云南9种常见菊科杂草叶片性状研究
2023-06-10浦艳飞宋起萱汪国鲜杨春梅吴丽芳余蓉培王继华
浦艳飞, 宋起萱, 汪国鲜, 杨春梅, 吴丽芳, 余蓉培, 王继华,4
(1.云南大学资源植物研究院,昆明 650504;2.云南省农业科学院花卉研究所,昆明 650205;3.玉溪云星生物科技有限公司, 云南 江川 653100;4.国家观赏园艺工程技术研究中心,昆明 650205)
【研究意义】中国杂草90科571属,共1454种,是影响中国农业生产的关键问题之一[1]。仅在昆明地区,常见农田杂草就达151种,其中菊科杂草有31种[2]。中国常年受杂草危害的土地面积超过0.74×108hm2,每年在杂草防治方面投入的费用为235×108元左右,直接导致的经济损失超过1000亿[3]。菊科杂草因其种子数量多,繁殖能力强[4],且普遍具有化感作用,通过释放化感物质来抑制周围物种的生长[5],同时,菊科植物具有较强的光合适应性,对干旱贫瘠生境有着极强的耐受力,因此对土地资源具有极强的侵占特性,严重破坏生态平衡[4]。开展云南常见菊科杂草叶片性状研究将为菊科杂草的适应性研究以及防治策略提供参考依据。【前人研究进展】目前,中国主要使用化学除草剂来进行杂草防治,但是,大量使用化学除草剂带来了严重的环境污染问题和食品安全问题。随着人们食品安全意识和环境保护意识增强,生物防治已经成为杂草防控的重要趋势和研究热点。了解杂草的生长习性及生理生态特征是进行生物防治的前提[6]。植物叶片具有生长周期短、更新快、数量多、对环境变化敏感等特点,是表征植物生长习性、生理生态特征以及生长对策权衡的重要指标[7-8]。青藏高原地区的火绒草、蒲公英和美丽凤毛菊等菊科植物的角质层和表皮层厚度随海拔升高而增加以适应高原低温、强辐射的环境[9]。科尔沁沙地的菊科植物为适应干旱环境,保持水分,则采取低比叶面积、高叶干物质含量等适应策略[10]。同时,植物对环境的不同适应策略会导致其化学计量特征和养分利用策略各有不同[11]。在魏海霞等[12]对毛乌素沙漠主要植物叶片性状的研究中显示,固定沙地的中间锦鸡儿具有较高的叶片氮(N)含量,其通过提高氮含量来保持较高的水分利用率,以适应沙地干旱环境。此外,叶绿素荧光作为光合作用探针,会随外界环境发生改变,亦能反映植物对环境的适应性[13-14]。在对华南地区几种菊科杂草的研究中发现,三裂叶蟛蜞菊(Wedeliatrilobata)和白花鬼针草(Bidensalba)的PSII最大光化学效率(Fv/Fm)分别高于蟛蜞菊(W.chinensis)和鬼针草(B.pilosa),具有较强的光合能力,表现出对当地环境较好的适应性[15]。【本研究切入点】蒲公英(TaraxacummongolicumHand-Mazz.)、苦苣菜(SonchusoleraceusL.)、鼠麴草(GnaphaliumaffineD. Don.)、青蒿(ArtemisiaannuaL.) 4个种属于中国土著种菊科杂草[16-18]。紫茎泽兰(EupatoriumcoelestinumMichx.)、银胶菊(PartheniumhysterophorusL.)、小蓬草(ConyzacanadensisL.)、三叶鬼针草(BidenspilosaL.)、牛膝菊(GalinsogaparvifloraCav.) 5个种属于菊科入侵种杂草[4]。上述9种菊科杂草在云南田间广泛分布,影响农作物生长[2, 4],其中,紫茎泽兰、三叶鬼针草、和银胶菊为云南Ⅰ级恶性入侵杂草[3-4],牛膝菊为Ⅱ级严重入侵杂草[4],均具有严重危害性[3]。但目前尚未见针对上述云南常见菊科杂草叶片性状的比较分析。【拟解决的关键问题】以上述9种云南常见菊科杂草为试验材料,对叶片解剖结构、气孔密度、叶脉密度、干物质含量、比叶面积、化学计量特征、叶绿素含量、叶绿素荧光参数等叶片性状进行比较分析,并对其适应性机理进行初步探究,研究结果为菊科杂草的防治提供重要理论依据。
1 材料与方法
1.1 供试材料
本研究以云南常见菊科杂草蒲公英(T.mongolicumHand. Mazz.)、苦苣菜(S.oleraceusL.)、鼠麴草(G.affineD. Don.)、青蒿(A.annuaL.)、紫茎泽兰(E.coelestinumMichx.)、银胶菊(P.hysterophorusL.)、小蓬草(C.canadensisL.)、三叶鬼针草(B.pilosaL.)、牛膝菊(G.parvifloraCav.)为试验材料。试验植株采自云南省农业科学院花卉研究所附近生境一致的荒坡(25°7′22′′ N,102°45′59′′ E),该研究区域气候干燥,昼夜温差大,年均温度13.5~16.8 ℃,年日照2200 h,无霜期285 d,年均降水量1006.5 mm[19]。2021年4月每个种分别选取3株长势一致的植株,每株选取中部3片成熟叶片进行相应指标的测定。
1.2 试验方法
1.2.1 叶片解剖结构测定 对叶片主脉两侧组织进行徒手切片,用光学显微镜观察并对叶片横切面进行拍照,利用软件ImageJ分析得到叶片厚度(Leaf thickness, LT)、角质层厚度(Cuticle thickness, CT)、表皮厚度(Epidermis thickness, ET)、栅栏组织厚度(Palisade parenchyma thickness, PT)、海绵薄壁组织厚度(Spongy parenchyma thickness, ST)[20]。
栅海比(P/S) =PT/ST
组织结构紧密度(Cell tense ratio, CTR)(%) =PT/LT×100
组织结构疏松度(Sponge ratio, SR)(%) =ST/LT×100
1.2.2 气孔密度测定 在叶片背面中部(避开主脉)用指甲油涂抹1 cm2区域,待指甲油干后,用镊子撕下成膜部分,制片观察并拍照。利用软件ImageJ对拍摄得到的气孔图片进行分析处理,得出单位面积内的气孔数目即气孔密度(Stomatal density, SD)[20]。
1.2.3 叶脉密度测定 剪取1 cm2叶片,参照张晓燕等[20]方法进行处理和制片,随后用体式镜观察并拍照,用软件ImageJ测量视野内叶脉的总长度,计算单位叶面积叶脉的总长度,即叶脉密度(Vein density, VD)。
1.2.4 叶片干物质含量和比叶面积测定 称量待测叶片得到鲜重,并对叶片进行拍照,使用软件ImageJ对照片进行分析,得到叶面积大小。随后,将叶片置于65 ℃烘箱烘至恒重,称取干重。按照以下公式计算叶片的干物质含量(Leaf dry matter content, LDMC)和比叶面积(Specific leaf area, SLA)。


1.2.5 全碳、全氮、全磷测定 将长势均一的成熟叶片分别装于信封袋内,65 ℃烘干至恒重,粉碎后用于全碳(C)、全氮(N)、全磷(P)的测定。利用滴定法以硫酸亚铁滴定测定全碳含量,凯氏定氮法测定全氮含量,钼锑抗比色法测定全磷含量[21]。每个种分别3次重复。
1.2.6 叶绿素含量测定 将各材料离体叶片剪碎,采用N, N-二甲基甲酰胺浸提,获得叶绿素提取液,分别于波长663和645 nm下测定OD值,结合Arnon公式计算各材料的叶绿素a含量(Chlorophyll a content, Chl a)、叶绿素b含量(Chlorophyll b content, Chl b)、叶绿素总含量(Total chlorophyll content, Total Chl)以及叶绿素a/叶绿素b(Chlorophyll a content/Chlorophyll b content, Chl a/Chl b)[22]。
1.2.7 叶绿素荧光参数测定 14:00时收集试验材料,连根带回实验室后,随即参照余蓉培等[22](2019)所述方法使用叶绿素荧光成像系统CFI (英国Techno-logica公司)开展叶绿素荧光参数测定,准备第1个待测样,将待测叶片先平展固定于样品台上,暗适应20 min,打开自动脉冲光(auto pulse),强度为6164 μmol/(m2·s),获取PSII最大光化学效率(Fv/Fm),打开作用光(Actinic light),强度为1000 μmol/(m2·s),与生长环境光强一致,持续5 min进行光适应,然后再次打开自动脉冲光,获取非光化学淬灭(NPQ)、PSII有效光化学量子产量(Fv′/Fm′)以及PSII实际光化学量子效率(ΦPSII)。依次对9种菊科杂草叶绿素荧光参数进行测定。光适应5 min后,设置连续光强梯度2000、1500、1000、600、400、200、100、50、20、0 μmol/(m2·s),每个光强梯度持续30 s,随后打开自动脉冲光,收集ΦPSII,通过公式ETR=光照强度(PPFD)×ΦPSII×0.85×0.5计算各光强梯度下的光合电子传递速率(ETR)。
1.2.8 数据处理 使用软件SPSS 13.0对数据进行最小显著差数法(Least significant difference, LSD)分析,并使用软件Origin 9.1绘图。
2 结果与分析
2.1 9种菊科杂草叶片解剖结构特征
从表1可知,9种菊科杂草叶片厚度为211.93~331.05 μm,角质层和表皮厚度分别为5.50~13.98和25.70~65.22 μm,其中银胶菊(13.98 μm)和三叶鬼针草(10.38 μm)的角质层较厚,苦苣菜的表皮最厚(65.22 μm),有助于减少水分蒸腾。9种菊科杂草中,银胶菊栅栏组织最厚(145.57 μm),而海绵组织最薄(53.19 μm),并且呈现出最大的栅海比(2.74)和最高的紧密度(48.78%),该结构有助于减少水分蒸腾。
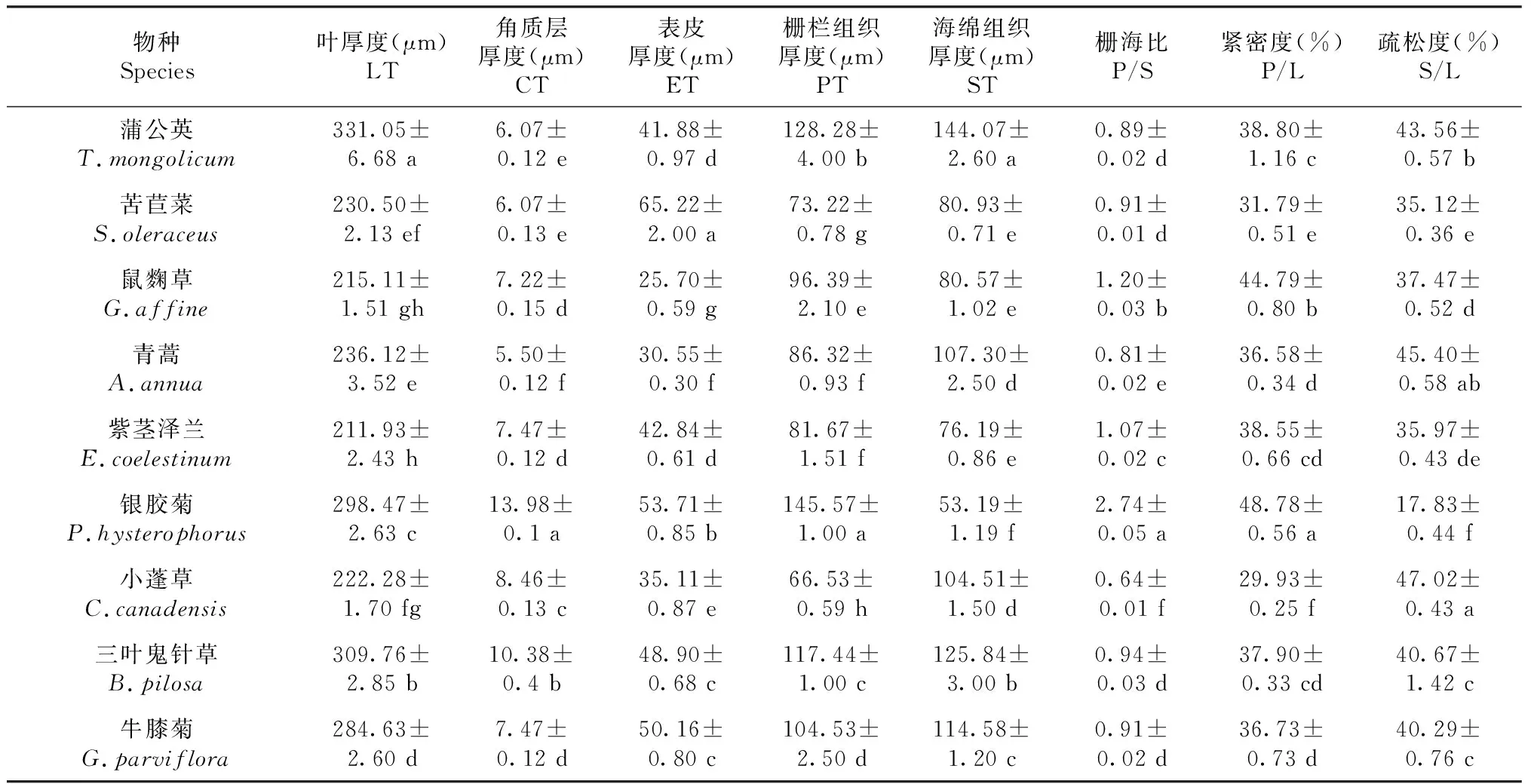
表1 云南9种常见菊科杂草的叶片解剖结构参数Table 1 Leaf anatomical parameters of nine common species of compositae weeds in Yunnan province (mean ± S.E.,n=3)
由图1可知,9种菊科杂草气孔密度为156.18~331.55 个/mm2,苦苣菜(331.55 个/mm2)和牛膝菊(329 个/mm2)的气孔较其它种类密集,有助于其根据环境情况对水分蒸腾进行调节。苦苣菜叶脉密度(10.32 cm/cm2)显著大于其它种类,有助于提升其水分运输效率。
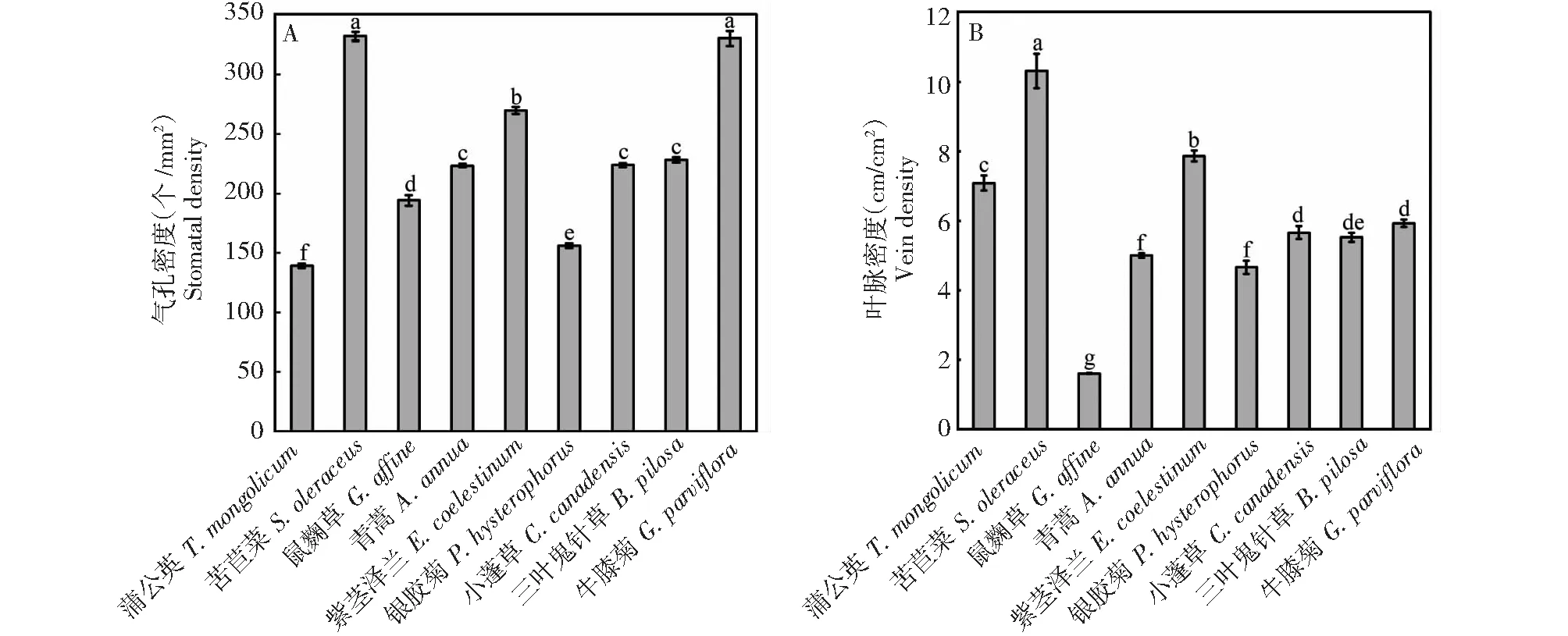
图1 云南9种常见菊科杂草的气孔密度和叶脉密度Fig.1 Stomatal density and vein density of nine common species of compositae weeds in Yunnan province
2.2 9种菊科杂草叶面积、叶干物质含量及比叶面积
从表2可知,9种菊科杂草中,除紫茎泽兰(36.41 cm2)和三叶鬼针草(36.00 cm2)的叶面积较为接近外,其他7种菊科杂草叶面积差异显著。小蓬草的叶干物质含量(0.30 g/g)显著高于其他种类,说明其具有较强的干物质积累能力。9种菊科杂草比叶面积为163.33~420.33 cm2/g,比叶面积最小的是紫茎泽兰(163.33 cm2/g),最大的是蒲公英(420.33 cm2/g),说明不同种类的物质分配和光适应策略存在一定差异。

表2 云南9种常见菊科杂草的叶面积、叶干物质含量以及比叶面积Table 2 Morphological characteristics of nine common species of compositae weeds in Yunnan province(mean±S.E., n=3)
2.3 9种菊科杂草的叶片化学计量特征
从表3可知,9种菊科杂草的C、N、P含量分别为464.84~497.47、16.38~63.52、1.61~1.89 mg/g,C/N、C/P和N/P分别为7.85~29.83、245.80~305.38、9.35~27.27。其中,紫茎泽兰C含量(513.54 mg/g)显著高于其他种类,具有较高的碳同化能力。小蓬草N含量(63.52 mg/g)显著高于其他种类,C/N (7.85)显著低于其他种类,说明其具有较强的N利用率。青蒿P含量最高,为1.89 mg/g,且C/P (245.80)显著低于其他种类,说明其具有较强的P利用率。
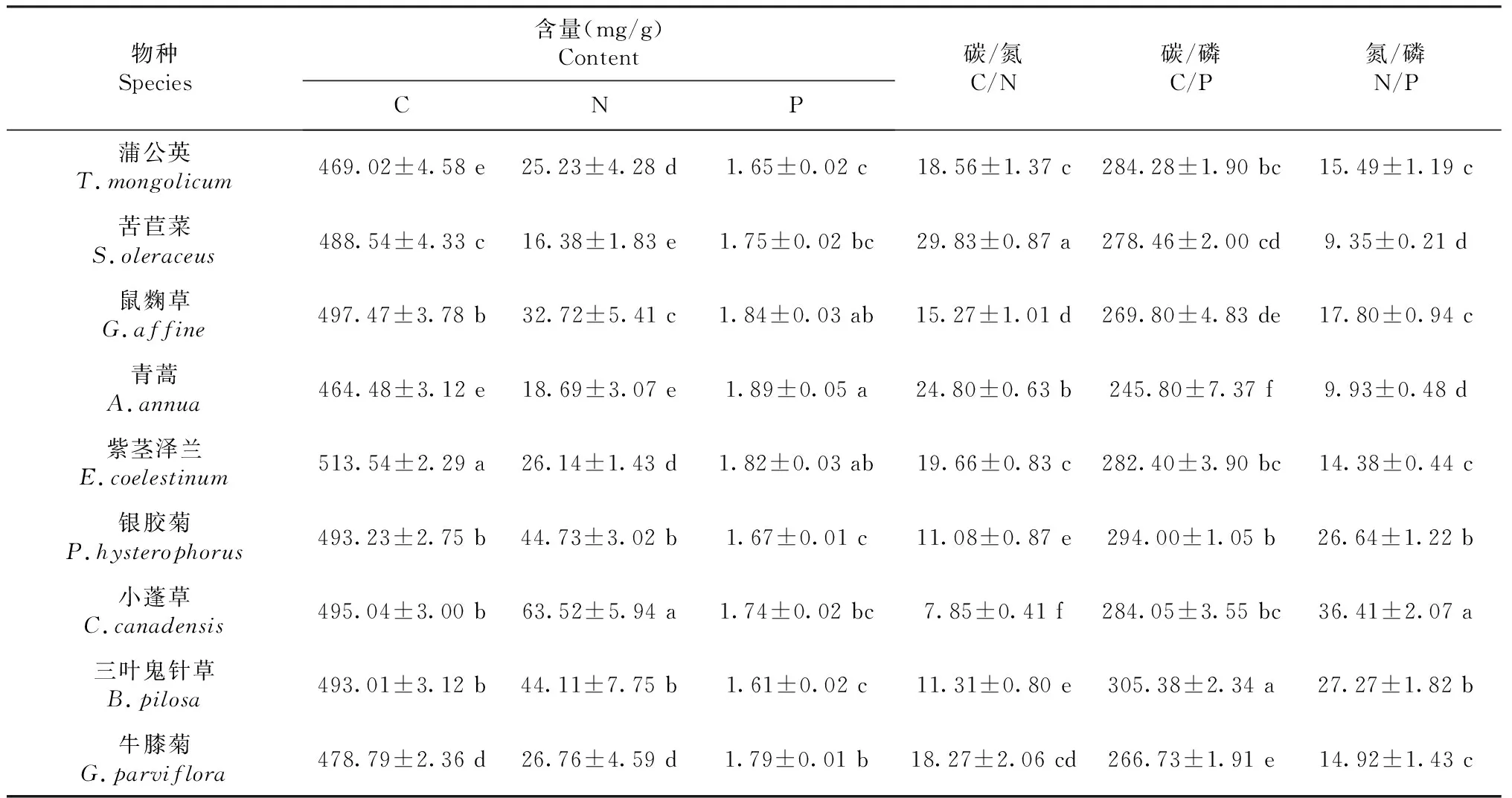
表3 云南9种常见菊科杂草叶片的化学计量特征Table 3 Stoichiometry characteristic of nine common species of compositae weeds in Yunnan province (mean ± S.E., n=3)
2.4 9种菊科杂草的叶绿素含量
从表4可知,9种菊科杂草叶绿素a含量为12.30~19.84 mg/g,叶绿素b含量为3.63~6.66 mg/g,叶绿素总含量为15.92~26.49 mg/g,蒲公英的总叶绿素含量最高,为26.49 mg/g。除鼠麴草和紫茎泽兰的叶绿素a/叶绿素b值较低外,其他种类均大于2.89,说明这些种类对强光环境具有较好的适应性。
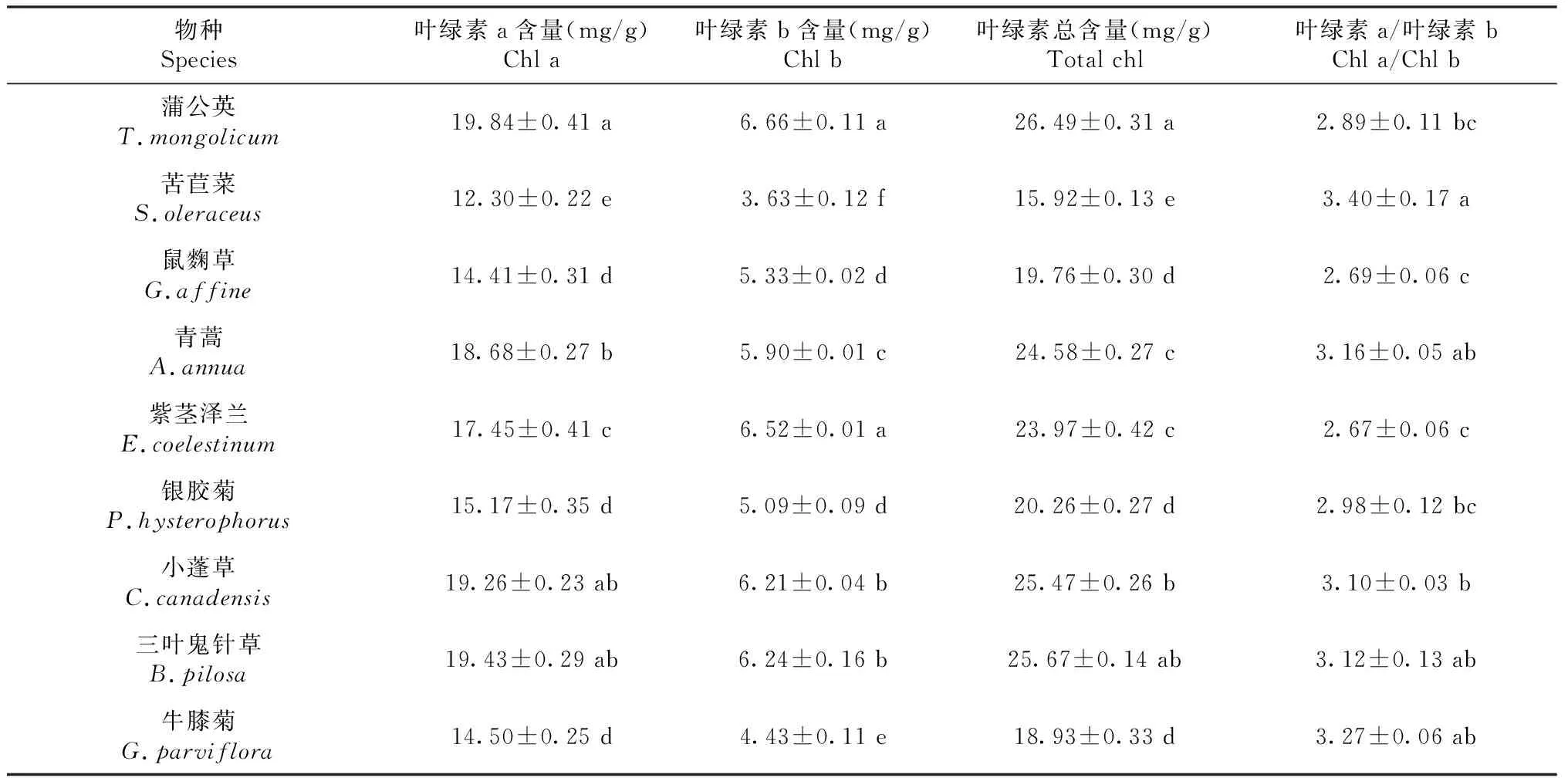
表4 云南9种常见菊科杂草的叶绿素含量Table 4 The chlorophyll content of nine common species of compositae weeds in Yunnan province (mean ± S.E., n=3)
2.5 9种菊科杂草离体叶片叶绿素荧光参数比较
从图2~3可以发现,9种杂草的叶绿素荧光参数及光响应曲线之间存在一定差异,但均表现出较好的光合特性,对环境适应性较强。其中,Fv/Fm均在0.7~0.8(图2-A),具有较好的光合能力,NPQ均在2.0~3.5(图2-B),均具有较好的热耗散能力,有助于减小强光和干旱环境对光合结构的损害。
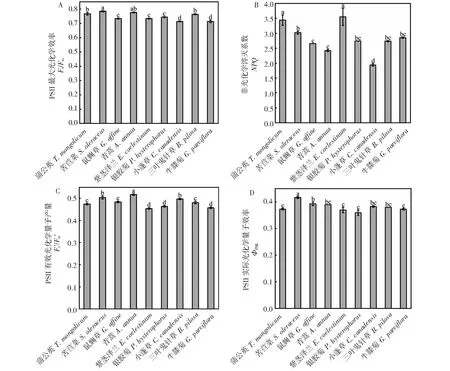
图2 云南9种常见菊科杂草的叶绿素荧光参数Fig.2 The chlorophyll fluorescence parameters of nine common species of compositae weeds in Yunnan province
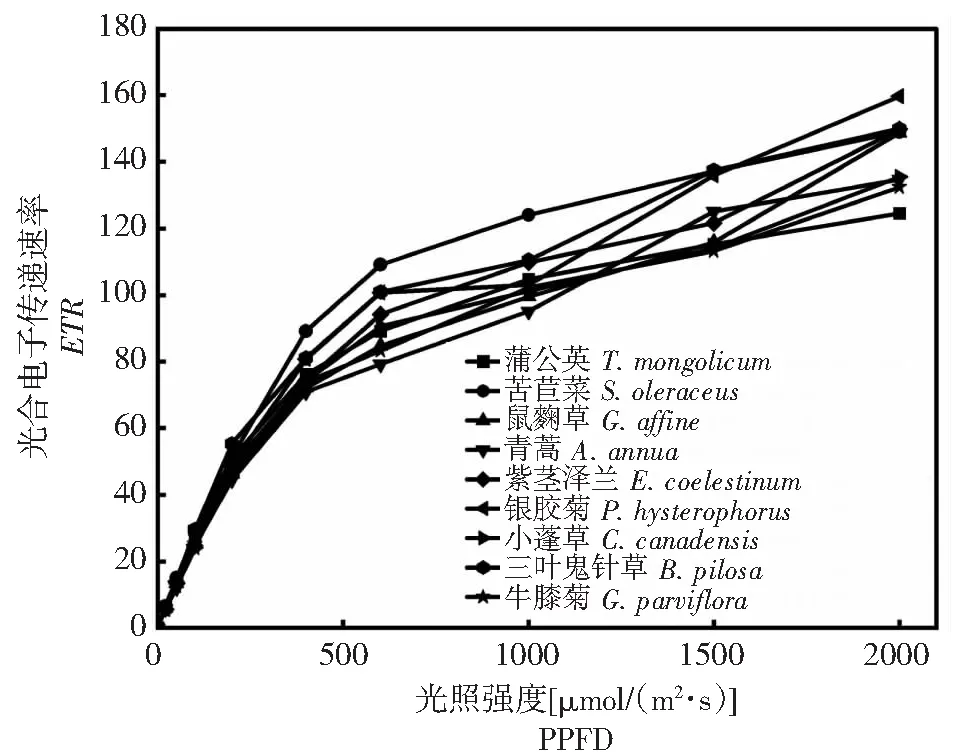
图3 云南9种常见菊科杂草光电子传递速率(ETR)的快速光响应曲线Fig.3 Rapid light-response curve of ETR in nine common species of compositae weeds in Yunnan province
2.6 9种菊科杂草叶片性状综合分析
由表5~6可知,9种菊科杂草中,土著种菊科杂草的叶片性状变异系数在2.51%~61.66%,入侵种菊科杂草的叶片性状变异系数在2.44%~80.89%。在所有叶片性状中,土著种和入侵种的叶面积变异系数均最大,且入侵种的叶面积变异系数(80.89%)显著大于土著种(61.66%),说明不同入侵种菊科杂草的叶面积差异更为显著。

表5 土著种菊科杂草叶片性状综合分析Table 5 Leaf trait analysis of indigenous species of compositae weeds

表6 入侵种菊科杂草叶片性状综合分析Table 6 Leaf trait analysis of invasive species of compositae weeds
9种菊科杂草中,入侵种紫茎泽兰的比叶面积(163.33 cm2/g)最小,但C含量(513.54 mg/g)和NPQ(3.55)最大,有助于其通过较高的碳同化能力和热耗散能力来适应干燥贫瘠的环境。入侵种银胶菊的海绵组织厚度(53.14 μm)最小,角质层厚度(13.98 μm)和栅栏组织厚度(145.57 μm)最大,有利于减少水分散失,进而适应干燥的气候环境。土著种蒲公英具有最厚叶厚度(331.05 μm)、最大比叶面积(420.33 cm2/g)和最高叶绿素含量(26.49 mg/g),有利于提高保水能力及光合速率,进而适应高温干燥的气候环境。土著种苦苣菜具有最厚的表皮组织(65.22 μm),有助于减少水分蒸腾,同时,土著种苦苣菜还具有最大的气孔密度(331.55 个/mm2)和叶脉密度(10.32 cm/cm2),有利于调节蒸腾作用和提高水分运输能力,进而适应干燥的气候环境。
3 讨 论
植物叶片结构变化会影响其生理生态功能进而调节植物对环境的适应性[23]。本研究中9种菊科杂草的叶片厚度和角质层厚度分别为211.93~331.05和5.50~13.98 μm,整体高于青藏高原海拔3600 m处火绒草的叶片厚度(205.51 μm)和角质层厚度(4.7 μm)[9]。气孔密度(156.18~331.55个/mm2)显著大于安徽野生菊属植物叶片气孔密度(51.8~114.1 个/mm2)[24],表明云南9种菊科杂草为了适应本地干燥气候特征,形成了较厚的叶片厚度、角质层厚度以及较高的气孔密度等叶片性状特征,有助于减少水分蒸腾,维持叶片水分供需平衡,同时减少强辐射对叶片的损伤。
碳、氮和磷是植物生长必需元素,C∶N、C∶P表征了植物吸收N、P时同化积累C的能力,反映出植物体的生长速率及营养元素利用效率[25]。本研究中,9种菊科杂草叶C含量为464.84~497.47 mg/g,均高于中国草地生态系统213个物种(438 mg/g)叶C含量[26],N含量为16.38~63.52 mg/g,除苦苣菜和青蒿外,其他7种菊科杂草的N含量均高于中国陆生植物(20.20 mg/g)和全球陆生植物(20.60 mg/g)N含量的平均水平[26-27],P含量为1.61~1.89 mg/g,均高于中国陆生植物(1.46 mg/g)P 含量的平均水平[26],鼠麴草、青蒿以及紫茎泽兰、牛膝菊的P含量与全球陆生植物P含量的平均水平(1.77 mg/g)接近[27]。总体上,9种菊科杂草的N、P含量均高于全国陆生植物的平均水平,在该取样地块表现出较好的生长适应性,这与胡朝臣等[28]对飞机草和紫茎泽兰的研究结果相似。综合分析9种菊科杂草的C、N、P水平,表明云南9种菊科杂草叶片吸收N、P时同化积累C的能力强,能为其光合作用及生理生化过程提供充足的底物和能量,因此能更好的适应外界环境的变化。
根据Koerselman限制性养分理论,当N/P <14时,植物生长主要受N限制,而当N/P>16时,植物生长主要受P限制[11]。本研究中,苦苣菜(9.35)和青蒿(9.93)的N/P <14,N成为这2种杂草在该取样地块生长的限制性因素。银胶菊(26.64)、小蓬草(36.41)以及三叶鬼针草(27.27)的N/P比值远大于16,P成为上述3种杂草在该取样地块生长的主要限制性因素,这与空心莲子草生长的限制性因素相同[11]。
叶绿素含量反应了植物光合作用的强弱[29-30]。研究中云南9种菊科杂草叶绿素含量(15.92~26.49 mg/g)显著大于白花鬼针草、芒萁、鸭跖草的叶绿素含量(1.0~2.5 mg/g )[29],同时也大于叶梅荣等[30]研究中5种草坪杂草的叶绿素含量(1.5~4.5 mg/g),说明9种菊科杂草通过较高的叶绿素积累来提高光合适应能力。同时,9种菊科杂草NPQ为1.94~3.55,具有较强的热耗散能力,说明其在逆境胁迫下对光合结构起一定的保护作用,与水稻在逆境胁迫下通过提高NPQ来降低强光对光系统的损伤机理一致[31]。
叶片性状是植物适应环境体现出的叶片水平上的特征参数[32],但生长在相同气候环境下的不同植物,如:荒漠植物柴柴花(Karelinicacaspica)和骆驼刺(Alhagisparsifolia),两者间的叶片性状亦存在显著差异(P<0.05),展现出对荒漠环境不同的适应性策略[32]。与此相似,云南9种菊科杂草中土著种和入侵种杂草的叶面积、比叶面积、叶干物质含量等叶性状指标变异系数存在显著差异(P<0.05),但在所有指标中,叶面积变异系数均最大,这可能是由于不同物种间叶片形态差异导致[33]。本研究中,尽管土著种杂草苦苣菜和入侵种杂草银胶菊均采自同一样地,但两者对云南的干燥环境采取不同的适应策略。9种菊科杂草中,入侵种银胶菊的海绵组织厚度最小,角质和栅栏组织厚度最大,有利于减少水分散失。土著种苦苣菜具有最厚的表皮组织,有助于减少水分蒸腾,同时,苦苣菜还具有最大的气孔密度和叶脉密度,分别有利于调节蒸腾作用和提高水分运输能力。
4 结 论
9种菊科杂草的叶片解剖结构特征数值、气孔密度、叶脉密度、叶面积、比叶面积等叶片性状存在显著差异(P<0.05),其化学计量特征和叶绿素荧光参数等指标显示9种菊科杂草具有良好的营养特征和光合特性,对云南当地气候环境均具有良好的适应性。叶片性状的差异反应出不同种类菊科杂草对云南干燥环境适应策略的差异,银胶菊具有最厚的角质层及栅栏组织,有助于减少水分散失,而苦苣菜具有最厚的表皮层和最大的气孔密度及叶脉密度,也有助于减少水分散失,提高蒸腾调节能力和水分运输能力,应对干燥环境。
