软/硬肉葡萄果实细胞壁结构、组分及降解酶活性的变化
2023-06-12海龙飞栗温新李志谦李猛陈超阳魏红丽邹东方何畅冯建灿叶霞
海龙飞 栗温新 李志谦 李猛 陈超阳 魏红丽 邹东方 何畅 冯建灿 叶霞
摘 要:【目的】探讨葡萄果实发育成熟过程中软/硬肉果实软化与细胞壁结构、组分以及降解酶活性之间的关系。【方 法】以硬肉品种黎明无核和软肉品种灰比诺花后35、49、63、77、91 d的果实为试材,对细胞壁组分及相关降解酶活性、 细胞形态进行分析研究。【结果】在果实成熟过程中,果实硬度呈现先上升后下降的趋势,成熟时黎明无核果实硬度比 灰比诺高2.8倍;果实硬度与原果胶含量呈显著正相关,与纤维素酶活性呈显著负相关。黎明无核果实硬度与原果胶 和半纤维素含量呈显著正相关,灰比诺果实硬度则与纤维素含量呈显著正相关。随着果实成熟软化,果肉细胞逐渐表 现大小不均匀、排列不整齐的趋势;灰比诺果肉细胞壁在果实转色时开始降解,而黎明无核则基本保持稳定。【结论】葡 萄果实成熟软化过程中,原果胶和半纤维素快速降解导致果肉细胞壁破裂是葡萄果实软化的关键因素;果实细胞壁组 分降解是由多种酶共同作用的结果,其中以纤维素酶的作用较为显著。
关键词:葡萄;果实软化;细胞壁物质含量;酶活性;组织结构
中图分类号:S663.1 文献标志码:A 文章编号:1009-9980(2023)04-0690-09
葡萄是一种深受大众喜爱的水果,果肉质地柔 软、多汁,成熟后易落粒和腐烂,严重地影响货架期 和商品质量[1] 。根据葡萄果肉质地差异和耐压力强 弱,葡萄果肉硬度分为硬、中和软三种类型,其中软 肉型的果实极不耐贮藏[2] 。解析软/硬肉葡萄果实硬 度变化的生理基础和果实细胞结构变化对葡萄采后 贮藏保鲜的影响具有重要意义。
果肉质地的软硬主要与果肉细胞壁组分变化、 果肉细胞排列、大小等有关[3-4] 。果实细胞壁组分主 要由果胶、纤维素、半纤维素及少量结构蛋白等构 成。果实的软化过程伴随着这些细胞壁组分降解和 细胞壁结构的损坏[4-5] 。研究发现细胞壁组分与降解酶活性的变化紧密相关,欧洲梨、安溪柿、杧果、番 茄、香蕉、大樱桃、草莓等果实软化或采后贮藏过程 中,果胶甲酯酶(PME)、多聚半乳糖醛酸酶(PG)、果 胶酸裂解酶(PL)等酶活性升高加速了原果胶的降 解,导致果实快速软化[4,6-10] 。此外,果实质地差异与 细胞壁结构有关,黑莓[11] 、蓝莓[12] 、山楂[13] 等不同质地 的果实,细胞壁的完整性和钙含量差异是造成果实 软化和果肉质地差异的主要原因之一。
葡萄果实成熟软化主要与细胞壁组分的降解与 重构[1,14,15] 、果胶物质的解聚[15-16] 、钙桥的形成[1,14] 等有 关。软/硬肉葡萄果实质地差异的生理和细胞学基 础尚不清楚,笔者在本研究中以成熟期相近的软/硬 肉葡萄为材料,系统分析了果实成熟软化过程中细 胞壁组分和相关酶活性的变化,以期为揭示葡萄果 实软化和质地差异的生理基础提供理论依据。
1 材料和方法
1.1 植物材料
于2020年5月至2020年8月,以硬肉品种黎明 无核和软肉品种灰比诺为试材[2] ,供试葡萄果实取 自于中国农业科学院郑州果树研究所国家葡萄种质 资源圃。分别在葡萄花后21、35、49、63、77、91 d,选 取大小一致、无病虫害的果实0.5~2.0 kg。每次采样 后将果实随机分为两组,一组用于硬度等生理指标 测定,另一组用液氮速冻,置于-80 ℃超低温冰箱中 保存备用。
1.2 方法
1.2.1 果实硬度测定 每组样品分别选取20~30个 葡萄果粒,使用数显果实硬度计(艾德堡,GY4)进行 硬度测定,竖直向下匀速缓慢按压至果实发生形变, 以峰值均值作为果实硬度,结果以N表示。
1.2.2 果实细胞壁物质含量的测定 细胞壁纤维素 和半纤维素含量均使用试剂盒测定(CLL- 1- Y, BXW-1-G,科铭生物,江苏苏州)。首先进行细胞壁 物质提取,称取约0.3 g果实,先后进行匀浆、水浴、 离心、洗涤,获得粗细胞壁;然后加入1 mL的DMSO 溶液浸泡15 h去除淀粉,4000g离心10 min,沉淀干 燥后获得细胞壁物质(CWM)。纤维素和半纤维素 含量的测定方法均参考试剂盒说明书。原果胶和可 溶性果胶含量测定参照曹建康等[17] 的方法进行。
1.2.3 细胞壁降解酶活性测定 PME活性和PL活 性均使用试剂盒进行测定(PME-2-G,PL-1-G,科铭 生物,江苏苏州)。PG活性测定参照曹建康[17] 的方 法进行。纤维素酶(Cx)活性测定参照王鸿飞等[18] 的 方法进行。
1.2.4 果实细胞解剖结构观察 将新鲜采集的葡萄果 实用超薄刀片横切成5 mm厚的薄片,迅速投入FAA 溶液(无水乙醇、水、37%甲醛、冰醋酸体积比10∶7∶2∶1) 进行真空固定,并低速震荡24 h(4 ℃,50 r·min-1 )。 依次用50%、70%、85%、95%和100%的乙醇进行逐 级脱水,分别用25%和100%二甲苯透明1次和3次, 最后用25%的石蜡和75%的二甲苯进行渗蜡和石蜡 包埋。
使用石蜡切片机(RM2016,徕卡仪器,上海)连 续切制厚度为4 μm的切片,挑选完整的切片放置在 载玻片上进行烤片(60 ℃),石蜡烤化后取出常温保 存备用。将石蜡切片依次放入二甲苯和无水乙醇浸 泡2次,使用75%酒精和自来水漂洗干净;放入甲苯 胺蓝染液中染色 2~5 min,用自来水漂洗 2 次后烤 干;放入二甲苯透明10 min后,使用中性树胶封片。 最后进行显微镜镜检(Eclipse E100,尼康,日本)和 图像采集分析(DS-U3,尼康,日本)。 1.2.5 数据分析 每组试验至少 3 次重复,试验数 据分别采用Excel 2016和SPSS 22.0进行统计和相关 性分析,并使用Duncan新复极差法檢验差异显著性。
2 结果与分析
2.1 软/硬肉葡萄果实硬度变化分析
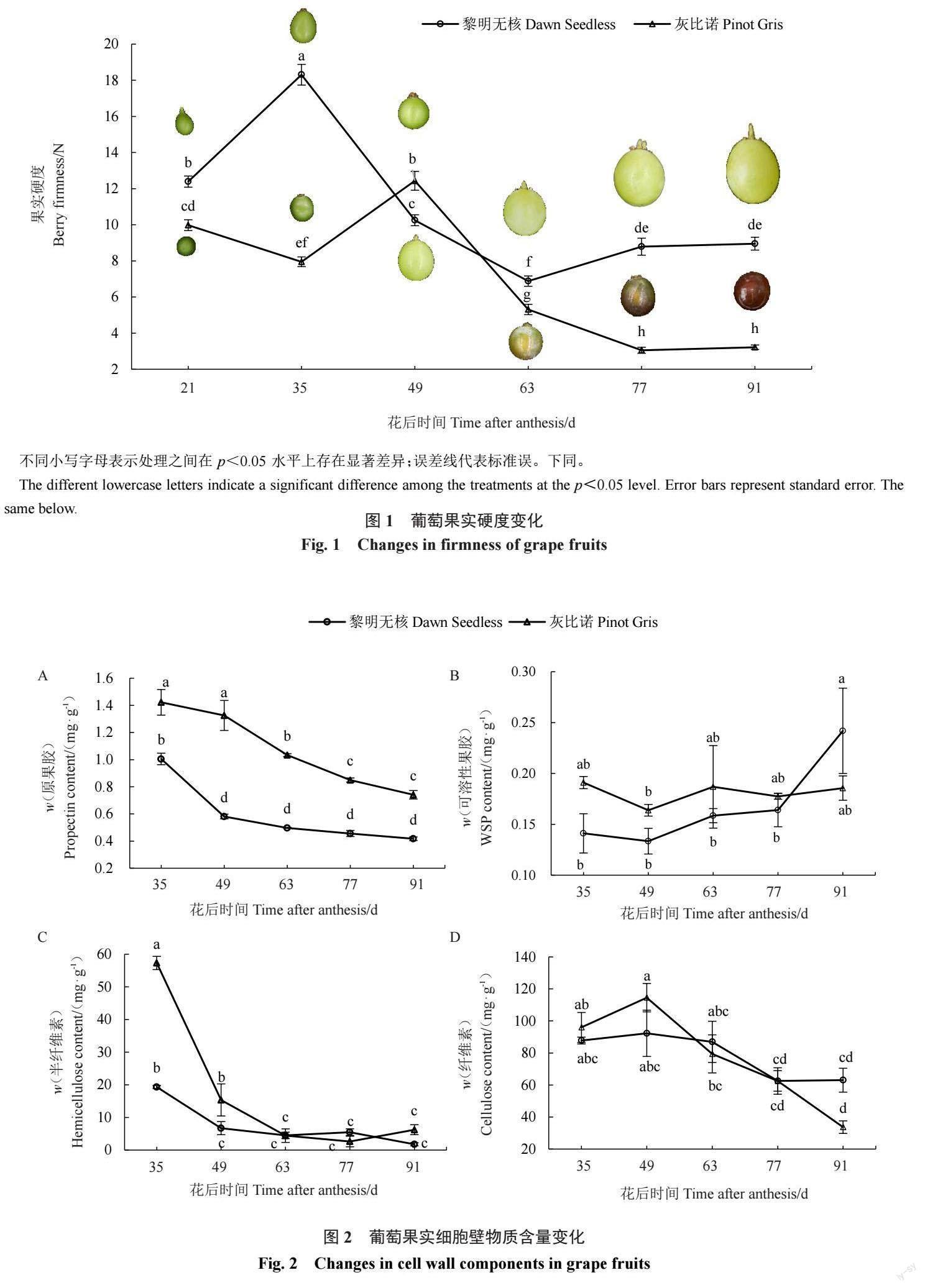
在葡萄果实的发育过程中,两个品种的果实硬 度均表现为先上升后下降的趋势(图1)。黎明无核 和灰比诺果实硬度分别在花后 35 d 和 49 d 出现峰 值,随后快速软化,花后 63 d 灰比诺果实开始转色 (图1)。在果实发育的各个时期,除花后第49天外, 黎明无核硬度均显著高于灰比诺;在果实完熟时(花 后91 d),黎明无核的果实硬度比灰比诺的果实硬度 高2.8倍。
2.2 果实细胞壁物质含量变化
在葡萄果实成熟软化过程中,两个品种的原果 胶含量总体呈下降趋势,且灰比诺的原果胶含量均 显著高于黎明无核原果胶含量(图2-A)。在黎明无 核葡萄成熟过程中,果实的原果胶含量在花后35 d 最高,然后迅速下降,但花后49~91 d含量变化差异 不显著;而 WSP 含量与原果胶的变化趋势相反, WSP含量在花后35~77 d无显著差异,而在花后91 d显著高于其他时期(图2-A,B)。灰比诺的原果胶含 量在果实成熟过程中一直呈下降趋势,而WSP含量 在各个时期均无显著差异(图2-B)。
对葡萄果实发育中半纤维素和纤维素含量分析 结果显示,半纤维素含量变化较为显著(图2-C)。黎 明无核和灰比诺的半纤维素含量均在花后35~49 d快 速下降,如灰比诺的半纤维素含量从花后 35 d 的 57.32 mg·g-1 下降到花后49 d的15.37 mg·g-1 ,降低了 73%。同时,在花后35~49 d灰比诺半纤维素含量显 著高于黎明无核的含量。在果实发育的63~91 d,两 个品种的半纤维素含量均维持较低水平且无显著差 异(图2-C)。两个品种的纤维素含量分析的结果表 明,在黎明无核果实成熟过程中,纤维素含量无显著 变化,而灰比诺纤维素含量总体呈下降趋势(图2-D)。
2.3 细胞壁降解酶活性变化
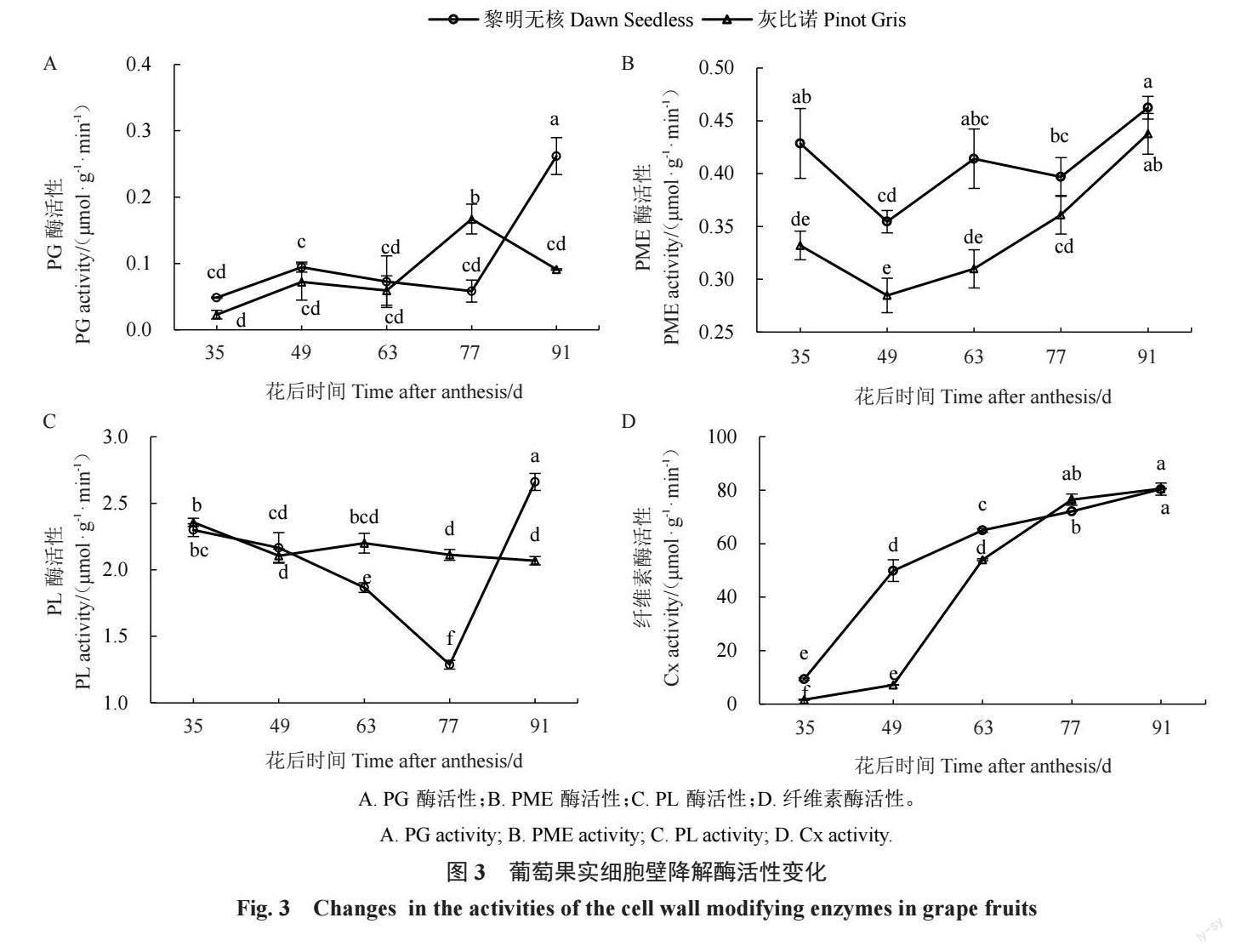
多聚半乳糖醛酸酶(PG)能将果胶主链水解为 单个半乳糖醛酸分子以降解果胶,实现果实的软 化。在果实生长发育过程中,黎明无核果实 PG 酶 活性在花后 35~77 d 无显著变化,在果实完熟时迅 速上升,达到最高。灰比诺果实 PG 酶活性呈先升 高后降低的趋势,峰值出现在花后77 d(图3-A)。 果胶甲酯酶(PME)主要催化甲基化的果胶甲酯 转化为果胶酸,去甲基后由多聚半乳糖醛酸酶(PG) 降解果胶主链。在生长发育过程中,两个品种PME 活性变化均呈现先下降后上升的趋势,在花后49 d 时 PME 活性最低。黎明无核的 PME 活性在花后 35~63 d均显著高于灰比诺,在果实发育的77~91 d, 两个品种PME酶活性无显著差异(图3-B)。 PL 活性在两个品种中也表现出不同的趋势。 在黎明无核果实中,PL活性呈现先降低再升高的趋 势,在花后77 d达到最低。在灰比诺果实中,PL活 性无显著变化(图3-C)。
纤维素酶(Cx)降解细胞壁中的纤维素,使细胞 壁结构松弛,导致果实软化。在葡萄生长发育过程 中,两个葡萄品种 Cx 酶活性均呈逐渐上升的趋势 (图3-D)。在花后35~63 d,黎明无核Cx活性均显著 高于灰比诺,而在花后77~91 d,两品种Cx活性无显 著差异(图3-D)。
2.4 生理指标的相关性分析
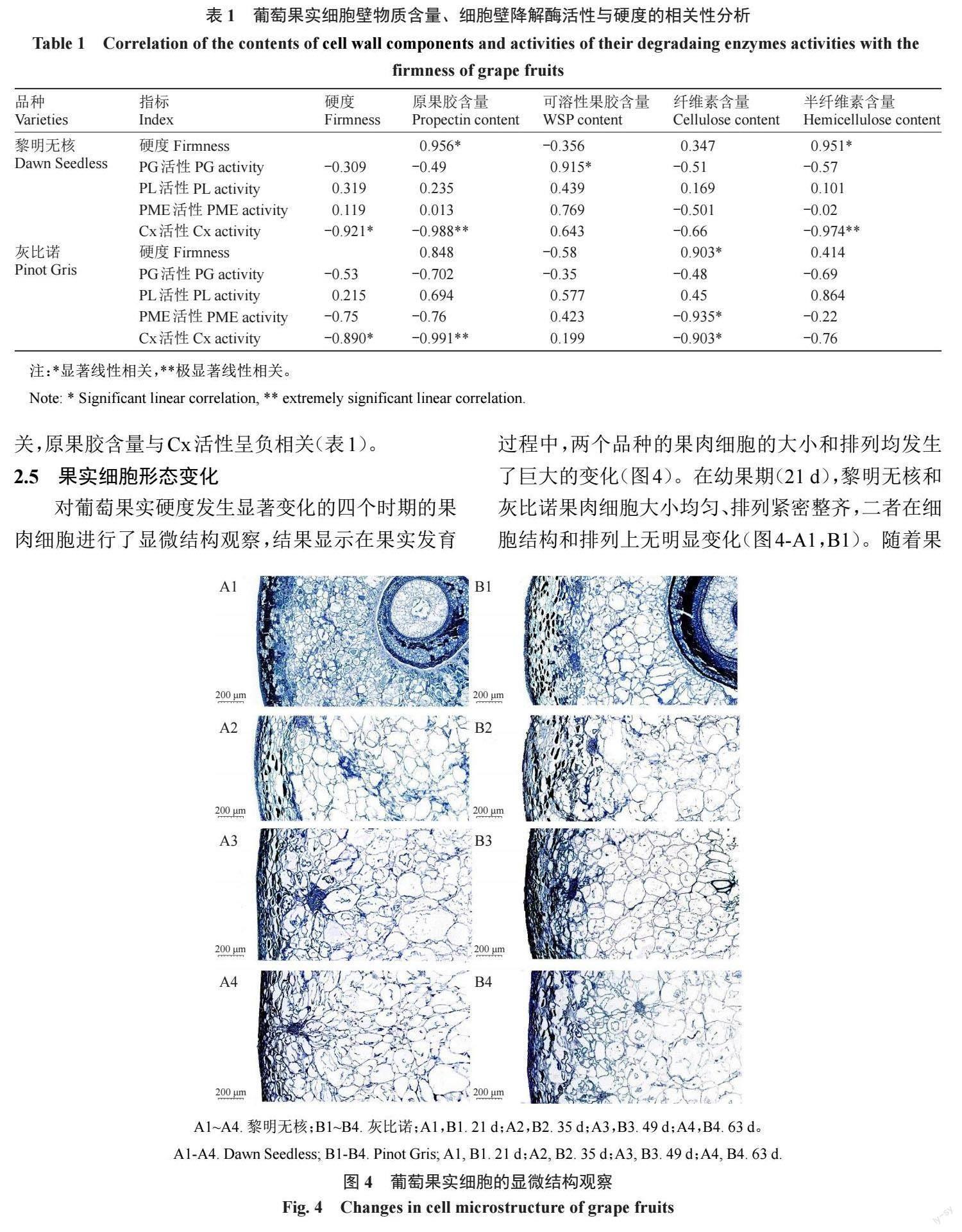
在黎明无核果实发育过程中,果实硬度的变化与原果胶和半纤维素含量呈显著正相关,与Cx活性 呈显著负相关;同时WSP含量与PG活性呈显著正 相关,原果胶和半纤维素含量与Cx活性均呈极显著 负相关(表 1)。灰比诺果实在发育过程中,硬度与 纤维素含量呈显著正相关,与Cx活性呈负相关;此 外纤维素含量与Cx活性和PME活性均呈显著负相关,原果胶含量与Cx活性呈负相关(表1)。
2.5 果实细胞形态变化
对葡萄果实硬度发生显著变化的四个时期的果 肉细胞进行了显微结构观察,结果显示在果实发育 过程中,两个品种的果肉細胞的大小和排列均发生 了巨大的变化(图4)。在幼果期(21 d),黎明无核和 灰比诺果肉细胞大小均匀、排列紧密整齐,二者在细 胞结构和排列上无明显变化(图4-A1,B1)。随着果实的发育和膨大(35~49 d),两个品种的果肉细胞明 显变大,但细胞大小不均匀、排列较松散(图4-A2, A3,B2,B3);其中灰比诺在花后49 d呈现果皮和果 肉细胞结合不紧密(图 4-B3)。在果实开始软化和 着色时(63 d),两个品种的细胞排列和大小均杂乱 无章,尤其灰比诺果肉细胞的细胞壁边界模糊,开始 出现大量降解现象;相比较而言,黎明无核的细胞壁 基本保持完整,未出现细胞壁降解现象(图4-A4)。
3 讨 论
3.1 原果胶、纤维素及半纤维素的降解导致葡萄果 实软化
纤维素是果实细胞壁的骨架,半纤维素与纤维 素形成交联结构,共同维持细胞骨架的稳定。在苹 果和梨果实成熟过程中,纤维素含量与秦冠的硬度 变化呈显著相关,而半纤维素含量与富士苹果和库 尔勒香梨硬度呈极显著相关[22-23] 。在樱桃果实中,纤 维素降解速率不同是导致质地差异的主要因素之 一[10] 。巨峰葡萄果实软化过程中,果胶和半纤维素的 降解、半纤维素和纤维素含量的降低是前期软化的 主要原因之一[16] 。硬肉红地球和软肉玫瑰香的成熟 软化与原果胶和可溶性果胶含量密切相关[15] 。不溶 性的原果胶降解为可溶性的果胶和果胶酸,引起胞 间层溶解和细胞壁结构破坏,导致果实软化[19-20] 。在 大樱桃的果实成熟软过程中,硬度高、耐贮藏品种可 溶性果胶含量低,软化速度慢,而硬度低,不耐贮藏 的品种可溶性果胶含量高,软化速度快[10] 。在桃果实 成熟过程中共价紧密结合型果胶转变为离子可溶性 果胶,软溶质品种下降速率明显高于硬溶质品种,细 胞壁多糖的降解速度亦存在明显差异[21] 。本研究结果 显示,在细胞壁的结构物质中,原果胶、半纤维素和 纤维素含量均与软/硬肉葡萄果实软化有关,尤其是原 果胶和半纤维素含量在两个品种中差异较显著,这进 一步表明了葡萄果实的软化与原果胶和半纤维素降 解均有关系,是细胞壁组分解聚综合作用的结果。
3.2 葡萄细胞壁降解酶活性分析
与果实细胞壁组分降解中最为关键的水解酶是 PME、PG和PL等,它们协同作用于果胶的降解,Cx 和木葡聚糖内糖基转移酶(XTH)分别参与纤维素 和半纤维素的水解。在苹果早熟品种(系)中,PME 酶活性呈先上升后下降又上升的趋势[24] 。在甜樱桃 生长发育过程中,PME与美早和红灯硬度呈显著负 相关,而软肉品种佳红 PME 活性几乎没有变化[10] 。 在葡萄果实发育中,硬肉NN107的PME活性比软肉 的Thompson Seedless高,且在果实完熟前活性无显 著变化[1] 。本研究的结果同样显示在果实转色前, 硬肉品种黎明无核的PME活性均比软肉品种灰比 诺高,预示 PME 活性变化可能不是造成软/硬肉葡 萄果实质地差别的主要原因。
PG 将去酯化的果胶分子降解为半乳糖醛酸, PL通过β-消除方式随机断裂多聚半乳糖醛酸来降 解细胞壁。PG活性在芒果和柿子的成熟软化过程 中呈上升趋势,在猕猴桃和苹果呈先上升后下降趋 势[25- 27] ,在樱桃中,PG 可能是硬度变化的次要原 因[10] 。在香蕉成熟过程中,PL酶活性的高峰与呼吸 跃变高峰相一致,跃变之后,PL酶活性逐渐降低[6] 。 沉默表达番茄 SlPL 基因显著降低果实可溶性果胶 含量、提高硬度,由此认为PL是番茄果实软化的关 键因子之一[9] 。本研究结果显示,在葡萄果实的成 熟软化过程中,两个品种的PG和PL活性变化差异 不显著,且与果实硬度的相关性不显著,这表明葡萄 果实中果胶降解可能是PME、PG、PL等降解酶综合 作用的结果。
木葡聚糖转移酶和纤维素酶分别是半纤维素与 纤维素降解的关键酶,在果实的软化过程中起着重 要的作用。在柿果实成熟和软化过程中,木葡聚糖 转移酶关键基因DkXTH8 的表达量最高,过表达DkXTH8 基因导致了番茄细胞致密性变差、果实软 化加速[28] 。在大樱桃软化过程,Cx活性并没有影响 到果实硬度[10] 。但在本实验中发现,软硬肉葡萄品 种的Cx活性均与果实的硬度呈显著相关,这表明纤 维素酶活性变化可能是引起葡萄果实软化的重要因 素之一。
3.3 果肉细胞壁结构的完整性影響了果实硬度的 变化
随着果实成熟软化,原果胶降解,侧链中性糖损 失,细胞壁发生不可逆转的变化,最终导致细胞的破 裂[4,29-31] 。在番茄果实成熟过程中,抑制细胞壁多糖 解聚,保持细胞壁结构完整,可抑制成熟软化进 程[32] 。果胶结构完整的樱桃具有较高果实硬度[33] 。 耐压力强的红地球、秋红、秋黑果肉细胞大且分布均 匀、紧密,而易软化的巨峰果肉细胞大小不均匀、松 散[3] ;同时,硬肉品种NN107的细胞壁解聚率较低, 也是不易软化的重要原因[1] 。笔者在本研究中发现果实开始软化和着色时(63 d),硬肉品种黎明无核 的细胞壁结构保持完整,而软肉品种灰比诺已经部 分降解;同时发现灰比诺转色前半纤维素含量急剧 下降,这表明果肉细胞壁组分降解速度和维持细胞 壁完整性是影响葡萄果肉质地的重要因素。
4 结 论
在葡萄果实成熟软化过程中,原果胶和半纤维 素快速降解,导致了果肉细胞壁的破裂是葡萄果实 软化的关键因素;果实细胞壁组分的降解是由多种 酶共同作用的结果,其中以纤维素酶的作用较为显 著。
参考文献 References:
[1] BALIC I,EjSMENTEWICZ T,SANHUEZA D,SILVA C, PEREDO T,OLMEDO P,BARROS M,VERDONK J C, PAREDES R,MENESES C,PRIETO H,ORELLANA A,DE FILIPPI B G,CAMPOS-VARGAS R. Biochemical and physio logical study of the firmness of table grape berries[J]. Posthar vest Biology and Technology,2014,93:15-23.
[2] 姜建福 . 葡萄果肉质地性状的评价、QTL 定位及候选基因预 测[D]. 杨凌:西北农林科技大学,2020.
JIANG Jianfu. Evaluation,QTL analysis and candidate gene pre diction for berry texture inVitis vinifera L.[D]. Yangling:North west A & F University,2020.
[3] 周会玲,李嘉瑞 . 葡萄浆果耐压力、耐拉力与果实结构的关系[J]. 西北农林科技大学学报(自然科学版),2007,35(2):106-109.
ZHOU Huiling,LI Jiarui. The relationship between fruit struc ture with pressure and pulling force of berry of grapes[J]. Jour nal of Northwest A&F University (Natural Science Edition), 2007,35(2):106-109.
[4] WANG D,YEATS T H,ULUISIK S,ROSE J K C,SEYMOUR G B. Fruit softening:Revisiting the role of pectin[J]. Trends in Plant Science,2018,23(4):302-310.
[5] AIRIANAH O B,VREEBURG R A M,FRY S C. Pectic poly saccharides are attacked by hydroxyl radicals in ripening fruit: evidence from a fluorescent fingerprinting method[J]. Annals of Botany,2016,117(3):441-455.
[6] PAYASI A,SANWAL G G. Pectate lyase activity during ripen ing of banana fruit[J]. Phytochemistry,2003,63(3):243-248.
[7] 莊军平,苏菁,陈维信 . 香蕉果实果胶裂解酶基因 cDNA 克隆 及序列分析[J]. 果树学报,2006,23(2):227-231.
ZHUANG Junping,SU Jing,CHEN Weixin. Cloning and se quence of cDNA fragments encoding pectate lyase genes from banana fruit[J]. Journal of Fruit Science,2006,23(2):227-231.
[8] 周厚成 . 草莓果实成熟软化相关基因的研究[D]. 杨凌:西北农 林科技大学,2014.
ZHOU Houcheng. Research on genes related to fruit ripening and softening of strawberry[D]. Yangling:Northwest A & F Uni versity,2014.
[9] YANG L,HUANG W,XIONG F J,XIAN Z Q,SU D D,REN M Z,LI Z G. Silencing ofSlPL,which encodes a pectate lyase in tomato,confers enhanced fruit firmness,prolonged shelf-life and reduced susceptibility to grey mould[J]. Plant Biotechnolo gy Journal,2017,15(12):1544-1555.
[10] 沈颖,李芳东,王玉霞,张序,李延菊,赵慧,张福兴 . 甜樱桃果 实发育过程中细胞壁组分及其降解酶活性的变化[J]. 果树学 报,2020,37(5):677-686.
SHEN Ying,LI Fangdong,WANG Yuxia,ZHANG Xu,LI Yan ju,ZHAO Hui,ZHANG Fuxing. A study on the variation of cell wall components and activities of their degradation enzymes in sweet cherry during fruit development[J]. Journal of Fruit Sci ence,2020,37(5):677-686.
[11] 熊振豪,张春红,李维林,吴文龙 . 不同硬度黑莓果实发育成熟 进程中的解剖构造[J]. 南京林业大学学报(自然科学版), 2019,43(1):149-153.
XIONG Zhenhao,ZHANG Chunhong,LI Weilin,WU Wenlong. Anatomical structure characteristics of blackberry fruits with dif ferential firmness during ripening[J]. Journal of Nanjing Forest ry University (Natural Science Edition),2019,43(1):149-153.
[12] MONTECCHIARINI M L,SILVA-SANZANA C,VALDERRA MO L,ALEMANO S,GOLLAN A,RIVADENEIRA M F,BEL LO F,VAZQUEZ D,BLANCO-HERRERA F,PODESTA F E, TRIPODI K E J. Biochemical differences in the skin of two blue berries (Vaccinium corymbosum) varieties with contrasting firm ness:Implication of ions,metabolites and cell wall related pro teins in two developmental stages[J]. Plant Physiology and Bio chemistry,2021,162:483-495.
[13] XU J Y,ZHAO Y H,ZHANG X,ZHANG L J,HOU Y L, DONG W X. Transcriptome analysis and ultrastructure observa tion reveal that hawthorn fruit softening is due to cellulose/hemi cellulose degradation[J]. Frontiers in Plant Science,2016,7: 1524.
[14] ZEPEDA B,OLMEDO P,EjSMENTEWICZ T,SEPULVEDA P,BALIC I,BALLADARES C,DELGADO- RIOSECO J, FUENTEALBA C,MORENO A A,DEFILIPPI B G,MENE SES C,PEDRESCHI R,CAMPOS-VARGAS R. Cell wall and metabolite composition of berries ofVitis vinifera (L.) cv. Thompson Seedless with different firmness[J]. Food Chemistry, 2018,268:492-497.
[15] MA L,SUN L J,GUO Y S,LIN H,LIU Z D,LI K,GUO X W. Transcriptome analysis of table grapes (Vitis vinifera L.) identi fied a gene network module associated with berry firmness[J]. PloS One,2020,15(8):e0237526.
[16] YAKUSHIJI H,SAKURAI N,MORINAGA K. Changes in cell wall polysaccharides from the mesocarp of grape berries during veraison[J]. Physiologia Plantarum,2010,111(2):188-195.
[17] 曹建康,姜微波,趙玉梅 . 果蔬采后生理生化实验指导[M]. 北京:中国轻工业版社,2007:79-88.
CAO Jiankang,JIANG Weibo,ZHAO Yumei. Experimental in struction of postharvest physiology and biochemistry of fruits and vegetables[M]. Beijing:China Light Industry Press,2007: 79-88.
[18] 王鸿飞,邵兴锋 . 果品蔬菜贮藏与加工实验指导[M]. 北京:科 学出版社,2012:56-59.
WANG Hongfei,SHAO Xingfeng. Experimental guidance for storage and processing of fruit and vegetables[M]. Beijing:Sci ence Press,2012:56-59.
[19] COSGROVE D J. Growth of the plant cell wall[J]. Nature Re views Molecular Cell Biology,2005,6(11):850-861.
[20] ROLLE L,SIRET R,SEGADE S R,MAURY C,GERBI V, JOURJON F. Instrumental texture analysis parameters as mark ers of table-grape and winegrape quality:A review[J]. American Journal of Enology and Viticulture,2012,63(1):11-28.
[21] 阚娟 . 不同溶质型桃果实成熟软化机理研究[D]. 扬州:扬州大 学,2011.
KAN Juan. Study on the mechanism of peach ripening and soft ening with different solutes[D]. Yangzhou:Yangzhou Universi ty,2011.
[22] 雷琴,任小林 . 秦冠和富士苹果果实成熟过程中的质地变化 特性[J]. 西北农业学报,2007,16(1):213-216.
LEI Qin,REN Xiaolin. Characteristics of texture change with Qinguan and Fuji apples during ripening[J]. Acta Agriculturae Boreali-occidentalis Sinica,2007,16(1):213-216.
[23] 陈湘颖,丁想,牛莹莹,尤璐瑶,廖康 ‘. 库尔勒香梨及其芽变 品种‘沙 01果实硬度差异的相关因素分析[J]. 西南农业学 报,2020,33(5):952-957.
CHEN Xiangying,DING Xiang,NIU Yingying,YOU Luyao, LIAO kang. Analysis on related factors of difference of fruit firmness between‘Korla Fragrant Pearand its bud mutation ‘Sha 01[J]. Southwest China Journal of Agricultural Sciences, 2020,33(5):952-957.
[24] 李世军,尹宝颖,李佳,李中勇,张学英,徐继忠 . 早熟苹果果 实软化过程中乙烯、相关酶及其基因表达变化[J]. 华北农学 报,2019,34(6):104-109.
LI Shijun,YIN Baoying,LI Jia,LI Zhongyong,ZHANG Xueying,XU Jizhong. Changes of ethylene,related enzymes and their genes expression during softening of premature apple fruits[J]. Acta Agriculturae Boreali-Sinica,2019,34(6):104-109.
[25] HUANG W J,CHEN M Y,ZHAO T T,HAN F,ZHANG Q, LIU X L,JIANG C Y,ZHONG C H. Genome-wide identifica tion and expression analysis of polygalacturonase gene family in kiwifruit (Actinidia chinensis) during fruit softening[J]. Plants, 2020,9(3):327.
[26] LI L,LI C B,SUN J,SHENG J F,ZHOU Z G,XIN M,YI P, HE X M,ZHENG F J,TANG Y Y,LI J M,TANG J. The effects of 1-Methylcyclopropene in the regulation of antioxidative sys tem and softening of mango fruit during storage[J]. Journal of Food Quality,2020,2020:6090354.
[27] WANG H,CHEN Y H,LIN H T,LIN M S,CHEN Y H,LIN Y F. 1-Methylcyclopropene containing- papers suppress the disas sembly of cell wall polysaccharides in Anxi persimmon fruit dur ing storage[J]. International Journal of Biological Macromole cules,2020,151:723-729.
[28] HAN Y,BAN Q Y,LI H,HOU Y L,JIN M J,HAN S K,RAO J P. DkXTH8,a novel xyloglucan endotransglucosylase/hydrolase in persimmon,alters cell wall structure and promotes leaf senes cence and fruit postharvest softening[J]. Scientific Reports, 2016,6:39155.
[29] TOIVONEN P M A,BRUMMELL D A. Biochemical bases of ap pearance and texture changes in fresh-cut fruit and vegetables[J]. Postharvest Biology and Technology,2008,48(1):1-14.
[30] CHRISTIAENS S,BUGGENHOUT S V,HOUBEN K,CHAU LA D,LOEY A M V,HENDRICKX M E. Unravelling process induced pectin changes in the tomato cell wall:An integrated ap proach[J]. Food Chemistry,2012,132(3):1534-1543.
[31] POS? S,PANIAGUA C,MATAS A J,GUNNING A P,MOR RIS V J,QUESADA M A,MERCADO J A. A nanostructural view of the cell wall disassembly process during fruit ripening and postharvest storage by atomic force microscopy[J]. Trends in Food Science and Technology,2019,87:47-58.
[32] XIE F,YUAN S Z,PAN H X,WANG R,CAO J K,JIANG W B. Effect of yeast mannan treatments on ripening progress and modification of cell wall polysaccharides in tomato fruit[J]. Food Chemistry,2017,218:509-517.
[33] ZHANG L F,CHEN F S,AN H J,YANG H S,SUN X Y,GUO X F,LI L T. Physicochemical properties,firmness,and nano structures of sodium carbonate-soluble pectin of 2 Chinese cher ry cultivars at 2 ripening stages[J]. Journal of Food Science, 2008,73(6):N17-N22.
