金沙江上游2 种近缘龙蜥物种逃逸行为比较研究
2023-06-01李凌向以华刘帆文宇浩郭鹏董丙君吴亚勇
李凌,向以华,刘帆,文宇浩,郭鹏,董丙君,吴亚勇*
(1.沈阳师范大学生命科学学院,沈阳 110034;2.宜宾学院农林与食品工程学部,四川 宜宾 644000)
逃逸行为是动物对周围环境现存和潜在捕食风险的一种行为反应(Dimond & Lazarus,1974)。通过逃逸可有效规避被捕食风险,增加自身和后代的适合度(Treves,2000;Bednekoff,2001;Beauchamp,2014)。但逃逸是一种高耗能活动,包括奔跑(或飞行)过程本身的耗能及觅食时间的减少导致个体能量获取的减少(Dowling & Bonier,2018),因此,动物需根据捕食风险,对逃逸行为策略进行权衡,只有当捕食压力增大时,才可能选择逃逸(Childress & Lung,2003;车烨,李忠秋,2014)。为降低逃逸成本,动物也可能通过信号展示的方式警示捕食者,如一些蜥蜴物种面对捕食者时的俯卧撑或抬腿等动作,可达到降低捕食风险的目的(Leal,1999;Fontet al.,2012)。动物一旦采取逃逸策略,通常也会权衡捕食风险和逃逸成本,从而优化逃逸距离,以便在躲避被捕食风险和保持能量之间做出权衡(Ydenberg & Dill,1986;Cooper &Frederick,2007;张凡梦等,2021)。当前,国内外关于动物逃逸行为的研究多集中于鸟类和兽类,而对其他类群的关注较少,特别是移动能力较弱的类群(如蛇、蜥蜴等)。因此,开展动物逃逸行为研究可有效揭示动物反捕食策略,并评估人类活动对野生动物的影响程度,这对濒危物种的保护具有重要意义(车烨,李忠秋,2014;汪开宝等,2018)。
动物的逃逸行为受多种因素的影响,包括性别(Vanhooydoncket al.,2007)、种群大小(Liet al.,2009)、年龄(Liet al.,2013)、捕食风险(Xuet al.,2013)和季节(Muposhiet al.,2016)等。雌雄两性由于生理周期、身体条件和繁殖角色的差异,逃逸行为常主要表现为3 种模式:(1)雄性偏好采取高风险行为,即雄性逃逸起始距离和逃逸距离显著短于雌性,如北美驯鹿Rangifer tarandus(Stankowich,2008)、雄性赭红尾鸲Phoenicurus ochruros(Kalbet al.,2019)和麻雀Passer montanus(García-Arroyo & MacGregor-Fors,2020)等,可能是雄性通常比雌性胆量更大,身体更强壮,逃逸能力(速度、耐力等)更强,成功逃逸概率更大,这种趋势在交配季更为明显;(2)雌性偏好采取高风险行为,即雌性逃逸起始距离和逃逸距离显著短于雄性,如澳洲南部石龙子Eulamprus tympanum(Schwarzkopf& Shine,1992)和扁石蜥Platysaurus intermedius wilhelmi(Lailvauxet al.,2003)等,可能是雌性在繁殖活动(如孵化、育幼等)中投入更多,逃逸代价更大,这种趋势在怀孕个体中更为显著;(3)雌雄逃逸风险程度差异不显著,即雌雄逃逸起始距离和逃逸距离无显著差异,如棱鳞隐耳鬣蜥Holbrookia propinqua(Cooper,2003)、普通壁蜥Podarcis muralis(Fontet al.,2012)等,可能是雌性在自然状态下体色普遍更暗淡,环境隐秘性更强,被捕食者发现的概率更小,因此允许捕食者接近(Vanhooydoncket al.,2007)。此外,由于遗传、表型的差异性,同一类群近缘物种间的逃逸行为也可能表现出一定的差异性,如白腰雪雀Montifringilla taczanowskii的惊飞距离显著大于棕颈雪雀M.ruficollis,这与2 个物种的体型差异有关(张贺等,2016)。Møller 和Erritzøe(2010)发现,体型越大的物种常具有更大的惊飞距离;但Smith和Lemos-Espinal(2005)研究了生存于墨西哥的4 种角蜥Phrynosomatid 的逃逸行为,发现2 种近缘刺蜥(Sceloporus gadoviae和S.anahuacus)的逃逸行为具有较大的相似性。整体来看,关于雌雄两性之间和近缘物种之间的逃逸行为的比较研究还需深入。
分布于我国横断山区的巴塘龙蜥Diploderma batangense和丽喉龙蜥D.formosgulae为探究近缘物种逃逸行为的种内和种间差异提供了很好的模型。这2种龙蜥为近年新描述物种,表型及遗传差异较小,为典型的近缘物种(李操等,2001;Wanget al.,2021)。野外观察发现,2 个物种频繁使用动作信号(点头、俯卧撑和摆尾)进行求偶、领地守卫等社会交流活动,但是否具有警示捕食者的功能还不明确。前期调查显示,2 种龙蜥均栖息于金沙江上游的干热河谷灌丛生境,分布区均极为狭小且紧密相连,属典型的狭域性物种和邻域分布物种(图1)。金沙江上游水电站、公路设施的大量修建已严重威胁到这2个物种赖以生存的栖息地,其生存状态受到极大挑战,种群数量呈下降趋势,在《国家重点保护野生动物名录》(国家林业和草原局,农业农村部,2021)中,巴塘龙蜥已被列为国家二级重点保护野生动物。因此,开展横断山区龙蜥物种逃逸行为研究,可有效评估其抗干扰能力,为龙蜥的保护研究提供一定的依据。本研究拟解决以下科学问题:(1)2 种龙蜥的逃逸行为模式是怎样的?(2)2 种龙蜥的逃逸行为是否存在种内和种间差异?(3)2 种龙蜥社会交流过程中频繁使用的动作信号,是否具有警示捕食者的功能?
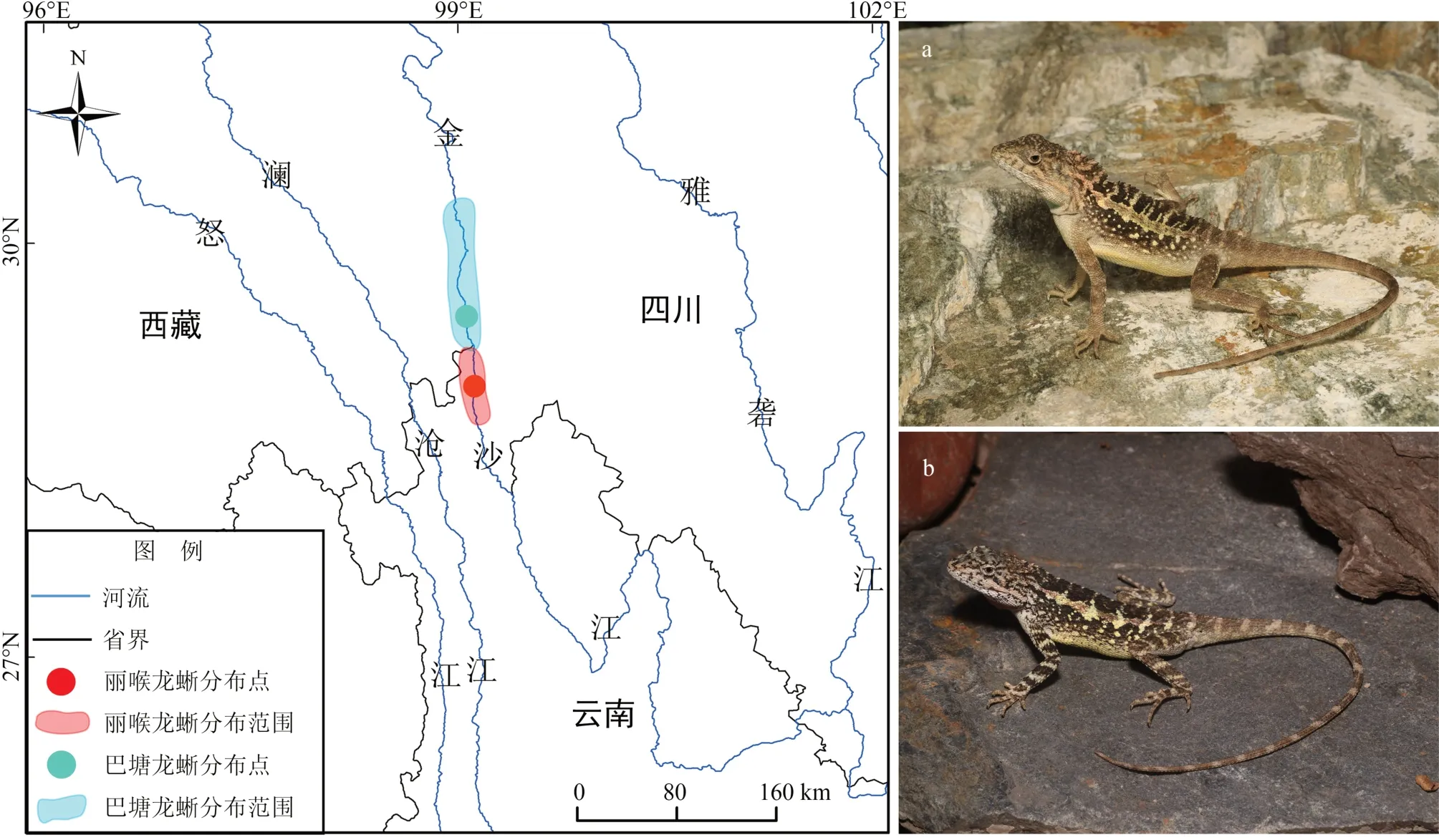
图1 巴塘龙蜥和丽喉龙蜥的分布区域、研究位点及形态Fig.1 Distribution,study site and morphology of Diploderma batangense and D.formosgulae
1 研究方法
1.1 研究区域
研究位点位于金沙江上游(图1),其中,巴塘龙蜥分布于四川省甘孜藏族自治州巴塘县苏哇龙乡(99.059 116°E,29.461 718°N,海拔2 412 m),丽喉龙蜥分布于云南省迪庆藏族自治州德钦县羊拉乡(99.119 812°E,28.954 233°N,海拔2 552 m)。研究位点均为2 种龙蜥的典型分布区,生境破坏较小,种群密度较大;属典型干热河谷稀疏灌丛草丛生境,裸岩较多;气候炎热少雨,年均温20~23 ℃,年均降水量600~800 mm;优势植物为岷谷木蓝Indigofera lenticellata、两头毛Incarvillea arguta、戟叶酸模Rumex hastatusy和白刺花Sophora davidii等。
1.2 逃逸行为量化
以人模拟天敌匀速靠近目标蜥蜴,测量其在面临捕食风险时的行为反应,量化2种龙蜥的逃逸行为(Qiet al.,2014;Donihueet al.,2021)。实验由2 名测试者共同完成:当发现蜥蜴个体时,一人迅速架设摄像机(SONY HDR-PJ670;上海)聚焦目标蜥蜴,持续追踪记录其后续行为;另一人在距目标蜥蜴约15 m 处以0.5 m·s-1匀速缓慢靠近,当目标蜥蜴开始逃逸(至少移动0.1 m)时停止前进,并迅速用标记物1(白色塑料瓶盖)记录测试者当前位置(位置1);待逃逸至避难所后,用标记物2(红色塑料瓶盖)和标记物3(黑色塑料瓶盖)分别标记逃逸初始位置(位置2)和逃逸停止位置(位置3)。用卷尺测量逃逸起始距离和逃逸距离(精度0.01 m)。逃逸起始距离定义为蜥蜴开始逃跑时,捕食者(测试者)距离目标蜥蜴逃逸初始位置之间的直线距离,即位置1 和位置2 的直线距离;逃逸距离定义为蜥蜴逃逸初始位置与逃逸停止位置之间的距离,即位置2和位置3的直线距离(图2)。
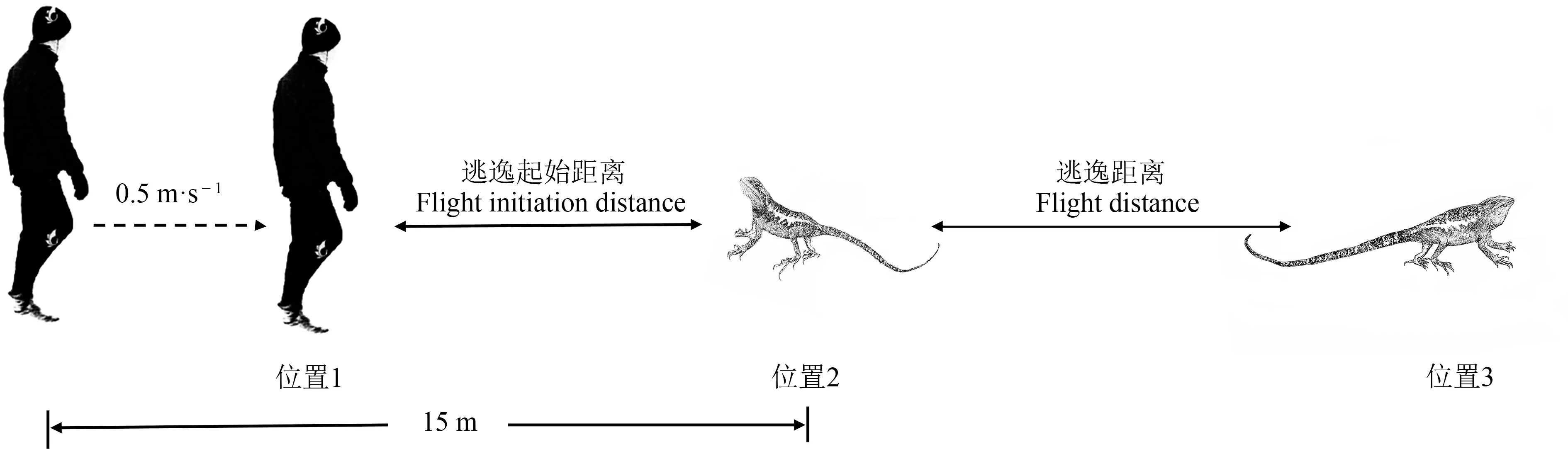
图2 龙蜥逃逸行为测试示意图Fig.2 Schematic diagram of measuring the escape behavior of mountain dragon lizards
一般而言,逃逸起始距离和逃逸距离越短,表明个体采取风险行为的偏好性更强。为降低“捕食者”个体差异导致的蜥蜴行为偏差,所有“捕食者”均由同一测试者完成,且身穿黑色服装。为避免蜥蜴重复测量,各实验个体之间距离至少相距100 m。测试完成后,采用钓竿制作的套索工具捕获目标蜥蜴,测量相关形态数据。形态指标测量完成后均放归至原捕捉位点。野外工作时间为2020 年8 月 下 旬 至9 月上旬每天09∶30—12∶00,即龙蜥活动的高峰期采集数据。所有数据采集对象均为成体。
野外工作完成后通过视频回放查看目标蜥蜴在面临捕食压力时的行为,量化目标蜥蜴的行为类型(信号展示、静止观察捕食者、逃逸等)和顺序,并记录最终是否使用避难所(洞穴或灌丛)。
1.3 形态测量
依据体色(雄性背部颜色鲜艳,有较宽的黄色颜色条纹;雌性背部颜色偏暗淡,无黄色条纹或条纹较窄),以及半阴茎有无判断个体性别。使用游标卡尺和电子秤分别测量头体长(精度0.01 mm)和体重(精度0.01 g)。
1.4 数据分析
由于逃逸起始距离和逃逸距离均不满足正态分布和方差齐性检验,因此,在进行种内(雌雄两性)和种间比较时均采用非参数检验(Mann-WhitneyUtest)。采用一元线性回归方法分别检验逃逸起始距离、逃逸距离和使用避难所比例与头体长和体重之间的相关关系;采用卡方检验分析避难所使用比例的种内和种间的差异。统计值均以xˉ±SD 表示,显著性水平设为0.05。所有数据统计分析均在SPSS 26.0进行。
2 研究结果
共获取95只龙蜥成体的逃逸行为数据,其中,巴塘龙蜥46 只(雄性25 只,雌性21 只),丽喉龙蜥49只(雄性27只,雌性22只)。
2.1 逃逸行为的模式
2 种龙蜥雌雄个体在面临捕食压力时,均展现出探视、调整姿势和逃逸3 种行为:当捕食者(人)靠近时,立即停止先前的行为(捕食、信号展示或晒背等),并通过探视的方式(转头注视捕食者)观察捕食者的后续行为;若捕食者继续接近,则静止并调整姿势(如四肢站立,身体调整至朝逃跑方向,并转头注视捕食者),继续观望捕食者;若捕食者进一步靠近,捕食压力增大,则迅速逃离至其他地点,继续观望或进入灌丛(或洞穴)(图3)。
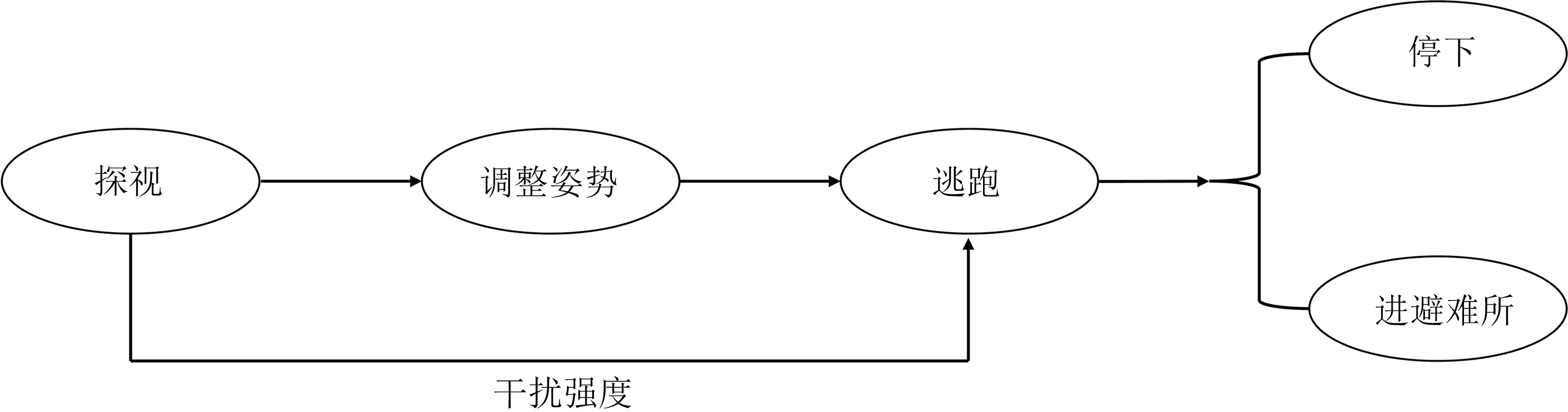
图3 巴塘龙蜥和丽喉龙蜥逃逸行为模式Fig.3 Escape behavior patterns of Diploderma batangense and D.formosgulae
2.2 逃逸行为的比较
丽喉龙蜥雄性个体的逃逸起始距离为222.93 cm±112.22 cm(n=27),雌性为197.64 cm±132.82 cm(n=22),两 性 间 的 差 异 不 显 著(Z=-0.784,P=0.433),逃逸起始距离与个体头体长和体重无相关性;巴塘龙蜥雄性的逃逸起始距离为216.36 cm±79.66 cm(n=25),雌性为206.62 cm±96.66 cm(n=21),两性间的差异也不显著(Z=-0.067,P=0.544),逃逸起始距离与个体头体长和体重无关(表1)。此外,2 种龙蜥的逃逸起始距离之间的差异均不显著(雄性:Z=0.165,P=0.869;雌性:Z=-1.021,P=0.307;图4)。
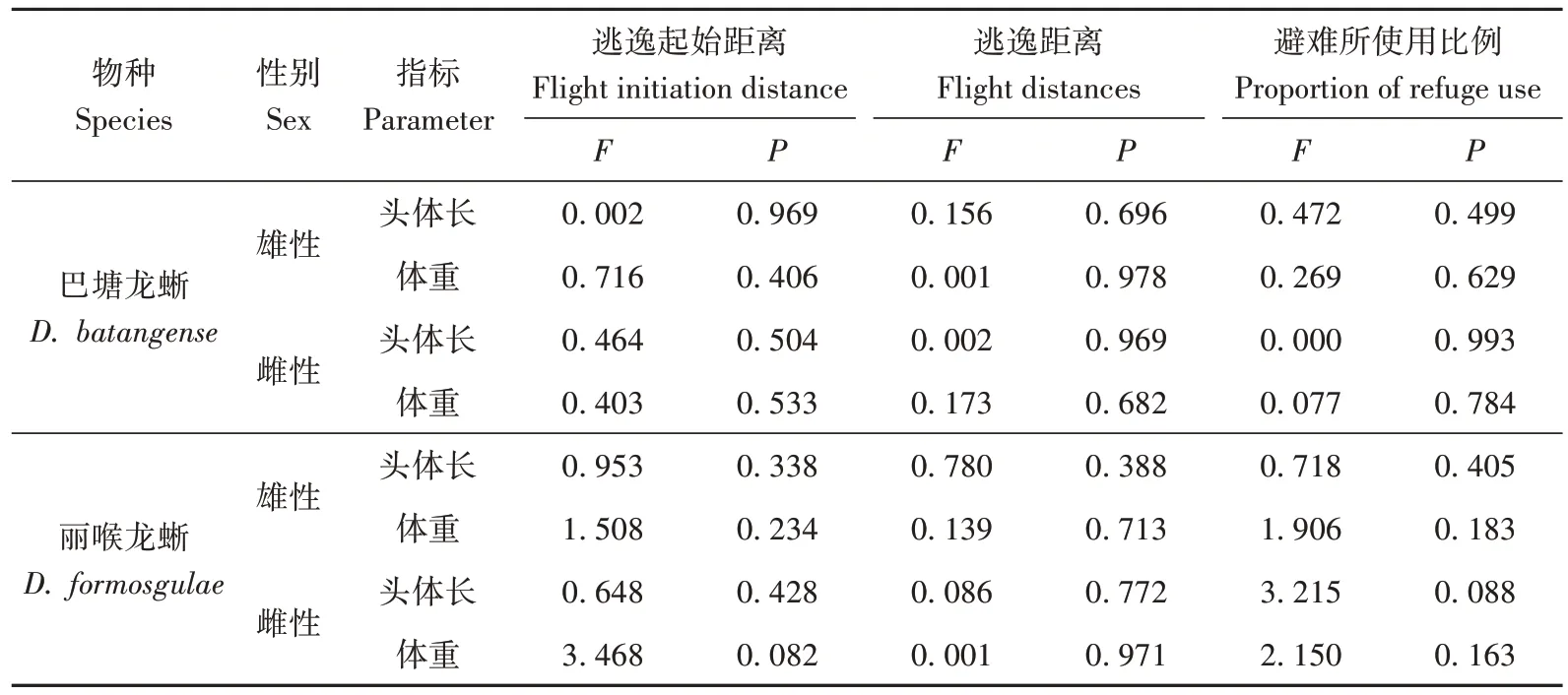
表1 巴塘龙蜥和丽喉龙蜥雌雄两性逃逸行为与体型大小的关系Table 1 Relationship of escape behavior with individual body size for both sexes of Diploderma batangense and D.formosgulae
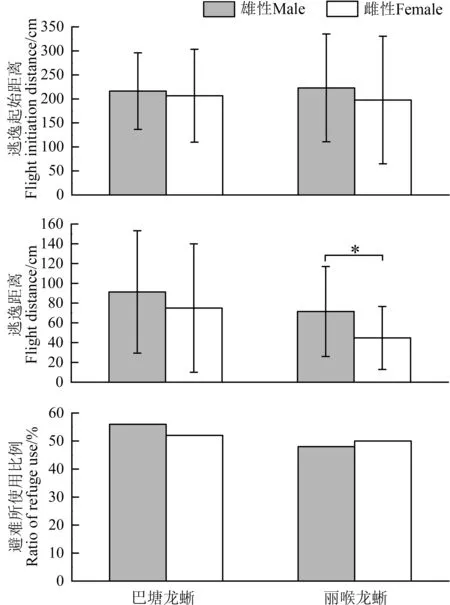
图4 巴塘龙蜥和丽喉龙蜥逃逸行为的种内(两性)和种间(同性别)比较Fig.4 Intraspecific (both sexes)and interspecific(same sex)comparisons of escape behavior of Diploderma batangense and D.formosgulae* P<0.05
雄性丽喉龙蜥的逃逸距离为71.48 cm±45.50 cm(n=27),显著大于雌性44.73 cm±31.86 cm(n=22)(Z=-2.785,P=0.005),但两性的逃逸距离与个体头体长和体重均无关;雄性巴塘龙蜥的逃逸距离为91.24 cm±61.90 cm(n=25),雌性为74.95 cm±64.92 cm(n=21),两性间差异不显著(Z=-1.346,P=0.178);两性的逃逸距离也与个体头体长和体重无关(表1)。2 种龙蜥雌雄个体的逃逸距离也均未表现出差异性(雄性:Z=-1.035,P=0.301;雌性:Z=-1.908,P=0.056;图4)。
当捕食风险增大时,2 种龙蜥所有个体均会逃逸至新地点。丽喉龙蜥48%(13/27)的雄性个体、50%(11/22)的雌性个体,巴塘龙蜥56%(14/25)的雄性个体、52%(11/21)的雌性个体会直接进入避难所(洞穴或灌丛中),而其余个体则会静止在新地点,继续观望。2 种龙蜥雌雄两性使用避难所的倾向与个体头体长和体重无关(表1)。2种龙蜥使用避难所的比例未表现出显著差异性(丽喉龙蜥:χ2=0.001,P=0.979;巴塘龙蜥:χ2=0.017,P=0.897);2 个物种雌雄两性使用避难所的比例亦未表现出显著差异性(雌性:χ2=0.024,P=0.876;雄性:χ2=0.000,P=0.991;图4)。
2.3 动作信号与反捕食
在面临捕食压力时,2 种龙蜥雌雄个体均未展示出点头、俯卧撑和摆尾等动作,表明在社会交流过程中使用的动作信号不具有警示捕食者的功能。
3 讨论
3.1 龙蜥逃逸行为模式
面临危险时,动物成功逃逸可有效降低被捕食风险,显著提升自身适合度(Beauchamp,2001)。逃逸活动主要包括探查、环视、探听和逃逸等过程(车烨,李忠秋,2014)。由于逃逸是一个耗能的过程,只有当被捕食压力足够大时,动物才会选择逃逸,以保证自身安全。本研究发现金沙江上游2种近缘龙蜥物种的逃逸行为与其他动物的逃逸行为模式相似性较高:当发现捕食者(人)时,龙蜥并不会选择直接逃逸,而是采取观望的方式关注捕食者的后续行为,并调整自身姿态;当捕食压力逐步增大时,才会选择逃逸至其他安全地点。这种逃逸行为模式是长期进化的结果,具有重要的适应性意义:一方面可有效规避捕食者的捕杀风险,提高自身和种群的存活率;另一方面,也可有效节省能量,减少非必要的逃逸耗能(Dial,1986;Cooper,2001)。
3.2 龙蜥逃逸行为的种内和种间差异
雌雄两性由于生理、表型和繁殖角色的差异,逃逸行为常表现出差异性。就蜥蜴类群而言,雄性个体因具有更大的体型和更高的雄性激素水平,胆量普遍更大、逃逸能力更强,面临捕食危险时,通常会采取更具有风险性的行为,即更短的逃逸起始距离和逃逸距离(Lailvauxet al.,2003;Nobleet al.,2014)。但本研究未检测到巴塘龙蜥雌雄两性逃逸行为的差异;雄性丽喉龙蜥具有更长的逃逸距离,但逃逸起始距离和使用避难所比例也未表现出两性差异。2 种龙蜥雌雄两性逃逸行为的差异不显著,一方面可能是雌雄两性经历了相似的捕食者驱动的选择压力,雄性个体虽然均具有更大的体型,但并未检测到雌雄两性的逃逸距离、逃逸起始距离和避难所使用比例与个体体型大小的相关性,表明体型并不是影响两性逃逸行为的因素,这与早期青海沙蜥Phrynocephalus vlangalii的研究结果较为一致(Qiet al.,2014)。另一方面,也可能与雌雄两性的繁殖状态有关,8 月下旬到9月上旬为雌性产卵季,雌性个体具有较高的雌性激素水平,面对捕食者时,胆量更大,可能会采取风险行为,以降低因逃逸带来的能量损耗,因此,具有较短的逃逸起始距离和逃逸距离。该推测已在部分蜥蜴物种中得到证实,如怀孕的雌性澳洲南部石龙子面对捕食者时比雄性和未怀孕的雌性个体胆量更大,逃逸起始距离和逃逸距离更短(Schwarzkopf & Shine,1992);Cooper(2009)研究条纹高原蜥Sceloporus virgatus的逃逸行为也发现,在面对捕食压力时,有卵泡或怀孕的个体会显著缩短其逃逸起始距离。本研究未测量龙蜥交配季的逃逸行为,无法比较不同繁殖状态龙蜥个体逃逸行为的差异,后续将拓展补充相关研究。
近缘物种因选择压力的差异,往往会在形态、行为、生境选择等方面表现出一定的差异性(Johnson & Wade,2010;Gabelaiaet al.,2018)。青藏高原同域分布的白腰雪雀、棕背雪雀M.blanfordi和棕颈雪雀的体型大小和逃逸行为均存在一定的差异性(Geet al.,2011);分布于智利的花环树鬣蜥Liolaemus lemniscatus和棕树鬣蜥L.fuscus虽头体长未表现出差异性,但逃逸行为策略却表现出显著的差异性(Jaksić & Núñez,1979)。近缘物种的逃逸行为差异往往与体型大小有关,特别是鸟类(McLeodet al.,2013,张贺等,2016,杨爱芳等,2021),但在一些蜥蜴类群中未得到支持(Bulova,1994;Smith & Lemos-Espinal,2005)。本研究选择了体型大小有一定差异的巴塘龙蜥和丽喉龙蜥作为研究对象,从种间层面比较了2个近缘物种的逃逸行为,但未检测到逃逸行为的种间差异,这与Labra 和Leonard(1999)对3 种近缘树鬣蜥的逃逸行为比较研究的结果一致。此外,物种的反捕食策略往往是长期适应性进化的结果,巴塘龙蜥和丽喉龙蜥均栖息于金沙江上游的干热河谷灌丛生境,属邻域分布物种,环境梯度变异小,且栖息生境中天敌及食物资源也具有较大相似性,2 个物种面临相似的选择压力,因此,推测2 个物种的反捕食策略(逃逸行为等)可能较为相似,该假说在安乐蜥Anolis cristatellus(Cooper,2010)和鸟类(Tätteet al.,2020)中也获得了证实。
3.3 动作信号与反捕食功能
蜥蜴类物种频繁使用的动态视觉信号(肢体动作信号)具有警示捕食者的功能,可有效降低因逃逸活动带来的能量消耗,如安乐蜥的俯卧撑动作(Leal,1999)、普通壁蜥的摆腿动作(Fontet al.,2012)。本研究也关注了巴塘龙蜥和丽喉龙蜥求偶和领地守卫活动中频繁使用的肢体动作(点头和俯卧撑),发现这2 种动作并未出现在警戒活动中,可能并不具有反捕食的警示功能,仅仅是一种社交信号,这与青海沙蜥的研究结果相似,摆尾和甩尾动作虽然在社交活动中占据主导地位,但与其反捕食策略无关(Qiet al.,2014)。
3.4 后续研究展望
龙蜥是横断山区干热河谷灌丛区域的代表性物种,对于维持区域生态系统的稳定具有重要意义。近年来,随着该区域人类活动的不断加剧,各种道路、水利以及生活设施大量修建已较大程度影响了本区域龙蜥物种的繁衍栖息。后续将持续关注其逃逸行为和抗干扰能力,从单一物种比较(季节、年龄、干扰强度等)到多物种比较,探讨在当前人类活动逐渐增强的背景下对环境干扰的适应能力,为横断山区龙蜥物种乃至其他类群动物的保护工作提供一定的理论依据。
