干旱胁迫对不同抗旱类型芜菁苗期生理特性的影响
2023-05-27高亚宁轩正英张凯浩阿依买木沙吾提马国财
高亚宁,轩正英,张凯浩,李 玉,常 娇,阿依买木·沙吾提,马国财
(1.塔里木大学园艺与林学学院,新疆 阿拉尔 843300;2.塔里木大学南疆特色果树高效优质栽培与深加工技术 国家地方联合工程实验室,新疆 阿拉尔 843300;3.塔里木大学分析测试中心,新疆 阿拉尔 843300)
芜菁(Brassicarapa.L)别名蔓菁、圆根、盘菜等,由油用亚种演化而来,隶属于十字花科芸薹属[1]。其维吾尔语又称为恰玛古,是生长在新疆天山南部、塔里木盆地西北,具有药用、食用、饲用三大应用价值的2 a生草本植物[2-3]。新疆属于极旱地区,土地沙漠化严重、日照时间长、水分蒸发量大,大多数土地为山脉、戈壁和沙漠[4]。芜菁虽耐瘠薄、耐干旱,有较强的适应性,但长时间的干旱少雨仍会在一定程度上对芜菁的叶片及肉质根生长发育造成严重影响。众多研究表明,干旱胁迫可引起植物的生理生化过程损害,如细胞膜结构[5]、关键酶活性[6]、活性氧合成与积累[7]、气孔调节和其他气体交换特性[8]。植物细胞可以调控基因的表达、产生新的蛋白质,进而引起植物生理和代谢上的变化来应对干旱胁迫[9]。例如降低净光合速率,改变植物代谢途径,累积可溶性物质,合成脯氨酸、甜菜碱等渗透调节物质,分解体内原有的蛋白质,同时合成参与各种代谢调节相关的酶[10]。Jabeen等[11]研究表明干旱胁迫导致芜菁的生长量(根和茎的干质量与鲜质量、根长与茎长)、色素含量(叶绿素a,b和总叶绿素)、总酚和抗坏血酸含量减少,显著增强了芜菁叶片与根部的游离脯氨酸、甜菜碱、丙二醛、过氧化氢的含量以及提高了其过氧化氢酶和超氧化物歧化酶活性。
当前在芜菁的营养品质、遗传多样性和非生物胁迫等多方面都开展了大量的研究[1-3,11],但鲜有关于芜菁抗旱性方面的研究。本试验选取前期筛选出的1个抗旱型资源和1个干旱敏感型资源,采用盆栽自然干旱法,研究不同抗旱类型芜菁在苗期干旱胁迫下的生理生化变化,旨为深入研究芜菁抗旱生理机制奠定基础,为芜菁抗旱品种选育及抗旱栽培提供理论依据。
1 材料与方法
1.1 试验设计
供试材料为本课题组前期获得的干旱敏感型资源WJC106和抗旱型资源WJC129,采取盆栽自然干旱的方法进行试验。于2021年8月在塔里木大学园艺试验站智能温室中进行育苗。分别将种子播种于装有125 g混合基质(草炭、蛭石、园土按2∶2∶1体积混合)的营养钵中(每个营养钵播3粒),每份试材共播90个营养钵。育苗期间采取正常管理,当幼苗长出第2片真叶时进行定苗,每盆保留1株苗。待幼苗长至四叶一心时,采用自然干旱的方法进行干旱处理。共设置对照组(CK)和干旱组(T)两个处理,各3次重复,每个重复15株。对照组:土壤相对含水量保持在70%~80%之间;干旱组:干旱处理前浇一次透水,之后不再浇水让其自然干旱。直到幼苗叶片出现严重萎蔫时(干旱第10 d)试验结束。
1.2 测定指标与方法
干旱处理结束后采用组内随机取样的方法,选取植株第4片长势一致的叶片测定其生理指标,每个重复5片,每个处理3次重复共15片叶。参考李合生[12]的方法测定其相对电导率、总叶绿素、可溶性糖、可溶性蛋白、丙二醛和游离脯氨酸含量。采用苏州科铭生物技术有限公司生产的试剂盒测定超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性和过氧化氢酶(CAT)活性。
采用发射扫描电子显微镜(APREO-S,Thermo Fisher Scientific公司,美国)对叶片的气孔特征进行观察,并用Image Pro Plus软件进行指标测量。每个叶片随机选取10个视野,主要测量气孔张开长度、气孔张开宽度、气孔长度、气孔宽度、气孔开度、保卫细胞厚度、气孔密度。气孔开度=气孔张开长度×气孔开张宽度×π/4,气孔密度=视野下气孔数/视野下的面积[13]。
1.3 数据处理
采用Microsoft excel 2019软件进行数据整理,用SPSS 25.0及 Origin 9.0软件对试验数据进行分析及作图。
2 结果与分析
2.1 干旱胁迫对不同抗旱类型芜菁苗期丙二醛含量及相对电导率的影响
丙二醛作为膜脂过氧化的最终分解产物,可反映膜系统的受伤害程度和抗旱性强弱,同时对细胞产生毒害作用[13];相对电导率是衡量细胞膜透性的重要指标[14-15]。如图1所示,两个芜菁资源在干旱胁迫下,其苗期叶片丙二醛含量和相对电导率相对于对照组均呈现出显著上升趋势。图1A显示,与对照组相比,WJC106在干旱胁迫下的丙二醛增量显著高于WJC129,增幅分别为59.71%和54.52%。图1B显示,WJC106和WJC129在干旱胁迫下相对电导率值与对照组相比,分别增加了27.05%和20.06%。干旱胁迫下,干旱敏感型的WJC106芜菁苗期细胞膜受损程度大于抗旱型的WJC129。
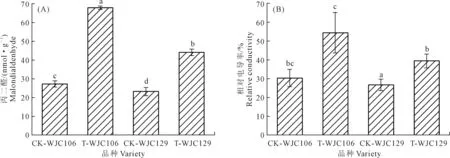
注:不同小写字母表示处理间差异显著(P<0.05),下同。Note:Different lowercase letters indicate significant differences between treatments at P<0.05,the same below.图1 干旱胁迫对芜菁苗期丙二醛含量及相对电导率的影响Fig.1 Effects of drought stress on MDA and relative electrical conductivity of turnips seedling
2.2 干旱胁迫对不同抗旱类型芜菁苗期抗氧化酶活性的影响
如图2所示,干旱胁迫使两个不同抗旱类型芜菁苗期的SOD、POD和CAT活性增加。图2A、2B表明,与对照组相比,WJC129芜菁幼苗叶片的SOD、POD活性受干旱胁迫的影响呈显著增加趋势,而WJC106叶片的SOD和POD活性与对照组无显著差异。从图2C可以看出,相比于对照组,两个不同抗旱类型芜菁在干旱胁迫下的CAT活性均显著增加,且干旱下WJC129的CAT活性显著大于WJC106。抗旱型的WJC129芜菁苗期保护酶活性变化大于干旱敏感型的WJC106。
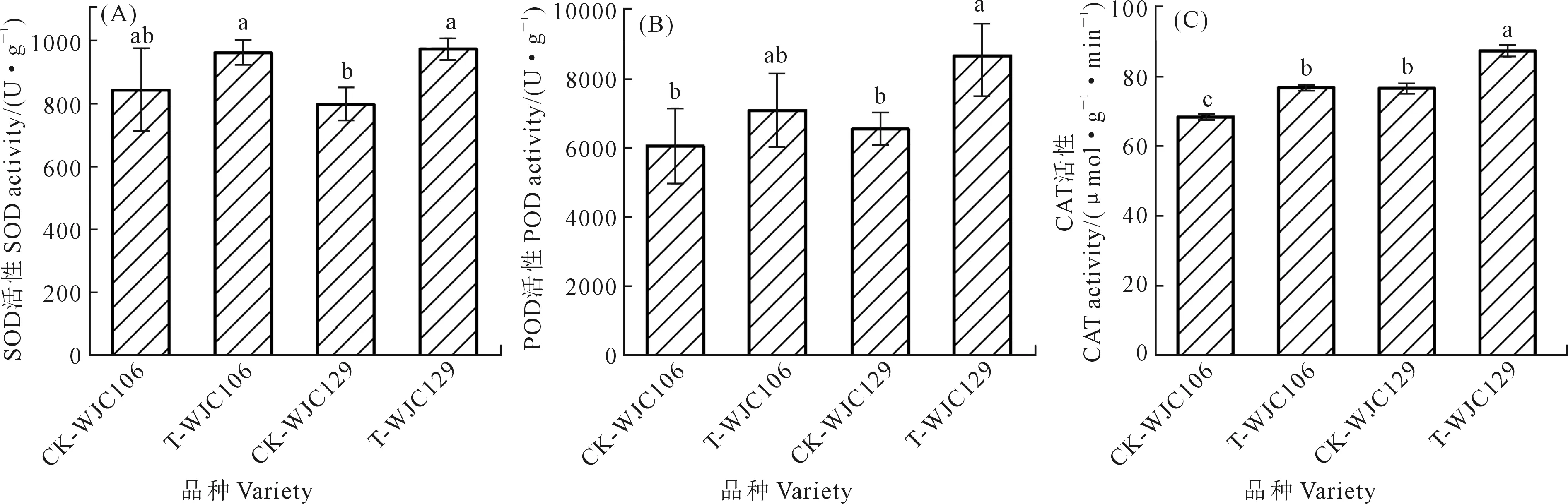
图2 干旱胁迫对芜菁苗期抗氧化酶活性的影响Fig.2 Effects of drought stress on antioxidant enzyme activity of turnips seedling
2.3 干旱胁迫对不同抗旱类型芜菁苗期渗透调节的影响
图3A、3B显示,干旱胁迫下WJC106和WJC129两个芜菁的脯氨酸和可溶性蛋白含量均显著高于对照组,其脯氨酸增量分别为85.22%和88.97%,可溶性蛋白增量为32.51%和58.26%。图3C显示,干旱条件下两个不同抗旱类型芜菁苗期叶片可溶性糖含量均增加,但无显著差异。由此可知,干旱胁迫使芜菁苗期渗透调节物质含量增加,且WJC129渗透调节物质变化量高于WJC106,抗旱型芜菁苗期在受到干旱胁迫时的渗透调节能力强于干旱敏感型;同时脯氨酸和可溶性蛋白在干旱胁迫下较为敏感,起主要渗透调节作用。
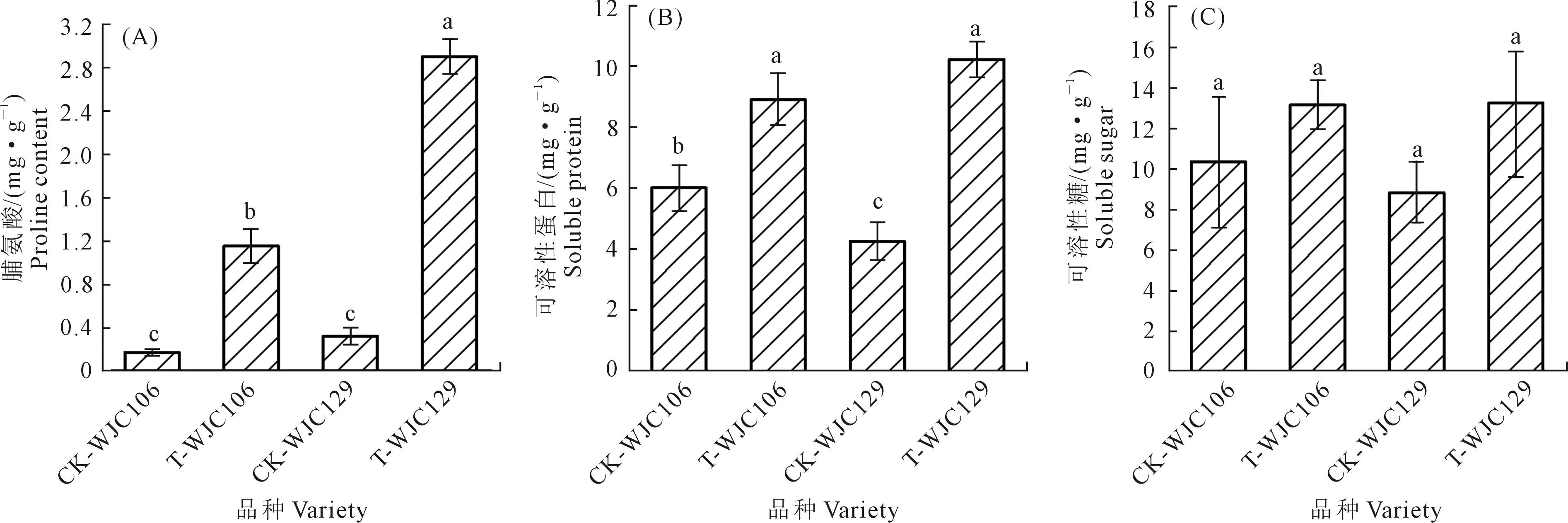
图3 干旱胁迫对芜菁苗期渗透调节物质含量的影响Fig.3 Effects of drought stress on contents of osmotic regulation substances of turnips seedling
2.4 干旱胁迫对不同抗旱类型芜菁幼苗叶绿素含量及光合荧光参数的影响
如表1所示,干旱胁迫使两个不同抗旱类型芜菁苗期叶绿素含量显著降低,且WJC129叶绿素含量高于WJC106。与对照组相比,两个芜菁处理组Pn和Tr呈现出显著降低。干旱胁迫下WJC106的Gs值显著低于对照组,而WJC129对照组与干旱组的Gs值无显著差异。两个不同抗旱类型芜菁Ci变化量不尽相同,抗旱型WJC129芜菁在干旱胁迫下呈现为下降趋势,而干旱敏感型WJC106表现为显著升高。两个资源的水分利用效率变化量也有不同变化趋势,在干旱胁迫的影响下,WJC106干旱敏感型芜菁的WUE表现为显著降低,而WJC129抗旱型资源出现显著增加的趋势。

表1 干旱胁迫对芜菁苗期叶绿素含量及光合参数的影响Table 1 Effects of drought stress on chlorophyll and photosynthetic parameters of turnips seedling
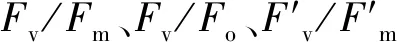
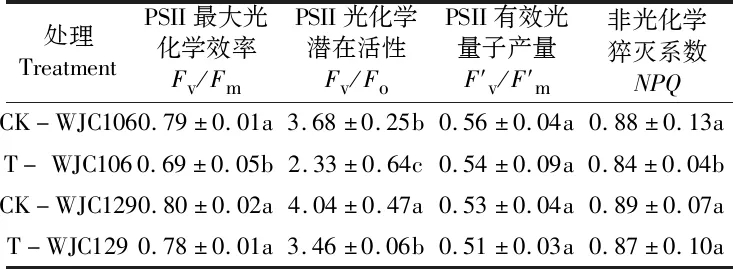
表2 干旱胁迫对芜菁苗期荧光特性的影响Table 2 Effects of drought stress on fluorescence characteristics of turnips seedling
2.5 干旱胁迫对不同抗旱类型芜菁苗期叶片气孔形态的影响
表3为干旱胁迫对不同抗旱类型芜菁苗期叶片气孔特征值的影响,图4为干旱胁迫对芜菁苗期叶片气孔形态的影响。由表3和图4(A~D)可知,干旱胁迫使两个不同抗旱类型的芜菁苗期叶片气孔张开长度、气孔张开宽度、气孔长度、气孔宽度、保卫细胞厚度和气孔开度极显著降低。干旱胁迫下,WJC129抗旱型芜菁苗期叶片气孔开度的降幅显著小于WJC106,降幅分别为65.90%和87.32%,并且气孔张开长度、气孔张开宽度、气孔长度、气孔宽度和保卫细胞厚度降幅也是WJC129明显小于WJC106。表3与图4(E~H)结合更直观可知,干旱胁迫下WJC106芜菁苗期叶片的气孔张开率极显著降低,而WJC129芜菁无明显差异。从气孔密度上看,WJC106呈极显著增加,WJC129无显著差异。干旱敏感型WJC106芜菁苗期叶片受干旱胁迫的损害较大。从图4(C、D)和图4(G、H)可以观察到WJC129抗旱型芜菁叶片表面有蜡质层覆盖,蜡质晶体呈管状形态。
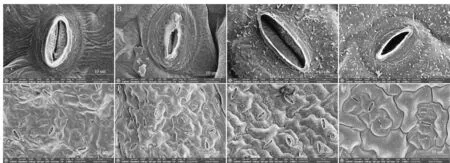
注:A为对照组WJC106芜菁苗期叶片下表皮单个气孔形态;B为干旱胁迫下WJC106芜菁苗期叶片下表皮单个气孔形态;C为对照组WJC129芜菁苗期叶片下表皮单个气孔形态;D为干旱胁迫下WJC129芜菁苗期叶片下表皮单个气孔形态;E为对照组WJC106芜菁苗期叶片下表皮气孔形态;F为干旱胁迫下WJC106芜菁苗期叶片下表皮气孔形态;G为对照组WJC129芜菁苗期叶片下表皮气孔形态;H为干旱胁迫下WJC129芜菁苗期叶片下表皮气孔形态。Note:B:A single stomatal morphology of the epidermis of the leaves at the seedling stage of turnip WJC106 in the control group;B:A single stomata morphology of the epidermis of the epidermis of the leaves at the seedling stage of turnip WJC106 under drought stress;C:A single stomatal morphology of the epidermis of the leaves at the seedling stage of turnip WJC129 in the control group;D:A single stomata morphology of the epidermis of the leaves at the seedling stage of turnip WJC129 under drought stress;E:The stomata morphology of the epidermis at the seedling stage of the control group WJC106 turnip;F:The stomata morphology of the epidermis of the leaves at the seedling stage of turnip WJC106 under drought stress;G:The stomata morphology of the epidermis of the leaves at the seedling stage of the control group WJC129 turnip;H:The stomata morphology of the epidermis of the leaves at the seedling stage of turnip WJC129 under drought stress.图4 干旱胁迫对芜菁苗期叶片气孔特征的影响Fig 4 Effects of drought stress on stomatal eigenvalue of turnips seedling
3 讨论与结论
环境在植物的生长发育过程中起着至关重要的作用,而水分因子更是占具主导地位[16]。本试验表明,干旱胁迫使芜菁苗期的丙二醛含量和相对电导率显著增加,细胞膜结构受损,这与前人研究结果一致[11]。本试验中WJC106干旱敏感型芜菁苗期叶片的丙二醛含量和相对电导率的增量显著高于WJC129抗旱型资源。干旱敏感型的芜菁苗期细胞膜受损程度大于抗旱型,严重影响细胞中物质的进出,破坏细胞代谢的稳定性,同时表明芜菁幼苗的抗旱性越强,电解质外渗量小,膜脂过氧化程度越低,其相对电导率和MDA积累量越低。
当植物受到干旱胁迫时,干旱信号会促进脯氨酸和可溶性糖等代谢物质的产生,激发抗氧化系统以维持氧化还原稳态,并通过过氧化物酶防止细胞损伤和膜完整性的破坏[17]。SOD、POD和CAT是保护酶系统中最重要的三种酶,其中SOD催化两个超氧自由基发生歧化反应形成O2和H2O2,产生的H2O2再由POD和CAT分解除去[18]。三种酶协同作用有助于抵抗氧化损伤,维持细胞膜的稳定性和完整性。本研究中,干旱胁迫使两个不同抗旱类型芜菁苗期的SOD、POD和CAT活性增加,并且抗旱型芜菁抗氧化酶活性的增幅显著大于干旱敏感型。可溶性蛋白、可溶性糖和脯氨酸是植物主要的渗透调节物质,干旱胁迫下可通过渗透调节使植物维持一定的膨压,从而维持细胞生长、气孔开放和光合作用等生理过程、增强抗旱性[19]。本研究中两个不同抗旱类型芜菁苗期的脯氨酸、可溶性糖和可溶性蛋白含量在干旱条件下均表现出增加趋势,但可溶性糖增幅差异不显著,且抗旱型WJC129渗透调节物质含量高于干旱敏感型WJC106,表明芜菁苗期可通过增加其渗透调节物质提高其抗旱性,且脯氨酸和可溶性蛋白在干旱胁迫下较为敏感,起主要的渗透调节作用。
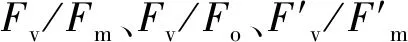
气孔是植物进行气体交换的重要器官,是水汽蒸腾和CO2进出的主要通道,受干旱影响,气孔可能会关闭[24],以减少水分损失。本研究中干旱胁迫使两个不同抗旱类型的芜菁苗期叶片气孔变小,保卫细胞变薄,气孔开度显著减小。抗旱型芜菁气孔张开率显著大于干旱敏感型的,仍能维持较高的光合作用。干旱状态下干旱敏感型WJC106芜菁叶片表皮细胞出现重叠褶皱和起伏,影响气孔开度和植物蒸腾,而抗旱型WJC129表皮细胞无明显起伏,并且其表面附有蜡质层,蜡质晶体呈管状形态。前人研究表明,蜡质是通过防止非气孔性水分的流失,使得植物体蒸腾作用得以下降,从而能够更好地维持植物体的水分来抵御干旱胁迫[25]。故抗旱型WJC129通过表面附有的蜡质层来增加其抗旱性。
综上所述,干旱敏感型芜菁苗期细胞膜受损程度大于抗旱型,且抗旱型芜菁有较强的自我恢复与协调能力,通过增加抗氧化酶活性和渗透调节物质含量来抵御干旱胁迫。渗透调节物质中,脯氨酸和可溶性蛋白在干旱胁迫下较为敏感,起主要的调节作用。干旱胁迫降低芜菁的光合荧光参数,气孔特征值下降,光合机构活性降低;抗旱型芜菁的各光合特性指标降幅均低于干旱敏感型,气孔特征值也显著大于干旱敏感型,故仍能维持较高的光合作用。抗旱型芜菁表皮附有蜡质层,从而增加其抗旱性。
