缩行匀株对小麦分蘖的影响及其生理机制
2023-06-27刘盼高珊李浩宇王翼尹宝重郭进考甄文超
刘盼, 高珊, 李浩宇, 王翼, 尹宝重, 郭进考, 甄文超,4,5*
(1.河北农业大学农学院,河北 保定 071001; 2.河北农业大学植物保护学院,河北 保定 071001; 3.石家庄市农林科学研究院,石家庄 050085; 4.河北农业大学,农业农村部华北节水重点实验室,河北 保定 071001; 5.河北农业大学,河北省作物生长调控重点实验室,河北 保定 071001)
小麦(Triticum aestivumL.)是我国第二大粮食作物,也是我国重要的商品粮和战略储备粮,在保障国家粮食安全中具有重要地位。分蘖是决定小麦叶面积和穗数的关键农艺性状,对小麦产量有重要贡献。同时,分蘖还可以减少包括干旱、低温等胁迫条件对植株的影响,提高植物对环境的适应性。因此,在生产中,可通过农艺措施来调控分蘖的数量和质量,进而为小麦的高产稳产奠定基础。分蘖的本质是一种侧枝,它们有独立的根和同化物形成器官,是保障小麦充足群体和穗数、提高产量的基础[1-2]。同时,分蘖的产生还会促进次生根的发生,对提高小麦水肥利用效率和抗逆能力具有重要意义。
分蘖是由腋生分生组织发育而来,是典型的数量性状,由遗传、激素和环境条件等共同调控[3-4]。研究显示,蘖芽生长通常受激素调节,其中吲哚乙酸(indoleacetic acid,IAA)和细胞分裂素(cytokinin,CTK)起重要作用。IAA在植物顶端合成并向下输送到分蘖基部,实现对蘖芽的抑制;CTK可加速IAA的运输,并促进其对蘖芽生长的抑制[5-6]。另外,小麦分蘖芽生长还受分蘖节中IAA和玉米素(zeatin,ZT)含量及IAA与ZT、脱落酸 (abscisic acid,ABA)与ZT比例的调节[7]。独脚金内酯(strigolactones,SLs)作为近年来发现的由类胡萝卜素衍生的植物激素,也能抑制腋芽生长[8-9]。赤霉素 (gibberellin,GA)可调节植株高度,其含量与分蘖数、株高呈负相关。研究表明,GA生物合成或信号传导缺陷的植物通常表现为矮秆、多蘖;多效唑是GA合成的抑制剂,外源施用多效唑能促进分蘖芽的萌发[1]。除此之外,糖也可作为某种信号,参与调控分蘖的发生[10]。
环境条件是影响植物分蘖的重要因素,如气温、土壤含水量、土壤养分含量、植株生存空间竞争等。其中,株行距配置是造成植株生存空间竞争效应的主要因素,对植物分蘖具有重要的调控作用[11]。研究表明,增加播种密度会加剧空间竞争,造成植株徒长,抑制分蘖[12-14];相同种植密度下,适度缩小行距会促进分蘖发生,但过小的行距则效果相反[15]。在小麦实际生产中,株距远小于行距,这也意味着小麦株间竞争要大于行间竞争[14]。因此,通过缩小行距、扩大株距,缩行匀株(row space reduction and plant space expansion,RRPE),减小株间竞争对促进小麦分蘖有积极意义。
海河平原是我国重要的小麦、玉米产区,其小麦产量约占全国总产15%[16-17]。由于该区为冬小麦-夏玉米一年两熟种植区,因此,前茬玉米的收获期及天气条件、土壤墒情等因素都会对后茬小麦播种造成影响,很多年份常因为各种因素造成晚播。播期晚、积温不足常导致分蘖偏少,因此,可通过缩行匀株的栽培模式来促进植株分蘖。虽然前人对缩行匀株促进分蘖有过一些研究,但研究降低株间竞争、促进植物分蘖的生理机制鲜见报道。基于此,本研究通过设置3个播期和2个行距处理,研究缩行匀株栽培模式对冬小麦分蘖的调控效应及其生理机制,旨在为该区冬小麦抗逆稳产栽培提供技术支持和理论依据。
1 材料与方法
1.1 试验设计
试验于2019—2021年小麦季(Y),在河北省辛集市马兰农场进行。试验田耕层土壤碱解氮1.27 g·kg-1,速效磷和速效钾含量分别为28.2和122.2 mg·kg-1。2个小麦生长季的降水量分别为139.8(2019—2020年)和72.4 mm(2020—2021年)。供试品种为‘马兰1号’。研究采用2因素随机区组试验,分别设置3个播期(sowing date, SD)和2个行距(row space, RS),共计6个处理。其中,3个播期包括1个适播期(SD1)和2个推迟播期(SD2和SD3)处理;行距包括15.0 cm等行距(15.0RS)和7.5 cm等行距(7.5RS),15.0 cm等行距播种为该地区生产常规对照。每处理4次重复。每小区面积65 m2(10 m×6.5 m)。SD1处理在2019—2021年均为10月8日播种,苗密度为3.3×106株·hm-2;SD2和SD3处理在2019—2020年为10月13日播种,2020—2021年为10月18日播种,苗密度分别为3.75×106和4.05×106株·hm-2。
前茬夏玉米收割后,秸秆粉碎返还田间。使用1GQN-230B旋耕机旋耕2次,深度为15 cm。播种前,施用120 kg·hm-2纯氮、112.5 kg·hm-2P2O5和112.5 kg·hm-2K2O。春季第1次灌溉量为120 m·3hm-2。使用Trimer Pico 64便携式土壤湿度计测量土壤含水量(体积分数)。麦田不同阶段灌溉的目标含水量为田间容量(体积分数)的80%,其中播种前灌溉的计算深度为0—40 cm土层;拔节期和开花期为0—60 cm土层。不同生长阶段的灌溉量如表1所示。
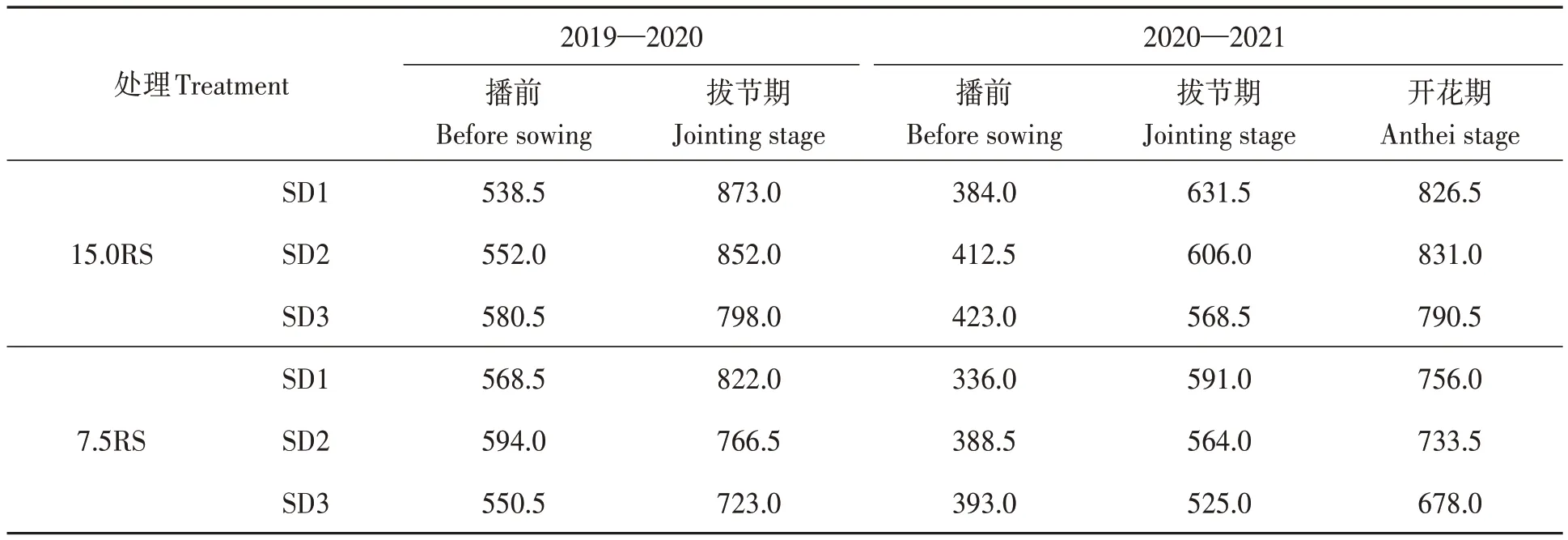
表1 不同处理下各生育期的灌水量Table 1 Irrigation amount of wheat fields under different treatments in different periods(m3·hm-2)
1.2 测定项目与方法
1.2.1 基本苗和分蘖数测定 于小麦3叶期,随机选择长势均一的3行作为调查样品段,长度为1.0 m,调查基本苗数。从3叶期开始,积温每升高60~80 ℃·d调查1次分蘖数,至越冬期(wintering period,WS)为止,分别记为D1、D2、D3…Dn、WS。以分蘖伸出叶鞘2.0 cm左右作为分蘖形成的标准,每次测定时将≥3叶的分蘖数单独计。越冬期调查的单株分蘖数记为TN。
1.2.2 生物量与内源激素含量测定 根据1.2.1中的分蘖调查时间,在每次分蘖数调查后,另选长势均匀的1行连续选取10株,去除根系,将分蘖与主茎分离后,置于120 ℃杀青30 min,烘干至恒重,测定主茎和分蘖的生物量(g)。同时选取长势具有代表性的植株5株,剪下分蘖节混匀,置于-80 ℃低温冷藏,用于内源激素含量的测定。其中,独脚金内酯(SLs)含量采用高效液相色谱-质谱联用(high performance liquid chromatographymass spectrometry,HPLC-MS)法[8]测定;吲哚乙酸(IAA)、玉米素核苷(zeatin nucleoside,ZR)、赤霉素(GA)、细胞分裂素(CTK)和油菜素甾醇(brassinosterol,BR)含量采用高效液相色谱(HPLC)法[16,18-19]测定;蔗糖(saccharose,SA)含量采用分光光度酶联法[20-21]测定。
1.2.3 缩行匀株条件下小麦分蘖节激素、蔗糖与分蘖数量的关联特征分析 以小麦冬前分蘖数(tillers number,TN)为因变量(Y),以冬前各处理SA(X1)、BR/CTK(X2)、BR/SLs(X3)、CTK(X4)、BR(X5)、SLs/GA(X6)、GA(X7)、IAA/ZR(X8)、ZR(X9)、IAA(X10)和SLs(X11)为自变量,将线性回归拟合自变量对因变量的直接通径系数(direct path coefficient, DPC)作为直接贡献效应;并进一步通过2个自变量之间的相关系数(r)与DPC确定间接通径系数(indirect path coefficient, IPC),作为间接贡献效应,计算公式见式(1)。
式中,IPCijy为Xi通过Xj对Y的间接贡献效应;rij为Xi与Xj的相关系数;DPCjy为Xj对Y的直接贡献效应。
1.3 数据统计与分析
使用SPSS 26.0 软件进行方差分析、T检验和多重比较,采用Pearson相关分析和线性回归模型统计自变量间的相关性及自变量对因变量的贡献效应。
2 结果与分析
2.1 缩行匀株对小麦茎(蘖)数量及生物量的影响
由表2可知,RRPE对各处理小麦基本苗数无显著影响。7.5RS处理显著增加小麦冬前的单株分蘖数和≥3叶的分蘖数,与15.0RS处理相比,平均增加29.5%和42.1%。将播期推迟后,7.5RS处理促进分蘖的效果更为明显。7.5RS处理在SD1播期下使单株分蘖数和≥3叶的分蘖数分别提高26.9%和25.0%;而在SD3播期下分别提高32.4%和58.8%。7.5RS处理在一定程度上提高了≥3叶分蘖数在总分蘖数中的占比,除2019年SD1处理外,其他处理均较15.0RS处理显著提高。在促进分蘖生长的同时,7.5RS处理的分蘖生物量占地上部生物量的比例也显著增加,平均增加19.1%和18.6%。不同播期相比,晚播处理对冬前分蘖生长及地上部生物量的促进作用更为显著。在SD1播期处理下,7.5RS处理的生物量较15.0RS处理增加26.2%;在SD2和SD3播期处理下,分别增加27.3%和36.0%。由此表明,RRPE处理可促进小麦冬前有效分蘖数、地上生物量和分蘖生物量占比。
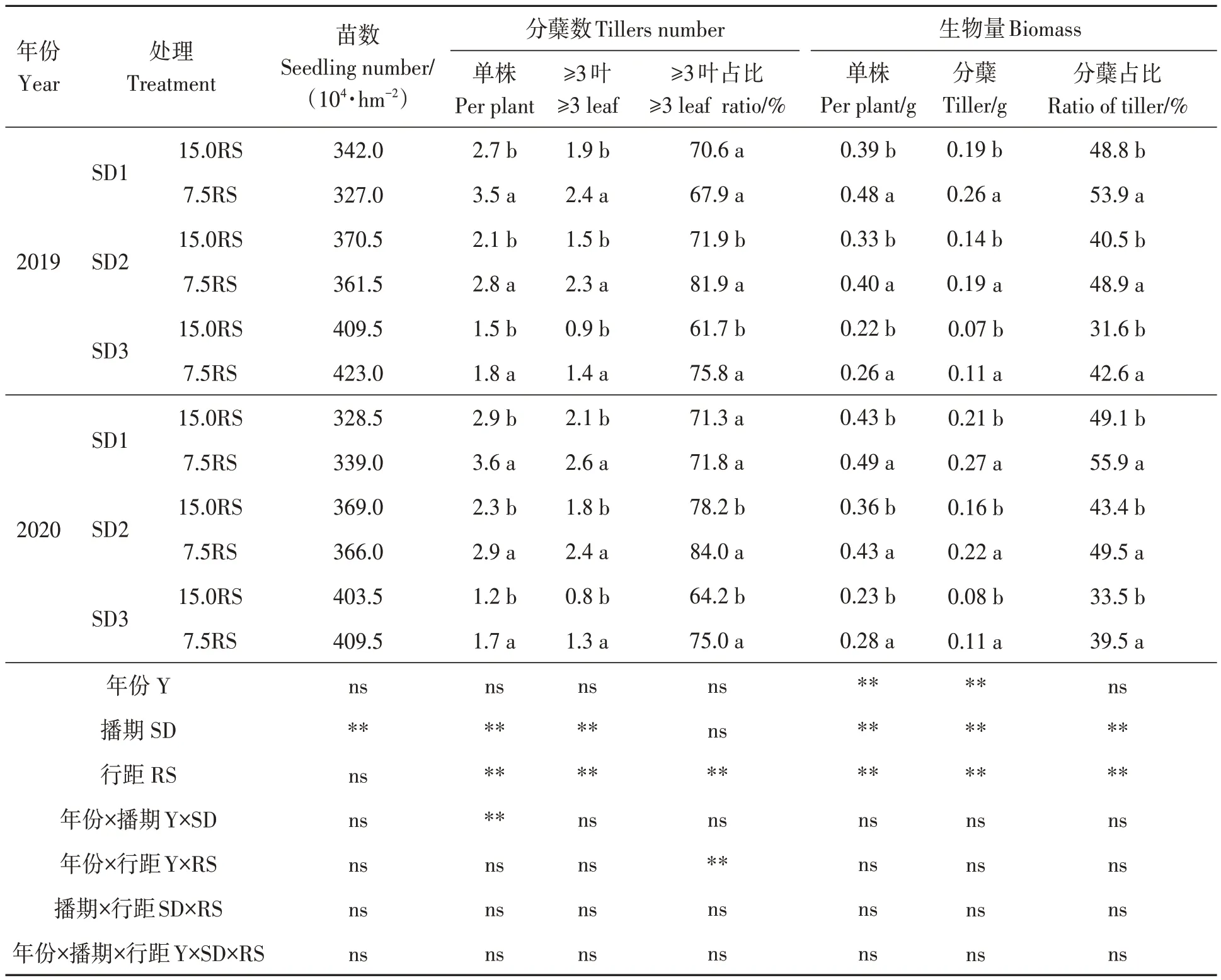
表2 不同处理下小麦的茎(蘖)数量及生物量Table 2 Tillers number and biomass under different treatments
分析不同变异来源的影响效应,结果(表2)表明,不同年份对生物量影响显著;播期和行距对分蘖数和生物量均有显著影响;且年份与播期间的互作对单株分蘖数影响显著。
2.2 缩行匀株对小麦分蘖节激素含量的影响
2.2.1 IAA和ZR含量 由图1可知,小麦分蘖节中IAA含量随生育进程的推进呈先降低后升高的趋势。SD1和SD2播期中,D3阶段前,7.5RS与15.0RS处理的IAA含量均有小幅下降趋势,且2个处理间差异较小;D3阶段之后,IAA含量均呈明显上升趋势,7.5RS处理的IAA含量平均较15.0RS处理显著降低21.6%和18.5%。SD3播期中,IAA含量在D2阶段前呈下降趋势;D2阶段后呈上升趋势。2个行距处理的IAA含量在SD1处理下D5阶段和SD3处理下的WS阶段时的差异最大,7.5RS处理较15.0RS处理平均降低29.9%和26.4%。总体看,RRPE明显降低了小麦分蘖节IAA含量,其中,SD2播期的IAA含量降幅最小,为16.5%;SD3播期的降幅最大,为23.5%。
在SD1播期的D3阶段后和SD3播期的D2阶段后,7.5RS处理明显提高了小麦分蘖节玉米素核苷(ZR)含量,较15.0RS处理平均提高16.4%。在SD2播期的D3至D4阶段,7.5RS处理的ZR含量较15.0RS处理显著提高,平均提高20.2%。由不同处理ZR含量的变化趋势可知,在3叶期,2个行距处理的ZR含量差异较小;3叶期后,处理间差异增大。2个行距处理的ZR含量峰值均出现在D3至D4阶段,且7.5RS处理的ZR含量均显著高于15.0RS处理。总体而言,小麦冬前较低的IAA含量和较高的ZR含量更利于分蘖产生,7.5RS处理下小麦分蘖节的IAA含量较15.0RS处理平均降低19.4%, ZR含量平均提高13.1%。
2.2.2 SLs和GA含量 由图2可知,在D1和D2阶段,2个行距处理的SLs含量均较低,且处理间差异较小;不同播期处理规律基本一致。2个行距处理的SLs含量均随着生育进程显著增加,其中,SD1和SD2处理的SLs含量在D4至D5阶段出现峰值;SD3处理在D3阶段出现峰值。在SLs含量峰值出现阶段,2个行距处理间的差值最大,在SD1的D5、SD2的D4和SD3的D3阶段,7.5RS处理的SLs含量较15.0RS处理分别平均提高37.0%、28.9%和22.5%。由此可见,随着播期的推迟,7.5RS处理对SLs的提升作用逐渐减弱;RRPE处理降低了小麦分蘖节SLs含量,7.5RS处理较15.0RS处理平均提高17.5%。
RRPE处理对GA的作用效应与SLs相反,3叶期至越冬期,7.5RS处理的GA含量较15.0RS处理整体下降,但不同播期的降幅存在一定差异。其中,SD1播期在D3至D5阶段降幅较大,平均降低21.2%;SD2和SD3播期在D2至D3阶段降幅较大,平均降低25.2%和21.4%。另外,在SD1的D3至D4、SD2的D2至D3、SD3的D3阶段,7.5RS处理的GA含量也显著低于15RS处理。由此表明,RRPE降低了GA含量,SD1、SD2和SD3分别降低15.7%、9.1%和9.3%,平均降低11.4%。
2.2.3 BR和CTK含量 由图3可知,与15.0RS处理相比,7.5RS处理的BR的含量在SD1播期的降幅最大,为15.0%,SD3次之,SD2降幅最小,为13.6%。其中,7.5RS处理的BR含量在SD1的D4和D5阶段降幅最大,较15.0RS处理平均降低21.4%;在SD2的D3和D4阶段,2019年降低20.5%,2020年降低29.6%;至越冬期,SD1和SD2播期中2个行距的BR含量差异不显著。在SD3播期下,2个行距处理的BR含量在D1至D2阶段无显著差异;在D3至WS阶段,7.5RS处理较15.0RS处理显著降低24.2%。推迟播期后,2个行距处理的BR含量均有不同程度降低,其中SD3播期降幅更大。由此表明,RRPE在整体上降低了小麦分蘖节BR含量,平均降低13.6%。
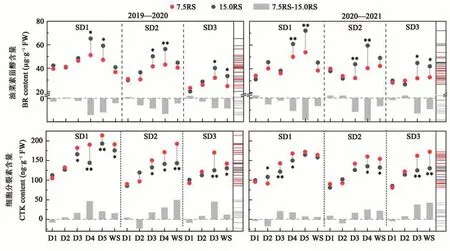
图3 不同处理下冬小麦分蘖节的油菜素甾醇和细胞分裂素含量Fig. 3 Contents of BR and CTK in wheat tillers node under different treatments
在冬前阶段,各处理CTK含量随生育进程呈增加趋势,其中,15.0RS处理在WS阶段的CTK含量较D1阶段分别平均增加57.4%(SD1)、65.0%(SD2)和40.9%(SD3);7.5RS分别增加76.7%(SD1)、92.2%(SD2)和83.4%(SD3)。对不同行距处理的CTK含量进行比较,结果(图3)表明,在D1至D2阶段,7.5RS处理与15.0RS处理差异不显著;在D3至WS阶段,SD2和SD3播期下7.5RS处理的CTK含量较15.0RS处理显著提高。整体看,RRPE在小麦分蘖初期对CTK的调控效应较小;随着植株生长、分蘖数量的增多,对CTK的促进效应逐渐增强。
2.2.4 IAA/ZR和SLs/GA 由图4可知,各处理IAA/ZR随生育进程呈先降低后升高趋势,在WS阶段达到峰值。其中,在D1阶段,7.5RS处理在3个播期的IAA/ZR均与15.0RS处理无显著差异;在D2阶段,15.0RS与7.5RS处理间的差值逐渐增大;在D3至WS阶段,7.5RS处理在SD2和SD3播期下的IAA/ZR较15.0RS处理显著降低32.9%和39.0%,在SD1播期下,2个行距处理间差异不显著。在D5至WS阶段,2个行距处理间差值(ΔIAA/ZR)较大,7.5RS处理较15.0RS处理平均降低31.3%,其中,在SD2和SD3播期下,2个行距处理间差值均在WS阶段最大;在SD1播期下2019年在D5阶段差值最大,在2020年在WS阶段差值最大。总体来看,RRPE处理降低了小麦3叶期至越冬期的IAA/ZR,且随着生育进程差异越显著。
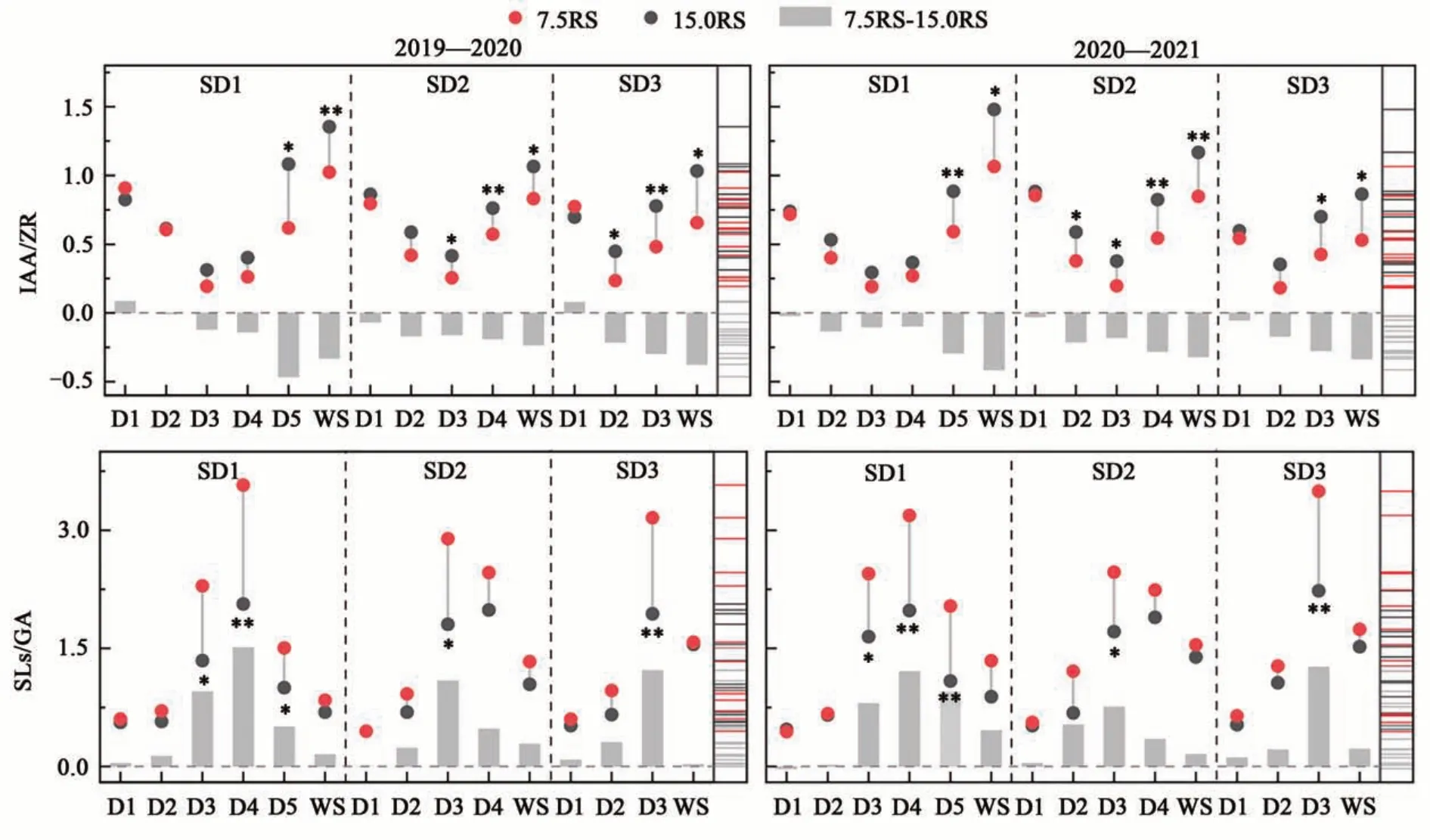
图4 不同处理下冬小麦分蘖节的IAA/ZR和SLs/GAFig. 4 IAA/ZR and SLs/GA of wheat tillers node different treatments
在D1至WS阶段,7.5RS处理的SLs/GA整体高于15.0RS处理,平均提高39.3%。其中,SD1播期下7.5RS处理对SLs/GA的提升效应最为明显,提升51.7%;SD2与SD3播期下相对较低,分别为32.1%和33.9%(图4)。进一步分析7.5RS与15.0RS处理间的差值(ΔSLs/GA)可知,随着生育进程ΔSLs/GA先升高后下降。不同播期相比,SD1的ΔSLs/GA峰值出现在D4阶段,SD2和SD3出现在D3阶段。3个播期的ΔSLs/GA变幅均存在明显的跃迁过程,其中,SD1和SD3均出现在D2至D3阶段;SD2在2019年出现在D2至D3阶段,2020年出现在D1至D2阶段。各处理在WS阶段的ΔSLs/GA较前一次明显降低,其中SD3播期下降幅最大(90.0%),SD1其次(60.9%),SD2最小(47.4%)。
2.2.5 BR/SLs和BR/CTK 由图5可知,7.5RS处理的BR/SLs整体要低于15.0RS处理,且在D1至WS阶段,2个处理间的差值(ΔBR/SLs)呈先增大后减小趋势,3个播期的ΔBR/SLs最大值均出现在冬前第2次调查,即SD1出现在D5阶段,SD2出现在D4阶段,SD3出现在D3阶段。随播期推迟,ΔBR/SLs先降低后升高,3个播期的ΔBR/SLs峰值分别为12.3(SD1)、9.2(SD2)和5.4(SD3)。在WS阶段,3个播期处理的ΔBR/SLs分别为6.6(SD1)、5.8(SD2)和3.1(SD3)。ΔBR/SLs在2个试验年度间差异较大,2019年,D1至WS阶段ΔBR/SLs均为负值,即7.5RS处理的BR/SLs低于15.0RS处理;2020年3个播期的D1阶段均出现正值,即7.5RS处理的BR/SLs高于15.0RS处理。

图5 不同处理下冬小麦分蘖节的BR/SLs和BR/CTKFig. 5 BR/SLs and BR/CTK of wheat tillers node under different treatments
2个试验年度下,7.5RS处理的BR/CTK整体低于15.0RS处理(图5)。两处理的差值(ΔBR/CTK)在D1至WS阶段呈先下降后升高趋势,峰值出现在D3至D5阶段,其中,SD1播期的峰值出现在D4(2019年)和D5(2020年)阶段;SD2播期峰值出现在D4阶段;SD3播期峰值出现在D3阶段。WS阶段,3个播期的ΔBR/CTK分别平均为0.05(SD1)、0.10(SD2)和0.11(SD3),7.5RS处理的BR/CTK低于15RS处理,其中在SD2和SD3播期下差异显著。
2.3 缩行匀株对小麦分蘖节SA含量的影响
不同处理下小麦分蘖节SA含量如图6所示。对于SD1播期,在D1至D2阶段,7.5RS处理的SA含量与15.0RS处理间差异较小;在D3阶段,两处理间的差异增大,但差异不显著;在D4阶段,7.5RS处理的SA含量较15.0RS处理显著提高26.6%;至WS阶段,两处理间的差异又减小。对于SD2播期,7.5RS处理在D4阶段与15.0RS处理间差异最大,较15.0RS处理显著提高29.5%。对于SD3播期种,2019年7.5RS处理在WS阶段与15.0RS处理间差异最大,较RS15.0处理显著提高31.0%;2020年在D3阶段差异最大,7.5RS处理较15.0RS处理显著提高25.7%。由此表明,RRPE处理有利于增加小麦分蘖节SA含量,7.5RS处理较15.0RS处理增加22.3%(SD1)、15.5%(SD2)和7.8%(SD3),平均增加15.2%。

图6 不同处理下冬小麦分蘖节的蔗糖含量Fig. 6 Content of SA in wheat tillers node under different treatments
2.4 缩行匀株条件下小麦分蘖节激素、蔗糖与分蘖数量的关联特征
2.4.1 小麦分蘖节激素、蔗糖与分蘖数量的关系 由图7可知,冬前分蘖数(TN)与蔗糖和多种激素含量密切相关。TN与SLs、ZR、BR、CTK、SA含量及SLs/GA、BR/SLs、BR/CTK呈显著正相关,其中与BR、CTK的相关系数最高,平均0.647;与BR/SLs、ZR和SA次之,相关系数为0.503~0.535。激素间也存在密切关系,ZR与SLs、ZR与SLs/GA和GA与IAA/ZR呈显著正相关,相关系数均大于0.800;GA与IAA、BR与GA、BR/SLs与IAA、BR/SLs与IAA/ZR、BR/SLs与GA、BR/CTK与IAA呈显著正相关,相关系数为0.638~0.748;SLs/GA与IAA、SLs/GA与IAA/ZR、BR/SLs和SLs/GA、BR/CTK与SLs/GA、SLs与IAA、SLs与AAI/ZR呈显著负相关,相关系数为0.405~0.588;SA与CTK、SLs/GA、ZR与SLs呈显著正相关,相关系数为0.431~0.915。
2.4.2 缩行匀株条件下小麦分蘖节激素、蔗糖对分蘖数量的贡献效应 小麦分蘖节激素、蔗糖对TN的贡献效应如图8所示。GA和BR/CTK对TN有较大的负向效应,其DPC分别为-1.35和-0.96;BR对TN的正向调节效应较高,其DPC为1.03;其他参数对TN的直接贡献效应较小,DPC为-0.18~0.55。BR/SLs通过BR对TN的正向间接贡献效应较高,其IPC为1.01;GA通过IAA/ZR和IAA对TN的负向间接贡献效应较高,其IPC分别为-1.13和-1.01;BR/SLs通过GA对TN的负向间接贡献效应也较高,其IPC为-0.97;BR/CTK通过BR、BR/SLs、IAA和GA对TN的负向间接贡献效应也较高,其IPC为-0.78~-0.64)。由此可见,单一因子对TN的贡献效应中,BR对TN的正向贡献最高;GA和BR/CTK对TN的负向贡献较高;IAA/ZR通过GA、IAA通过GA对TN的负向贡献较高。

图8 缩行匀株处理下小麦分蘖节激素、蔗糖对分蘖数的贡献效应Fig. 8 Contribution effects of phytoormones and SA in tillers node on tillers number under RRPE treatment
3 讨论
3.1 缩行匀株小麦分蘖的影响
改变植物生存空间会影响植物生长发育及植株间的竞争程度。在禾谷类作物中改变株行距对分蘖有显著影响,因此选择最佳行距可提高作物生产力。行距过宽可能无法有效利用光、水和养分等资源;行距过窄会导致行间竞争激烈。调控行间距还能优化群体结构、增强叶片光合能力、有利于干物质分配,从而提高作物产量[22-25]。研究表明,适度缩小行距、均匀株距可以促进小麦分蘖,获得高产[24,26]。这是由于行距过宽会降低单位面积的有效穗数,同时浪费光、水和养分等资源。为解决这一问题,就需要更多的穗数和粒重来弥补。但在实际生产上,穗数不足时依靠其他产量要素弥补的效果是有限的[27]。Dwyer等[28]认为,适当缩小行距可以有效地增加叶面积指数,提高叶片对光的截获与利用,并减少杂草的侵扰。然而,过于窄小的行距又会加剧行间竞争[29],进而影响植株分蘖。
本研究表明,RRPE可显著增加冬前小麦分蘖数和≥3叶的分蘖数,7.5RS处理较15.0RS处理分别平均增加29.5%和42.1%;推迟播期,RRPE对单株分蘖和≥3叶分蘖数的促进效应更加明显。7.5RS处理在促进地上部分蘖生长的同时,也显著增加了分蘖生物量在地上部生物量中的占比,较15.0RS处理平均增加19.1%和18.6%,与前人研究结果一致。本研究表明,推迟播期、缩小行距对小麦分蘖的促进效应更加明显。这对海河平原晚播麦田具有重要意义。适播期播种,积温充足,常规行距下小麦植株可以形成较多的分蘖,此时RRPE处理的优势不明显;播期推迟,积温减少,小麦叶片较少,可能会导致某种特殊波段的光对小麦茎基部的透过率增加[25,30],此时RRPE处理有利于增加小麦分蘖,提高单位面积有效穗数。RRPE处理是否与品种特性有关,尤其是分蘖与抗寒特性,还有待进一步研究。
3.2 缩行匀株对小麦激素与蔗糖的影响
植物蘖芽萌发和生长受多种因素调控,包括激素、碳水化合物水平及环境因素等[31-34]。研究显示,IAA、CTK、SLs以及碳水化合物都可能是参与植物分蘖的信号,它们单一或组合共同调控植物分蘖[35-37]。SA在激活分蘖生长方面发挥双重作用。一方面,SA起着营养作用[38];另一方面,SA可能是分蘖起始的信号分子[39]。研究表明,Palatinose是一种不可代谢的蔗糖类似物,可触发蘖芽的生长[40]。另外,SA对分蘖的调控还可能是通过与IAA、CTK和SLs联合作用,共同调节腋芽发育。但这种联合作用的通路及机制还有待进一步研究 。研究表明,BR是参与调节分蘖数的重要激素之一,BR和SLs在D53-OsBZR1复合物控制下通过FC1(TB1)拮抗调节水稻分蘖[41-42]。
本研究表明,RRPE处理降低了小麦3叶期至越冬期分蘖节中IAA、SLs、GA和BR含量,但显著提高了ZR、CTK和SA含量;还降低了IAA/ZR、BR/SLs和BR/CTK,但提高了SLs/GA。分析蔗糖和不同激素对小麦TN的相关性与贡献可知,CTK和BR与小麦TN相关系数较高,且BR对TN的正向贡献效应最高。由此可见,RRPE处理下,小麦分蘖节中CTK与BR含量的积极响应在小麦冬前分蘖产生中发挥重要的促进作用。本研究还发现,BR/SLs通过GA对小麦TN的间接贡献效应非常显著,与前人研究结果一致。由此表明,小麦分蘖信号的激发和调控,可能由多种因子共同作用[31-34]。RRPE处理调控小麦分蘖是否还涉及其他生理信号,以及这些信号响应的上游和下游环节如何关联,仍有待于进一步研究。
