百合ATP合成酶家族基因ATPase3的克隆和生物信息学分析
2023-06-10贾文杰马璐琳杜文文王祥宁崔光芬
贾文杰,马璐琳,段 青,杜文文,王祥宁,李 想,崔光芬
( 云南省农业科学院花卉研究所/国家观赏园艺工程技术研究中心,昆明 650205)
【研究意义】百合属于百合科(Liliaceae)百合属(Lilium),是多年生草本球根花卉,因其花色艳丽、花型美观、芳香宜人,既能作盆花、切花,又可应用在园林庭院中,是目前国际市场上最为流行的鲜切花种类之一[1]。百合鲜切花主要作为室内观赏用,但其花粉污染给人们造成很大困扰,目前一般都采用人工去雄进行处理,不仅过程繁琐,还浪费大量人力物力,因此雄性不育无花粉百合的培育一直是百合育种的重点方向[2]。【前人研究进展】ATP合成酶(ATPase)是生物体细胞内能量代谢中非常关键的酶,其活性和上游基因表达量对 ATP含量有直接影响,进而影响植物的育性。例如对葱的雄性不育性研究发现,不育系花蕾中ATPase的活性阶段性降低导致不育[3]。对甜菜研究表明,在不育系的营养生长过程中,由于 ATPase活性发生改变,从而导致它产生能量亏缺现象,这是导致甜菜雄性不育系产生的一个主要因素[4]。Dieterich等[5]研究发现,在植物线粒体中可能存在某个基因干扰ATPase的合成,导致植物在花粉形成过程中能量不足,形成雄性不育现象。魏磊等[6]研究发现,在紫稻中存在ATPase基因转录本RNA编辑导致不能翻译成正常的多肽,从而影响紫稻的育性。在玉米、水稻、大豆、小麦等的研究中也发现,ATPase家族基因中的atp6和atp9基因对植物雄性不育的发生也有重要影响[7]。前人研究发现,ATPase活性变化,转录本RNA编辑,ATPase受到干扰,ATPase基因重组或表达量高低等因素都有可能导致植物雄性不育发生。且ATPase家族基因导致雄性不育主要集中在对atp6和atp9基因的研究。【本研究切入点】虽然ATPase导致的雄性不育在许多植物中很早就被广泛发现[8],但目前在百合等观赏园艺植物中还鲜有相关报道。本研究从百合可育系与不育系的转录本中发现1个显著上调的ATPase家族基因,经基因克隆后,进行生物信息学分析,推测ATPase家族基因与百合雄性不育的关系。【拟解决的关键问题】通过转录组数据,克隆百合ATPase基因,并借助相关的生物信息学软件,对该基因序列进行同源比对,并对其所编码的蛋白做进一步分析与功能预测,为研究百合ATPase基因导致的雄性不育以及该基因编码的相应蛋白质功能及性质提供理论基础。
1 材料与方法
1.1 试验材料
本研究以东方百合可育系与不育系为试验材料,该材料种植于云南省农业科学院花卉研究所百合育种基地。根据百合花苞大小判断[9],分别采集花粉母细胞时期可育系与不育系子房、花瓣、叶片、花柱、花丝、花药进行试验。
1.2 试验方法
1.2.1 百合各器官总RNA提取和cDNA反转录 利用改良的CTAB方法[10]提取百合可育系与不育系花粉母细胞时期的子房、花瓣、叶片、花柱、花丝、花药的总RNA,再按照iScriptTMgDNA Clear cDNA Synthesis Kit试剂盒说明书冰浴配制去gDNA体系16 μL,使RNA使用量最大,具体过程为25 ℃孵育5 min,75 ℃孵育5 min,最后去除gDNA。同时冰上配制反转录体系20 μL,具体过程为25 ℃孵育5 min,46 ℃孵育20 min,95 ℃变性1 min,冰上冷却后立即用于qRT-PCR检测。将检测得到的cDNA置于-20 ℃冰箱内保存待用。
1.2.2 百合ATPase3基因实时荧光定量PCR表达分析 分别提取百合可育系与不育系花粉母细胞时期的子房、花瓣、叶片、花柱、花丝、花药的总RNA,反转录成cDNA,进行qRT-PCR分析百合ATPase3基因在百合不同部位中的表达情况。设计引物百合ATPase3-qF、ATPase3-qR,参照bimake的2×SYBR Green qPCR Master Mix试剂盒的操作说明书进行荧光定量PCR分析。配制qRT-PCR反应体系20 μL:SYBR Green qPCR Master Mix (2×)10 μL,上游、下游引物各1 μL,cDNA 1 μL,无菌超纯水7 μL。反应程序为95 ℃预变性2 min,95 ℃变性15 s,60 ℃退火30 s,72 ℃延伸30 s,40个循环。制作溶解曲线:95 ℃15 s,60 ℃60 s,95 ℃15 s。每样品设3个重复。以actin基因为内参,对百合ATPase3基因在百合可育系与不育系花药发育关键时期不同部位的表达进行荧光定量分析。相对表达量按2-ΔΔCt进行计算,方差分析采用软件SPSS中的One-way ANOVA进行,样品间显著性(P<0.05)采用软件Turkey test进行分析,作图使用软件Origin 2018。
1.2.3 百合ATPase3基因全长序列克隆 根据百合可育系与不育系花粉母细胞时期转录组测序结果,挑选表达差异大、数据库功能注释为拟南芥质膜ATPase3基因,设计2对引物(CTG0755-3-YZF1、CTG0755-3-YZR1和CTG0755-3-YZF2、CTG0755-3-YZR1)分别在百合可育系花药中验证百合ATPase3基因。验证后,进行3’和5’ Race扩增。设计引物(CTG0755-3-F1、CTG0755-3-F2)进行3’Race实验,CTG0755-3-F1与试剂盒中的Outer引物进行1stPCR 扩增,CTG0755-3-F2与试剂盒中的Inner引物进行2ndPCR 扩增。设计引物CTG0755-3-F01、CTG0755-3-YZR1,并根据试剂盒说明进行5’Race,在5’和3’末端已知序列设计包括完整ORF 框的特异引物,以百合可育系花粉母细胞时期花药总RNA 的反转录产物为模板,然后进行PCR 扩增。对目的条带进行回收纯化与克隆载体连接后测序。引物序列见表1。

表1 引物序列Table 1 Primer sequence
1.2.4 生物信息分析 利用在线分析软件对百合ATPase3基因进行相关生物信息学分析,软件名称、网址以及用途如表2所示。对所克隆的基因编码蛋白在NCBI数据库中进行BLASTP分析,并与拟南芥ATPase序列进行同源比较分析。

表2 软件的名称及用途Table 2 Software name and usage
2 结果与分析
2.1 百合ATPase3基因荧光定量PCR
如图1~2所示,百合ATPase3基因的扩增和溶解曲线良好。但在百合不同部位的基因表达水平完全不同(图3),在百合可育系中,百合ATPase3基因在花器官中表达量较高,其中在子房中表达量最高,平均相对值达到1.25;而在叶片中表达量极低,平均相对值为0.05。在不育系中,除花药外,其他花器官中百合ATPase3基因表达量也较高,叶片的表达量同样相对较低。在不育系的花药中百合ATPase3基因相对表达量仅为0.02,而在可育系中,该值为0.14,约为不育系中的7倍。
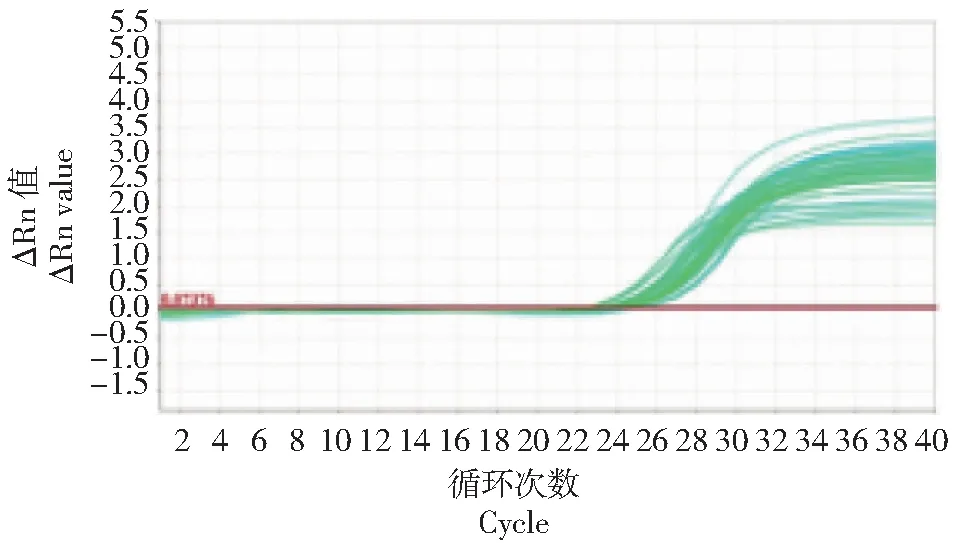
图1 百合ATPase3基因扩增Fig.1 Amplification curve of ATPase3 gene
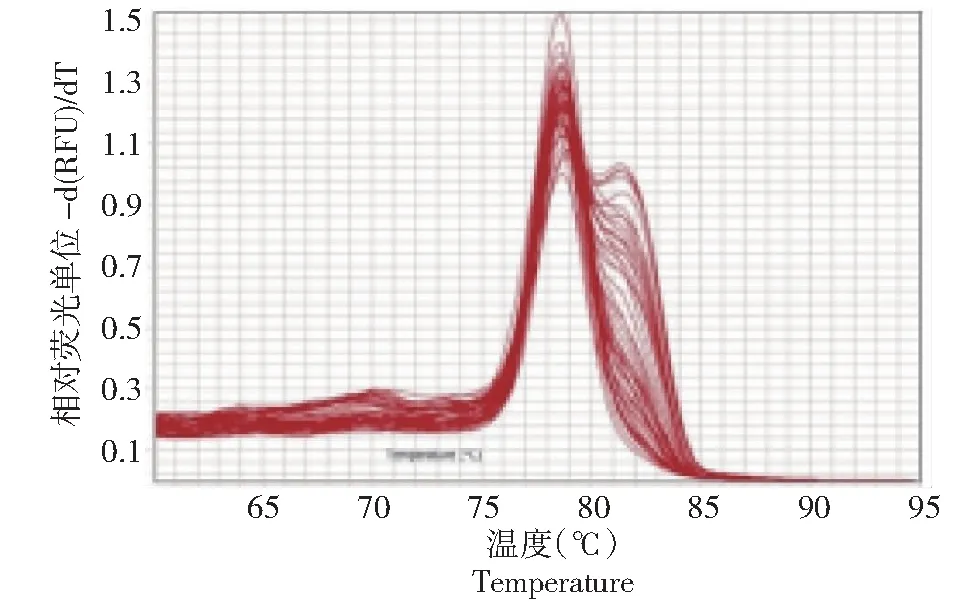
图2 百合ATPase3基因溶解曲线Fig.3 Dissolution curve of ATPase3 gene
2.2 百合ATPase3基因全长克隆
根据ATPase3基因在百合可育系与不育系中不同部位的荧光定量PCR结果可以判定,该基因与百合雄性不育具有相关性。本研究根据转录组获得的序列,设计相关引物验证已知序列,扩增电泳结果如图4所示,回收3号泳道约300 bp条带进行测序,与转录组测序结果一致,已知序列验证成功。同时通过3’和5’Race成功克隆并获得百合ATPase3基因全长序列的目的条带(图5),NCBI Conserved Domain保守结构域网站分析[11-14]显示,百合ATPase3基因编码的蛋白氨基酸区间含有1个典型的保守ATPase3结构域,证实百合ATPase3基因编码的蛋白属于ATPase家族成员蛋白(图6)。

M:DL2000 DNA Marker;1、2:CTG0755-3-YZF1/YZR1 1st 产物、负对照;3、4:CTG0755-3-YZF2/YZR1 2nd 产物、负对照。M:DL2000 DNA Marker;1,2:CTG0755-3-YZF1/YZR1 1st product, negative control;3,4:CTG0755-3-YZF2/YZR1 2nd product, negative control.图4 百合ATPase3基因花药验证Fig.4 Validation of ATPase3 gene in Lilium anthers

1:百合ATPase3基因克隆产物。1:ATPase3 gene cloning results.图5 百合ATPase3基因克隆Fig.5 ATPase3 gene cloning

图6 百合ATPase3基因编码蛋白质的保守域Fig.6 Conserved domain of ATPase3 gene encoded protein
2.3 百合ATPase3基因编码蛋白的理化性质预测
Prot Param预测该蛋白质分子质量约为12.14 KD,理论等电点(pl)为7.71,分子式为C544H871N141O160S6,总原子数为1722;氨基酸序列由19种氨基酸组成,其中异亮氨酸(Ile)含量最为丰富,占总氨基酸含量的13.8%;组氨酸(His)含量和谷氨酰胺(Gln)含量均最低,分别占总氨基酸含量的1.8%;带正电荷残基(Arg+Lys)的总数为13,带负电荷残基(Asp+Glu)的总数为12,其不稳定指数为37.97,属于稳定蛋白。脂肪系数为98.44,总平均亲水性(Grand verage of hydropathicity)为0.138。
2.4 百合ATPase3基因编码蛋白的亲、疏水性预测
利用ExPASy系统中的Prot Scale在线软件分析百合ATPase3基因编码蛋白的亲、疏水性,正值代表疏水性,负值代表亲水性。结果如图7所示,百合ATPase3基因编码的氨基酸中有较多区位点在0刻度线以上,其中最大的位点是73号位点,疏水值为2.544;最小的位点是21号位点,亲水值为-1.878,说明其疏水氨基酸的数目多于亲水性氨基酸,因此推测该蛋白是稳定的疏水性蛋白。
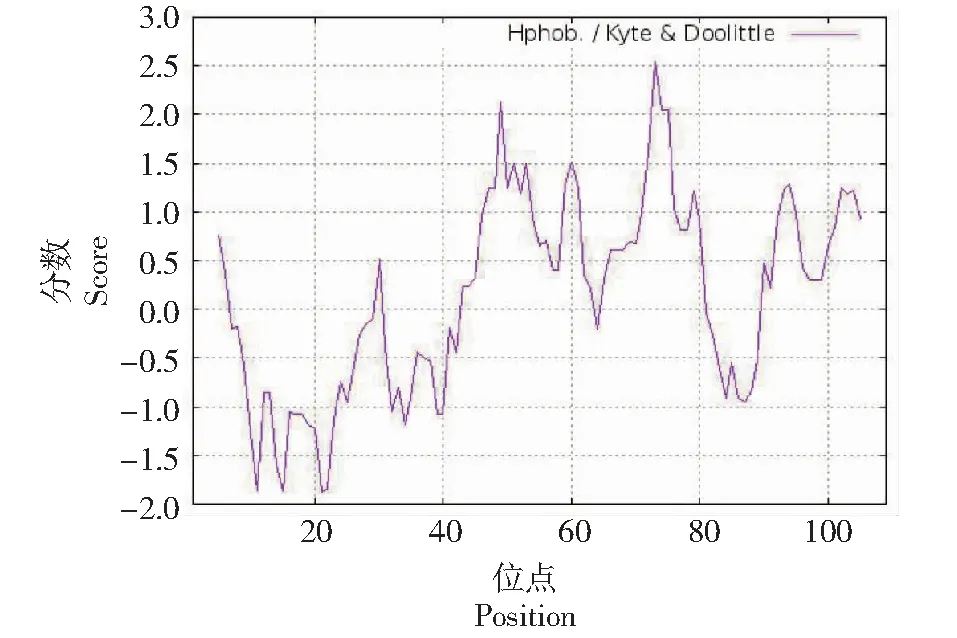
图7 百合ATPase3蛋白的疏水性预测Fig.7 Hydrophobicity prediction of ATPase3 protein
2.5 百合ATPase3基因编码蛋白的跨膜区及亚细胞定位预测
结果(图8)显示,该蛋白存在跨膜螺旋,预期的跨膜螺旋的氨基酸数量为3.36105,该蛋白的前60个氨基酸中预期的跨膜螺旋的氨基酸数量为0.14799,N-端位于细胞质侧膜的总概率为0.19371。从图9可以看出,预测整个蛋白为膜内蛋白和跨膜蛋白的总概率为0.2,为膜外蛋白的概率为0.8,故该蛋白存在跨膜结构。

图8 Predictprotein在线预测百合ATPase3蛋白的跨膜区Fig.8 Prediction of the transmembrane structure of ATPase3 protein using Predictprotein

图9 TMHMM Server v.2.0在线预测百合ATPase3蛋白的跨膜区Fig.9 Prediction of the transmembrane structure of ATPase3 protein using TMHMM Server v.2.0
在线软件WoLF PSORT预测百合ATPase3蛋白的亚细胞定位结果显示,百合ATPase3蛋白定位于亚细胞的分值分别为细胞核(Nuclear)2;细胞质(Cytoplasm)4;叶绿体(Chloroplast)6,质膜(Plasma membrane)1,说明作为ATPase,主要在细胞质与细胞器中发挥作用。
2.6 百合ATPase3基因编码蛋白二级预测
结果表明,该蛋白包含3种二级结构:64个氨基酸形成α-螺旋(Alpha helix),占比58.72%;19个氨基酸形成延伸链(Extended strand),占比17.43%;26个氨基酸形成随机卷曲(Random coil),占比23.85%;该蛋白的主要结构为α-螺旋(图10)。
2.7 百合ATPase3基因编码蛋白三级结构预测
通过把氨基酸序列输入到Swiss-Model程序中,对百合ATPase3基因所编码蛋白的三级结构进行预测(图11),该蛋白主要以α-螺旋为主,3种模式的预测都符合二级结构预测结果。

图11 百合ATPase3蛋白的三级结构预测Fig.11 Predicted tertiary structure of ATPase3 in Lilium
2.8 百合ATPase3同源比较分析
通过百合ATPase3与拟南芥ATPase比较,该蛋白与拟南芥质膜ATPase3同源性最为接近,聚为一类,与拟南芥H[+]ATPase1,H[+]ATPase2,H[+]ATPase3,H[+]ATPase8同源性也较为接近。同时与质膜Ca2+转运ATPase也有一定的同源性,推测属于质膜离子转运蛋白(图12)。

图12 百合ATPase3与拟南芥ATPase同源性比较分析Fig.12 Comparative analysis of homology between Lilium ATPase3 and Arabidopsis ATPase
同时将百合ATPase3蛋白序列在NCBI数据库中进行BLASTP,选取比对结果中一致性最高的其它物种ATPase蛋白序列进行进化树分析(图13)。结果显示,百合ATPase3蛋白与甘蓝(Brassicaoleraceavar.)的亲缘性较近,与洋蓟(Cynaracardunculusvar.scolymus)的亲缘性较远。
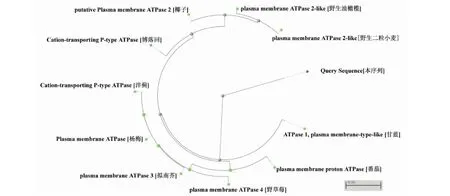
图13 ATPase3蛋白的系统进化树分析Fig.13 Phylogenetic analysis of ATPase3 protein
3 讨 论
3.1 ATPase家族基因与植物雄性不育的关系
作为植物生长发育过程中不可缺少的关键酶之一,ATPase基因的功能和表达差异直接影响到植物能量的供应。在植物生殖生长过程中需要消耗大量能量,因此 ATPase活性,转录本RNA编辑,ATPase受到干扰,ATPase基因重组,ATPase基因表达量高低等因素很可能影响到植物花器官的形成,从而导致雄性不育。目前已经有较多研究表明 ATPase是植物发生雄性不育的原因之一[7]。 在葱和甜菜的研究中表明, ATPase是生物体细胞内能量代谢中非常关键的酶,其活性及上游基因表达量高低对葱和甜菜ATP含量进行直接影响,从而影响到植物的育性[3-4]。魏磊等[6]研究发现,在紫稻中ATPase基因转录本RNA编辑影响紫稻的育性。Singh等[15]认为,ATPase基因上游一个类似tRNA因子因其加工过程不同,造成ATP6基因表达部分差异。Handa等[16]发现,重组发生在不育胞质线粒体DNA在靠近ATP6基因位点的地方,该区域形成了一个新的可编码105个氨基酸的orf 224,在育性恢复过程中,这段基因转录受抑制,这可能导致ATPase基因表达的差异,致使芸苔发生胞质雄性不育。本研究通过百合可育系与不育系花粉母细胞期不同部位的荧光定量PCR发现,ATPase3基因表达的高低影响百合的育性,这与芸苔胞质雄性不育的研究相同,许多学者的研究均发现芸苔胞质雄性不育系和可育系的差异表现在ATPase基因的调控水平上[17]。同时本研究通过保守序列同源分析,发现百合ATPase3与拟南芥质膜ATPase3同源性最为接近,也与拟南芥H[+]ATPase1,H[+]ATPase2,H[+]ATPase3,H[+]ATPase8较为接近。有研究表明质膜 H[+]ATPase对植物的生长发育至关重要,它是植物体内提供能量的重要离子转运蛋白[18]。同时大量研究也表明,H[+]ATPase作为质膜上的关键蛋白,不仅会调节植物的生理功能,在植物适应干旱、重金属、盐、温度、营养缺陷等非生物胁迫时也能发挥巨大作用[19-21]。最新研究还表明,质膜 H[+]ATPase对植物花粉发育和花粉管萌发起着重要作用[22]。在本研究中百合不育系ATPase3基因在花粉母细胞期的花药中显著下调,推测导致花粉质膜离子转运蛋白数量与活性降低,从而影响百合花粉的发育。
3.2 ATPase基因生物信息学预测分析在植物中的应用
目前ATPase基因的生物信息学预测分析已在许多植物中应用,并为实验验证提供方向与理论基础,张云鹤等[23]和熊勇等[24]分别对甘蔗ATPase基因与药用植物黄花蒿ATPase基因进行了电子克隆和生物信息学分析,张静等[25]进行了高粱ATPaseE亚基基因的克隆及其抗逆功能研究,利用电子克隆和实验方法相结合,将为 ATPase的研究提供更丰富的方法及材料。本研究获得百合ATPase3基因,并对其进行生物学信息分析,这些分析说明该预测蛋白具有典型ATPase特征,且预测其为疏水性蛋白,存在跨膜结构,亚细胞定位与质膜离子转运蛋白特性较为一致,推测其在百合中主要作为提供能量的离子转运蛋白发挥作用,它的表达下调,导致百合的雄性不育。
4 结 论
本研究验证ATPase3基因与百合雄性不育相关,并从百合中成功克隆得到ATPase3基因。通过其编码的蛋白生物信息学分析表明,该蛋白含有1个典型的保守ATPase3结构域,属于ATPase家族基因。蛋白相对分子量约为12.14 KD,理论等电点为7.71,为疏水性蛋白,存在跨膜结构。且大多分布在质膜、叶绿体、细胞质中,二级结构预测结果表明该蛋白的α-螺旋占比较高,蛋白稳定,这与三级结构预测结果相符合。对该基因的克隆和生物信息学分析,为百合ATPase3基因编码的相应蛋白质功能及性质的进一步探索提供理论基础,同时也为应用该基因进行百合无花粉分子育种提出了一定的现实依据。
