上调miR-21对长波紫外线诱导光损伤模型人皮肤成纤维细胞的生物学作用
2023-06-07齐立攀赵亚平刘竞闫军飞庞利霞
齐立攀,赵亚平,刘竞,闫军飞,庞利霞
在紫外线辐射,特别是长波紫外线(ultraviolet A,UVA)辐射下,皮肤真皮组织中成纤维细胞易受到紫外线照射发生氧化损伤,引起红斑、皮肤干燥、色素沉着、炎症反应、皮肤松弛、皱纹增加等皮肤性疾病,严重时甚至可引起皮肤癌(包括黑素瘤和非黑素瘤)[1-2]。因此,需要找到一种可针对性防治UVA引起皮肤光损伤的方法。近年,许多学者研究发现皮肤组织中微小RNA-21(microRNA-21,miR-21)与皮肤成纤维细胞的生长、增殖和抗氧化等生理过程有密切关系[3],然而其具体机制不明确。有研究[4]发现,磷酸酶和张力蛋白同源物(PTEN)/磷脂酰肌醇3激酶(PI3K)/蛋白激酶B(AKT)信号通路与氧化损伤密切相关,调控PTEN/PI3K/AKT通路可保护骨髓间充质干细胞免于氧化应激损伤。因此,本研究通过建立UVA照射诱导人皮肤成纤维细胞光损伤模型,采用慢病毒上调miR-21表达,观察miR-21过表达对光损伤模型人皮肤成纤维细胞生物活性、氧化损伤及PTEN/PI3K/AKT信号通路的影响,以期为紫外线辐射的防治提供依据。
1 材料与方法
1.1材料
1.1.1细胞 人皮肤成纤维细胞(批号:CLH50193)购自深圳市豪地华拓生物科技有限公司。
1.1.2试剂与仪器 miR-21模拟物(miR-21 mi-mics)及其阴性对照(miR-21 NC)慢病毒载体(批号分别为2375S和6637S)均购自上海吉凯基因有限公司;PI3K抑制剂LY294002、PI3K激活剂740Y-P(批号分别为IHK-007和ISK-009)均购自北京西美杰科技有限公司;Trizol试剂、细胞计数试剂盒-8(CCK-8)、BCA试剂盒、ECL显色试剂盒(批号分别为4860-056、2903-062、5063-117、2378-021)均购自北京启维益成科技有限公司;DMEM培养基、反转录试剂盒、Annexin V-FITC凋亡检测试剂盒、β-半乳糖苷酶染色试剂盒、超氧化物歧化酶(SOD)、丙二醛(MDA)酶联免疫吸附(ELISA)试剂盒、RIPA裂解缓冲液(批号分别为CK90172、CK2920、CK8316、CK2837、CK3899、CK8249、CK3487)均购自上海碧云天生物技术有限公司;兔源PTEN、PI3K、AKT、磷酸化AKT(p-AKT)、β-肌动蛋白(β-actin)一抗、羊抗兔二抗(批号分别为BC2873、BC4308、BC2908、BC2938、BC4509、BC4305)均购自北京凯瑞力枫科贸有限公司;CO2孵育箱(型号WJ-80A)、UVA发生器(型号SS-08A)、荧光定量PCR(qRT-PCR)仪(型号FQD-98C)均购自合肥市瑞思奇科研仪器有限公司;光学显微镜(型号SMZ25)、酶标仪(型号TECAN spark)均购自北京同德创业科技有限公司;凝胶成像系统(型号SH-520)购自上海智岩科学仪器有限公司。
1.2方法
1.2.1细胞培养、分组及转染 人皮肤成纤维细胞接种于用含10%胎牛血清(FBS)的DMEM培养基的无菌培养瓶中,置于37 ℃ 环境中培养。收集对数生长期细胞以密度1×106个/mL接种200 μL于24孔板,并分为4组:空白对照组、UVA组、UVA+NC组、UVA+miR-21 mimics组、UVA+miR-21 mimics+PI3K抑制剂组、UVA+miR-21 mimics+PI3K激活剂组。除空白对照组外,其余各组细胞均用UVA发生器在距离细胞15 cm处照射,照射剂量为10 J/cm2,时间为12 min[5],照射完毕后更换新的含10%FBS的DMEM培养基;照射1次/d,连续照射7 d。空白对照组细胞不接受UVA照射,正常培养。
取4支EP管,第1支加入miR-21 NC慢病毒载体(24 μL)和完全培养基(180 μL),第2支加入miR-21 mimics慢病毒载体(24 μL)和完全培养基(180 μL),第3支加入miR-21 mimics慢病毒载体(24 μL)和含50 μmol/L PI3K抑制剂LY294002[6]的完全培养基(180 μL),第4支加入miR-21 mimics慢病毒载体(24 μL)和含50 μmol/L PI3K激活剂740Y-P的完全培养基(180 μL),混匀。将EP管中的miR-21 NC慢病毒载体和完全培养基、miR-21 mimics慢病毒载体和完全培养基、miR-21 mimics慢病毒载体和含50 μmol/L PI3K抑制剂LY294002的完全培养基、miR-21 mimics慢病毒载体和含50 μmol/L PI3K激活剂740Y-P的完全培养基,分别加入UVA照射处理后的UVA+NC组、UVA+miR-21 mimics组、UVA+miR-21 mimics+PI3K抑制剂组、UVA+miR-21 mimics+PI3K激活剂组中;空白对照组和UVA组分别加入204 μL完全培养基;培养24 h。各组均设置6次重复。
1.2.2qRT-PCR法检测人皮肤成纤维细胞中miR-21的表达 用Trizol试剂从转染继续培养24 h后的各组细胞中提取总RNA,每组吸取2 μg RNA用反转录试剂盒合成cDNA,将qRT-PCR反应体系置于PCR仪中进行扩增反应,参数为95 ℃预变性11 min;95 ℃ 变性30 s、56 ℃ 退火20 s、72 ℃延伸40 s,39个循环。具体反应环境、条件参照PCR试剂盒说明书进行。以U6为内参,采用2-△△Ct法分析miR-21相对表达量。引物由上海博尔森生物科技有限公司设计合成,所用引物序列见表1。

表1 引物序列
1.2.3CCK-8法检测人皮肤成纤维细胞活力 将转染继续培养24 h后的细胞接种在48孔板上(5×103/孔),每孔中加入14 μL CCK-8溶液,4 h后酶标仪检测590 nm处的吸光度,并计算细胞活力(细胞活力=各干预组吸光度/空白对照组吸光度×100%)。
1.2.4流式细胞仪检测人皮肤成纤维细胞凋亡 收集转染继续培养24 h后的各组细胞,胰蛋白酶消化,用PBS缓冲液将细胞重悬为2×105个/mL的细胞悬液,加入Annexin V-FITC、碘化丙啶(PI)各15 μL 孵育24 min,流式细胞仪检测细胞凋亡率。
1.2.5β-半乳糖苷酶染色法观察人皮肤成纤维细胞衰老情况 取转染继续培养 24 h后的细胞,添加2 mL β-半乳糖苷酶染色液,室温固定30 min。PBS清洗3次,3 min/次。显微镜下观察染色结果,随机选取5个视野范围细胞,高倍镜下进行衰老细胞(染色为深蓝的细胞)计数,计算衰老细胞比例(衰老细胞比例=镜下衰老细胞数/总细胞数×100%),取平均值。
1.2.6ELISA法检测人皮肤成纤维细胞中SOD和MDA含量 用细胞刮收集转染继续培养24 h后的各组细胞,1 300 r/min离心7 min,弃上清,PBS清洗3次,沉淀中加入500 μL双蒸水,在冰浴上匀浆3 min,2 500 r/min离心14 min,取上清液按照SOD和MDA ELISA试剂盒说明书,使用酶标仪测定SOD和MDA含量。
1.2.7蛋白印迹法检测人皮肤成纤维细胞中 PTEN/PI3K/AKT通路相关蛋白表达 收集转染继续培养24 h后的各组细胞,PBS清洗后加入RIPA裂解液,提取蛋白并定量蛋白浓度,取58 μg蛋白进行电泳,蛋白切胶,转膜,加入兔源PTEN(1∶750)、PI3K(1∶500)、p-AKT(1∶510)、AKT(1∶510)、内参β-actin(1∶1 200)一抗孵育,放置4 ℃过夜,TBST清洗,加入羊抗兔二抗(1∶2 300)孵育3 h,加入ECL发光液对蛋白进行显影。Image J软件分析条带灰度值。
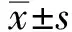
2 结果
2.1各组人皮肤成纤维细胞中miR-21表达水平比较 与空白对照组(1.00±0.00)相比,UVA组(0.69±0.07)、UVA+NC组(0.64±0.06)人皮肤成纤维细胞中miR-21表达水平显著降低(q=4.514、5.329,P<0.05);与UVA组和UVA+NC组相比,UVA+miR-21 mimics组(2.18±0.20)、UVA+miR-21 mimics+PI3K抑制剂组(2.14±0.22)、UVA+miR-21 mimics+PI3K激活剂组(2.21±0.27)人皮肤成纤维细胞miR-21表达水平显著升高(q=21.695、21.113、22.132,22.511、21.928、22.948,P<0.05);UVA+miR-21 mimics组、UVA+miR-21 mimics+PI3K抑制剂组、UVA+miR-21 mimics+PI3K激活剂组人皮肤成纤维细胞中miR-21表达水平差异无统计学意义(q=0.582、0.437、1.019,P>0.05)。
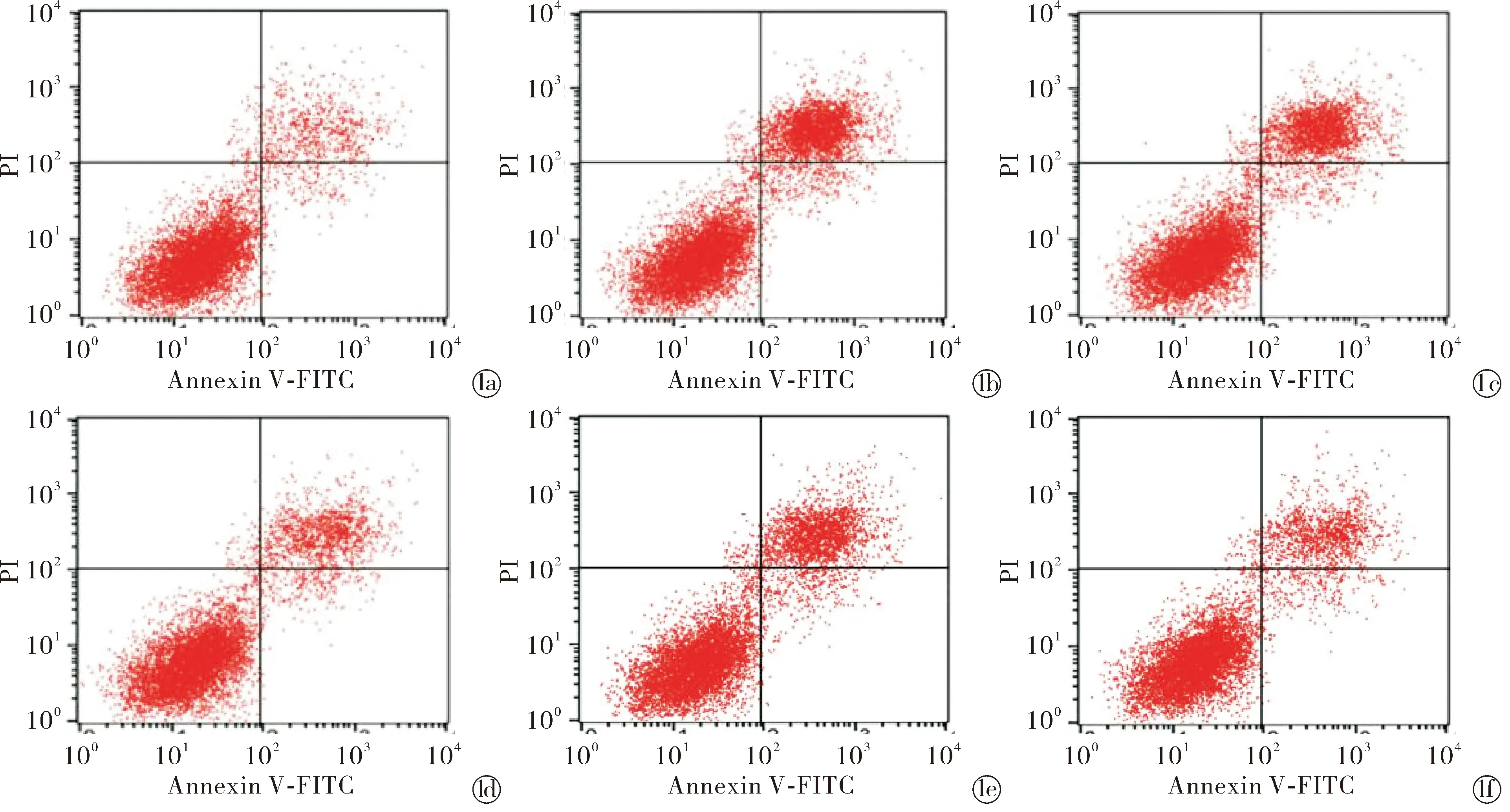
2.2各组人皮肤成纤维细胞活力、凋亡率、衰老细胞比例比较 与空白对照组相比,UVA组、UVA+NC组人皮肤成纤维细胞活力显著降低,凋亡率、衰老细胞比例显著升高(P<0.05);与UVA组和UVA+NC组相比,UVA+miR-21 mimics组、UVA+miR-21 mimics+PI3K激活剂组人皮肤成纤维细胞的活力显著升高,凋亡率、衰老细胞比例显著降低(P<0.05);与UVA+miR-21 mimics组相比,UVA+miR-21 mimics+PI3K抑制剂组人皮肤成纤维细胞活力显著降低,凋亡率、衰老细胞比例显著升高(P<0.05),UVA+miR-21 mimics+PI3K激活剂组人皮肤成纤维细胞活力显著升高,凋亡率、衰老细胞比例显著降低(P<0.05),见表2,图1~2。

表2 各组人皮肤成纤维细胞活力、凋亡率、衰老细胞比例比较
Blank control group; UVA group; UVA+NC group; UVA+miR-21 mimics group; UVA+miR-21 mimics+PI3K inhibitor group; UVA+miR-21 mimics+PI3K activator group
Blank control group; UVA group; UVA+NC group; UVA+miR-21 mimics group; UVA+miR-21 mimics+PI3K inhibitor group; UVA+miR-21 mimics+PI3K activator group
2.3各组人皮肤成纤维细胞中SOD、MDA含量比较 与空白对照组相比,UVA组、UVA+NC组人皮肤成纤维细胞中MDA含量显著升高,SOD含量显著降低(P<0.05);与UVA组和UVA+NC组相比,UVA+miR-21 mimics组人皮肤成纤维细胞中MDA含量显著降低,SOD含量显著升高(P<0.05);与UVA+miR-21 mimics组相比,UVA+miR-21 mimics+PI3K抑制剂组人皮肤成纤维细胞中MDA含量显著升高,SOD含量显著降低(P<0.05),UVA+miR-21 mimics+PI3K激活剂组人皮肤成纤维细胞中MDA含量显著降低,SOD含量显著升高(P<0.05),见表3。

表3 各组人皮肤成纤维细胞中SOD、MDA含量比较
2.4各组人皮肤成纤维细胞PTEN/PI3K/AKT通路相关蛋白表达水平比较 与空白对照组相比,UVA组、UVA+NC组人皮肤成纤维细胞中PTEN蛋白表达水平显著升高,PI3K、p-AKT蛋白表达水平显著降低(P<0.05);与UVA组和UVA+NC组相比,UVA+miR-21 mimics组人皮肤成纤维细胞中PTEN蛋白表达水平显著降低,PI3K、p-AKT蛋白表达水平显著升高(P<0.05);与UVA+miR-21 mimics组相比,UVA+miR-21 mimics+PI3K抑制剂组人皮肤成纤维细胞PTEN蛋白表达水平差异无统计学意义(P>0.05),PI3K、p-AKT蛋白表达水平显著降低(P<0.05),UVA+miR-21 mimics+PI3K激活剂组人皮肤成纤维细胞PTEN蛋白表达水平差异无统计学意义(P>0.05),PI3K、p-AKT蛋白表达水平显著升高(P<0.05);6组人皮肤成纤维细胞AKT蛋白表达水平差异无统计学意义(P>0.05),见图3,表4。

Note:A indicated Blank control group; B indicated UVA group; C indicated UVA+NC group; D indicated UVA+miR-21 mimics group; E indicated UVA+miR-21 mimics+PI3K inhibitor group;F indicated UVA+miR-21 mimics+PI3K activator group.
表4 各组人皮肤成纤维细胞PTEN/PI3K/AKT通路相关蛋白表达水平的比较
3 讨论
太阳光中紫外辐射波长主要包括UVA(320~420 nm)、中波紫外线(UVB,275~320 nm)和短波紫外线(UVC,200~275 nm)[7],长期暴露于UVA可通过诱导活性氧的产生和DNA损伤来激活下游信号和炎性细胞因子的形成,从而引起皮肤损伤和光老化[8]。目前尚无药物针对性预防或治疗UVA导致的皮肤损伤性疾病。近年越来越多的证据表明miRNAs可用于修复皮肤损伤。miRNAs是一类新型内源性、小分子非编码RNA,通过诱导靶mRNAs降解或抑制翻译负调节50%以上人类基因的表达[9]。其中miR-21可参与免疫应答、自身免疫性疾病、创伤愈合过程、纤维化等多种病理生理过程[10-11]。多项研究发现,miR-21能促进成纤维细胞增殖、迁移、胶原合成和增强内皮细胞血管生成活性,利于皮肤损伤区愈合[12]。miR-21可促进皮肤损伤区皮肤再生,在皮肤损伤后上调miR-21表达可促进角质形成细胞迁移,加速损伤区皮肤的再生[13]。抑制miR-21可明显延迟损伤面愈合,损伤面成纤维细胞和胶原减少,因此miR-21是成纤维细胞功能的正调节因子[14-15]。本研究采用UVA照射人皮肤成纤维细胞诱导光损伤模型,结果发现,UVA照射的人皮肤成纤维细胞miR-21表达水平、活力降低,凋亡率、衰老细胞比例升高;经miR-21 mimics慢病毒载体转染的人皮肤成纤维细胞miR-21表达水平、活力升高,凋亡率、衰老细胞比例降低。提示上调miR-21可增强UVA诱导光损伤模型人皮肤成纤维细胞生物活性。
氧化应激是导致许多不良反应和病理条件的原因[16],可导致细胞衰老凋亡。过量UVA照射可使皮肤的前氧化剂-抗氧化平衡向高度氧化状态转变,导致细胞产生过量活性氧,从而抑制抗氧化防御机制引起皮肤成纤维细胞DNAs、蛋白质和其他生物大分子的氧化损伤细胞衰老、皱缩[17]。氧化和抗氧化系统的防御平衡受损,会导致氧化组织损伤,有研究报道[18-19]人皮肤成纤维细胞氧化损伤后会引起MDA含量升高,而SOD含量降低,增强细胞氧化应激状态。本研究发现,UVA诱导光损伤模型人皮肤成纤维细胞中MDA含量升高,SOD含量降低,提示UVA诱导光损伤模型人皮肤成纤维细胞氧化应激水平升高。进一步分析发现,miR-21 mimics慢病毒载体转染的人皮肤成纤维细胞中MDA含量降低,SOD含量升高,提示上调miR-21可减轻UVA诱导光损伤模型人皮肤成纤维细胞氧化应激损伤程度,然而具体作用机制尚不清楚。
PTEN/PI3K/AKT通路是细胞氧化应激过程中的重要通路。研究证实,miR-21可通过PTEN/PI3K/AKT途径增加人端粒酶逆转录酶的表达,抑制细胞衰老凋亡[20-21]。另有研究表明,miR-21可靶向肾纤维化成纤维细胞中PTEN表达,诱导PI3K/AKT途径激活[22]。本研究发现,UVA诱导光损伤模型人皮肤成纤维细胞PTEN蛋白表达升高,PI3K、p-AKT蛋白表达降低;经miR-21 mimics慢病毒载体转染后,细胞中PTEN蛋白表达降低,PI3K、p-AKT蛋白表达升高;提示miR-21过表达可能通过下调PTEN,激活PI3K/AKT信号通路,减轻UVA诱导的人皮肤成纤维细胞损伤。为了进一步验证miR-21过表达对PI3K/AKT信号通路的调控作用,本研究在miR-21 mimics慢病毒载体转染UVA诱导光损伤模型人皮肤成纤维细胞的基础上,采用PI3K抑制剂LY294002和PI3K激活剂740Y-P分别进行干预,结果显示,PI3K抑制剂LY294002可逆转miR-21过表达对UVA诱导的人皮肤成纤维细胞损伤的保护作用,而PI3K激活剂740Y-P则可增强miR-21过表达对UVA诱导的人皮肤成纤维细胞损伤的保护效果。
综上所述,上调miR-21可增强UVA诱导光损伤模型人皮肤成纤维细胞活力,抑制其凋亡、衰老和氧化损伤,可能与调控PTEN/PI3K/AKT信号通路有关。但本研究还存在一定的局限性,miR-21对人皮肤成纤维细胞生物活性、氧化损伤的影响可能涉及其他通路,有待深入研究。
