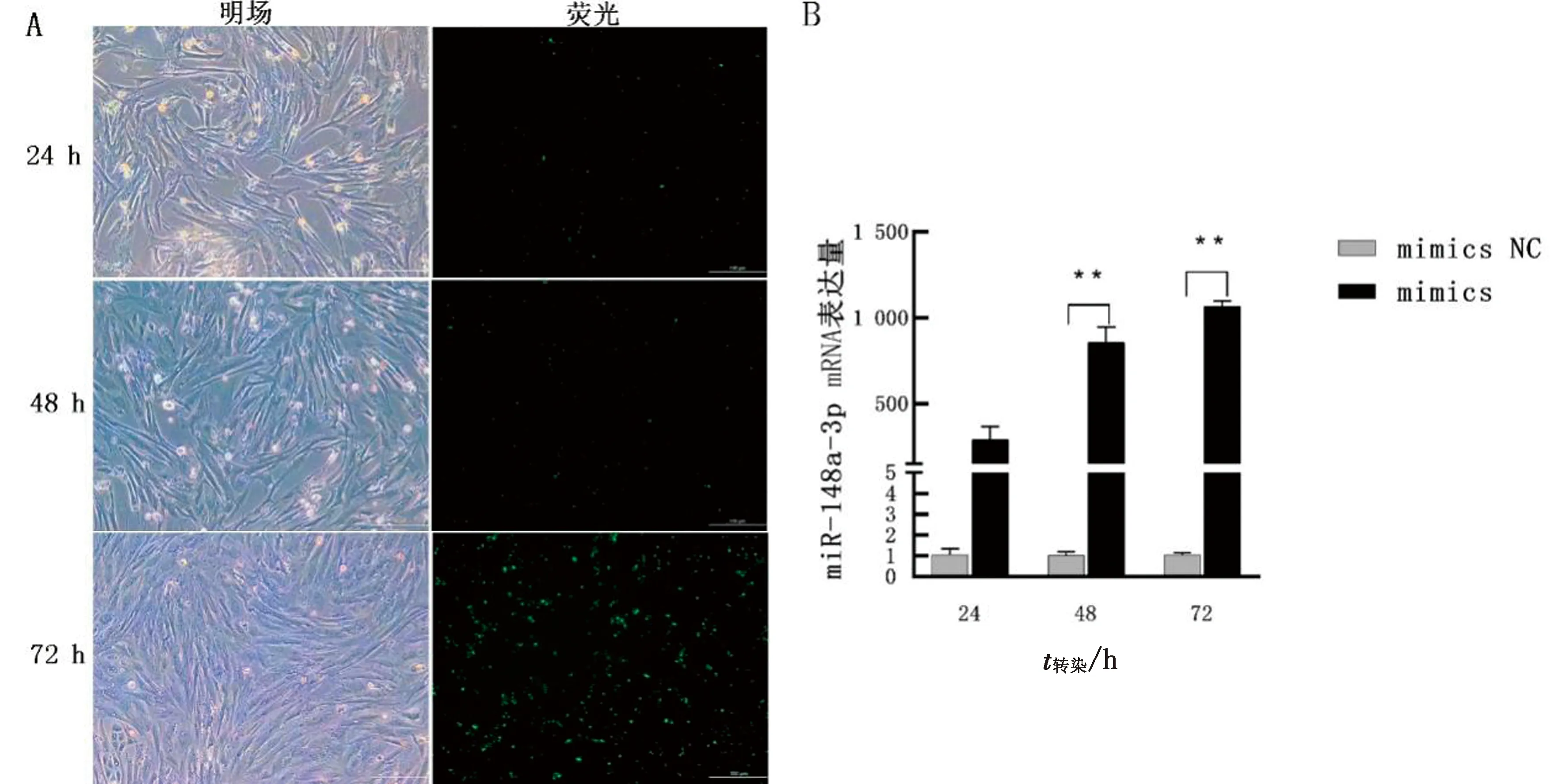
A.颗粒细胞转染miR-148a-3p不同时间后的荧光图(×100);B.转染后不同时间miR-148a-3p在颗粒细胞中的表达量
2.2 miR-148a-3p对卵巢颗粒细胞增殖的影响利用CCK8法测定细胞增殖活力,结果显示,转染后72 h,过表达miR-148a-3p极显著提高了细胞增殖活力(P<0.01),抑制miR-148a-3p降低了细胞增殖活力(P<0.05)(图2A)。过表达miR-148a-3p极显著上调EdU阳性细胞的比例(P<0.01)(图2B),但抑制miR-148a-3p对EdU阳性细胞的比例无显著影响(P>0.05)。通过qRT-PCR和Western blot检测基因的表达水平,结果显示,过表达miR-148a-3p极显著上调PCNA mRNA和蛋白的表达量(P<0.01),抑制miR-148a-3p显著下调PCNA mRNA和蛋白的表达量(P<0.05)(图2C、D)。流式细胞术结果显示,过表达miR-148a-3p使G1期细胞占比极显著降低(P<0.01),处于G2期的细胞占比极显著增加(P<0.01),但对处于S期细胞的占比没有影响。抑制miR-148a-3p表达使G2期细胞的占比显著降低(P<0.05),但对处于G1期和S期细胞的占比没有显著影响(图2E)。qRT-PCR结果显示,过表达miR-148a-3p后,显著升高了CCND1的mRNA(P<0.05),但其对CDK4和CCNE1没有显著影响(P>0.05);抑制miR-148a-3p对CDK4、CCND1和CCNE1的mRNA水平均无显著影响(P>0.05)(图2F)。

A.CCK8检测转染miR-148a-3p不同时间对颗粒细胞增殖活性的影响;B.EdU检测转染miR-148a-3p后颗粒细胞的增殖情况(×100);C,D.转染miR-148a-3p后颗粒细胞中PCNA的实时荧光定量及蛋白免疫印迹分析;E.miR-148a-3p对颗粒细胞周期的影响;F.转染miR-148a-3p后颗粒细胞中CDK4、CCND1和CCNE1的实时荧光定量分析
2.3 miR-148a-3p对卵巢颗粒细胞凋亡的影响使用流式细胞术检测过表达或抑制miR-148a-3p后颗粒细胞的凋亡情况。结果显示,过表达miR-148a-3p极显著下调细胞凋亡率(UR+LR)(P<0.01),由2.18%降低到1.19%,而抑制miR-148a-3p显著上调细胞凋亡率(UR+LR)(P<0.05),由1.35%升高到8.21%(图3A,B)。通过qRT-PCR和Western blot检测显示,过表达miR-148a-3p极显著上调Bcl-2/Bax的mRNA和蛋白的表达量(P<0.01)(图3C,D)。抑制miR-148a-3p对Bcl-2/Bax的mRNA和蛋白的表达无显著影响(P>0.05)(图3C,D)。上述结果表明,过表达miR-148a-3p可抑制颗粒细胞凋亡。
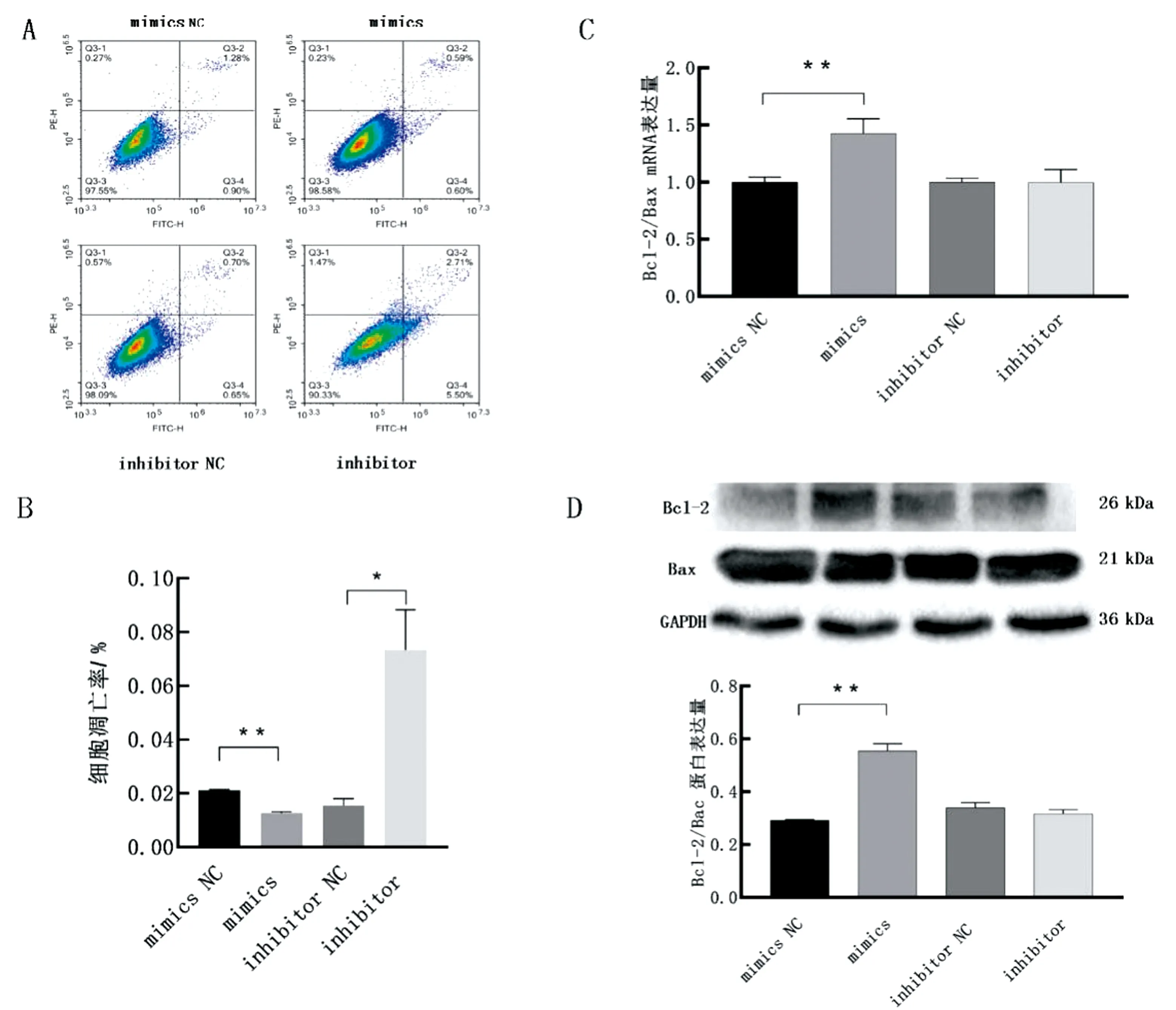
A,B.转染miR-148a-3p后颗粒细胞凋亡情况及定量分析;C,D.转染miR-148a-3p后颗粒细胞中Bcl-2/Bax的实时荧光定量及蛋白免疫印迹分析
2.4 miR-148a-3p对卵巢颗粒细胞自噬的影响在转染miR-148a-3p后24 h接种自噬LC3双标腺病毒,继续培养后48 h利用荧光倒置显微镜观测miR-148a-3p mimics NC组、miR-148a-3p mimics组、miR-148a-3p inhibitor NC组和miR-148a-3p inhibitor组细胞的自噬体数量。结果显示,过表达miR-148a-3p显著增加了自噬体的个数(P<0.05)(图4A,B),但抑制miR-148a-3p对自噬体的数量无明显影响。Western blot结果显示,过表达miR-148a-3p显著上调LC3-Ⅱ/LC3-Ⅰ比值(P<0.05),但对p62的蛋白表达量无明显影响,而抑制miR-148a-3p对LC3-Ⅱ/LC3-Ⅰ比值无明显影响(P>0.05),但显著上调p62的表达(P<0.05)(图 4C)。结果表明,在卵巢颗粒细胞中过表达miR-148a-3p可诱导细胞自噬。

A.用mCherry-GFP-LC3B腺病毒转染颗粒细胞检测转染miR-148a-3p后的自噬水平(×100);B.转染miR-148a-3p后mCherry-GFP-LC3B的定量分析;C.转染miR-148a-3p后颗粒细胞中LC3B及P62的蛋白免疫印迹分析
2.5 miR-148a-3p对颗粒细胞分泌类固醇激素的影响使用ELISA试剂盒检测在颗粒细胞中过表达或抑制miR-148a-3p 72 h时细胞培养液中E2和P4的含量。结果显示,过表达miR-148a-3p极显著促进了P4的分泌(P<0.01),而抑制miR-148a-3p则极显著下调了P4的分泌(P<0.01)。但是过表达和抑制miR-148a-3p都对E2的分泌无显著影响(P>0.05)(图5A)。qRT-PCR检测显示,在颗粒细胞中过表达miR-148a-3p后3β-HSD的mRNA相对表达量极显著增加(P<0.01),StAR和CYP11A1的mRNA相对表达量也显著增加(P<0.05),但其对CYP19A1的mRNA表达无显著影响。抑制miR-148a-3p对3β-HSD、StAR、CYP11A1和CYP19A1均无明显影响(图5B)。Western blot结果显示,过表达miR-148a-3p显著上调3β-HSD、StAR、CYP11A1的蛋白表达(P<0.05),但对CYP19A1的蛋白表达无显著影响。抑制miR-148a-3p对3β-HSD、StAR、CYP11A1及CYP19A1的蛋白表达量均无显著影响(图5C)。结果表明,上调miR-148a-3p可促进卵巢颗粒细胞孕酮分泌。

A.ELISA测定转染miR-148a-3p后颗粒细胞E2及P4的分泌量;B,C.转染miR-148a-3p后颗粒细胞中3β-HSD、StAR、CYP11A1和CYP19A1的实时荧光定量及蛋白免疫印迹分析
3 讨论
miRNA在卵巢颗粒细胞、卵母细胞和卵泡液中表达,影响家畜的繁殖能力。本试验探究了miR-148a-3p对卵巢颗粒细胞的增殖、凋亡、自噬及类固醇分泌的影响。先前研究发现,miR-148a-3p在调控细胞增殖中起重要作用,如SHANG等[14]发现miR-148a-3p可显著促进内皮细胞的增殖;LIU等[15]发现在肾小球中过表达miR-148a-3p可显著促进PCNA表达和细胞增殖。本研究结果与先前研究结果一致,发现过表达miR-148a-3p可促进PCNA在mRNA及蛋白水平的表达,促进卵巢颗粒细胞的增殖。进一步利用流式细胞术研究发现,miR-148a-3p能降低G1期细胞的比例,同时升高G2期细胞的比例。过表达miR-148a-3p可上调CCNE1的表达量,而CCNE1的主要功能是促使细胞进入S期,但本研究结果中S期细胞的比例未见增加,可能是细胞经过S期合成了DNA,进入了G2期。在颗粒细胞中过表达miR-148a-3p后G2期细胞比例增多,可能是完成S期后等待进入M期进行有丝分裂的细胞增多。以上结果表明miR-148a-3p可能通过上调CCNE1的表达促进细胞及时从G1期进入S期,从而促进卵巢颗粒细胞的增殖。
有研究显示,大多数miRNA在哺乳动物卵巢颗粒细胞凋亡中发挥着促进作用。如ZHANG等[16]发现,小鼠卵巢颗粒细胞中miRNA-122-5p通过调节BCL9促进细胞凋亡;但也有研究显示,miRNA对细胞凋亡有抑制作用,如ZHU等[17]发现,miR-222通过靶向THBS1基因来抑制猪卵巢颗粒细胞的凋亡。本研究通过流式细胞术、qRT-PCR及Western blot检测转染miR-148a-3p后卵巢颗粒细胞的凋亡率及Bcl-2和Bax的表达量。发现卵巢颗粒细胞中过表达miR-148a-3p下调细胞的凋亡率,同时上调Bcl-2/Bax数值;说明过表达miR-148a-3p可抑制卵巢颗粒细胞凋亡。
自噬是一种从酵母到哺乳动物都存在的保守机制,对生长发育等一系列生理过程有重要作用。尽管之前研究发现了颗粒细胞自噬对卵泡生长或闭锁的作用,但自噬在卵巢颗粒细胞中的具体作用及其机理仍不清楚。已有研究发现miR-148a-3p对自噬有调控作用,如LI等[18]发现在胃癌细胞中过表达miR-148a-3p会抑制细胞自噬。本研究利用qRT-PCR及Western blot检测了转染miR-148a-3p后卵巢颗粒细胞自噬情况及自噬相关基因LC3B、P62的表达量。结果发现在卵巢颗粒细胞中过表达miR-148a-3p上调自噬体的数量以及LC3-Ⅱ/LC3-Ⅰ的比值,说明过表达miR-148a-3p可诱导卵巢颗粒细胞自噬。
颗粒细胞是类固醇激素合成和分泌的主要来源,大量研究探究了miRNA影响颗粒细胞类固醇激素分泌的分子途径。如GAO等[19]发现miR-31靶向HSD17B14和FSHR,miR-20b靶向HSD17-B14,以影响猪卵巢颗粒细胞的凋亡和类固醇激素分泌。本研究利用ELISA、qRT-PCR及Western blot检测了E2、P4的分泌量和类固醇合成相关基因3β-HSD、StAR、CYP11A1和CYP19A1的表达量。结果发现在颗粒细胞中过表达miR-148a-3p使得P4的分泌量增加,并且上调了3β-HSD、StAR和CYP11A1的表达量,但不影响E2的分泌。
本研究中,抑制miR-148a-3p表达,没有下调EdU阳性细胞率,对Bcl-2/Bax的mRNA和蛋白表达也没有影响。相比较过表达miR-148a-3p能显著促进自噬水平,抑制miR-148a-3p并没有抑制自噬水平,抑制miR-148a-3p对类固醇合成相关基因3β-HSD、StAR、CYP11A1和CYP19A的表达也没有抑制作用。miR-148a-3p调控细胞增殖的靶基因和通路有很多,如miR-148a-3p通过调节DNMT1和UTF1的表达抑制宫颈癌细胞的增殖[20];miR-148a-3p通过靶向周期蛋白依赖性激酶6(CDK6)抑制急性髓细胞增殖[21]。当单独抑制某个基因或通路时,可能还会有其他的基因在起作用,不会造成显著的影响。因此,miR-148a-3p在卵巢颗粒细胞中具体通过哪些通路和途径调控细胞增殖还有待于进一步研究。
综上所述,本研究证明过表达miR-148a-3p可促进卵巢颗粒细胞增殖,抑制凋亡,诱导卵巢颗粒细胞自噬,促进孕酮的分泌,但不影响雌二醇的分泌。本研究为进一步了解miRNA调控卵巢颗粒细胞的生理功能提供了一定的理论基础。
