河西走廊东段不同生境对沙米形态结构及生物量分配的影响
2023-05-27姜生秀
姜生秀,赵 鹏
(甘肃省治沙研究所,甘肃省荒漠化与风沙灾害防治国家重点实验室(培育基地),甘肃 兰州 730070)
植物形态结构和生物量分配特征是植物和环境共同作用的结果,是植物生长发育规律及其对环境变化响应的直接体现[1]。生物量分配是指植物生长发育过程中所同化的资源用于根、茎、叶、花、果等各器官的比例,也就是各器官累积的资源在总资源中所占的比例[2]。植物在不同的生境条件下其生长特性与生物量分配规律也不同[3]。植物在其生长过程中,不同器官中的生物量分配不是固定的,这不仅是植物自身生长特性的体现,也反映了器官生物量分配对土壤水分、养分、光照、温度的响应[4]。例如在土壤水分或养分含量低的情况下,植物通过促进根系的伸长生长来增加根系生物量的分配[5];在光照资源缺乏而土壤水分和养分充足时,植物快速生长,株高的增加会使更多的生物量分配到地上部分以增加光合作用[6]。因此,研究不同生境条件下植物形态特征、生物量分配和二者相关性有助于理解全球变化背景下环境因子对植物生长发育、生活史进程、种群稳定性及分布的影响。
沙米(Agriophyllumsquarrosum)属藜科(Chenopodiaceae)沙蓬属(Agriophyllum)一年生植物,具有抗干旱和盐碱、耐贫瘠和风蚀沙埋等特性,适应能力强,是流动沙丘恢复过程中的主要先锋物种[7]。因其独特的生理生态特性,在流动沙丘上易定居、而且生长速度极快,分布比较集中密集,当其覆盖度达到70%以上时,对风沙流就能达到很好的控制效果,从而流动沙丘被固定、不被风蚀[8]。另外,沙米种子富含蛋白质、脂肪、碳水化合物、粗纤维和灰分,营养价值极高,可加工成食品供人食用[9]。骆驼和山羊喜食嫩茎叶,是良好的饲用植物。国内外有关沙米的研究报道甚多,多集中在种子萌发[10-11]、幼苗生长[12]、生理生化[13-14]、形态结构[8,11]及营养价值[7,12]等方面,但是沙米生物量分配特性以及这些特性与环境因子的关系还不清楚。因此,本文研究甘肃民勤、武威和古浪3个不同生境下沙米形态特征和生物量分配及其与环境之间的相关性,将有助于了解荒漠地区1 a生植物对环境变化的响应机制及适应策略。
1 材料与方法
1.1 研究区概况
研究区位于腾格里沙漠东南缘的甘肃省武威市长城乡、民勤县西沙窝和古浪县海子滩,民勤县西沙窝属于典型的温带大陆性荒漠气候,年均气温7.4℃,极端低温-30.8℃,极端高温40.0℃,年均降雨量113.2 mm。武威市长城乡属于典型的温带半荒漠大陆性气候,年均气温7.7℃,年均降雨量 139.9 mm,年均蒸发量2 486 mm。古浪县海子滩属于典型的温带半荒漠大陆性气候,海拔1 620 m,年均气温6.8℃,年均降雨量为170 mm,主要集中在7—9月,年均蒸发量为2 807 mm。武威、民勤和古浪年均降雨量和年均气温均存在较大差异,因此选择这3个生境作为研究区域。
1.2 样地设置与采样
选择流动沙丘作为试验样地,在3个生境中根据沙丘走向在植被分布相对均匀处设置1 m×1 m 的3个样方。样品采集工作于2020年6—9月进行,于沙米开花末期,在每个小样方内分别选择沙米20~30株,将每株沙米的根、茎、叶、花分离出来,置于80℃烘箱内烘干称重,并计算根冠比。在每个小样方内利用对角线取样法采集0~30 cm的混合土样,再将土样风干,带回进行理化分析,采用凯氏定氮法测定土壤全氮含量;采用重铬酸钾稀释热法测定土壤有机质;采用钼锑抗比色法测定速效磷含量。剔除土壤样品的杂物,过筛后分别测定并计算土壤砂粒(2~0.05 mm)、粉粒(0.05~0.002 mm)、粘粒(<0.002 mm)含量。在每个小样方用土钻取0~5、5~10、10~20、20~30 cm层土壤样品,带回实验室用烘干法测量每层土壤含水量,最后将各土层的含水量合在一起求平均值。土壤日均温度由土壤温度计测得,在每个小样方地表下10 cm处放置1个温度计,每0.5 h测定1次温度,连续监测24 h,最后取其平均值。收集2020年3个生境的气象资料,得到年均气温、年均日照时数和年太阳总辐射数据。
1.3 数据处理
利用Excel 2007软件对实验数据进行整理和统计。采用Prism 5.0软件进行方差分析和显著性检验。植物形态与生物量指标间进行Pearson相关性分析。异速生长关系采用Y=β·Xα模型进行研究,将两边取对数后(以10为底数)转化为lgy=lgβ+αlgx,其中β为标准化常数,α为异速生长指数,α=1为等速关系,α≠1为异速生长关系。根据Warton等[15]方法对α 进行异质性检验,如果α没有异质性,则给出一个共同的斜率;若α不同,则异速生长关系完全不同。采用标准化主轴估计(Standardized major axisestimation,SMA)方法确定相关生长模型的参数,使用SMAR-TR软件完成等速生长检验、共同斜率检验及α的方差分析。
2 结果与分析
2.1 不同生境中主要环境因子
由表1可以看出,武威年均气温最高,为8.58℃,和民勤之间无显著性差异(P>0.05),但二者均显著高于古浪(P<0.05);武威年均日照时数最高(2 873.4 h)、民勤次之(2 833.0 h)、古浪最小(2 669.0 h),但三者之间无显著性差异(P>0.05);民勤年太阳总辐射最高(6 330 MJ·m-2),显著高于武威和古浪(P<0.05);不同生境土壤日均温度之间存在显著性差异(P<0.05),武威最高、民勤次之、古浪最低;古浪土壤含水量和土壤粉粒均最高,分别为4.53%和17.51%,均显著高于武威和民勤(P<0.05),武威土壤含水量和土壤粉粒均显著高于民勤(P<0.05);3个生境的土壤粘粒和土壤砂粒之间均无显著性差异(P>0.05);土壤有机质和土壤全氮在不同生境变化趋势一致,均是古浪最高、民勤次之、武威最低,且3个生境之间均差异显著(P<0.05);古浪土壤速效磷最高(6.81 g·kg-1),显著高于武威和民勤(P<0.05),而武威和民勤之间无显著性差异(P>0.05)。整体来看,3个生境相比,古浪气温较低,日照时数和太阳辐射较小,但土壤含水量较好,土壤肥力最佳;民勤太阳辐射最高,土壤含水量最小;武威和民勤土壤肥力接近。
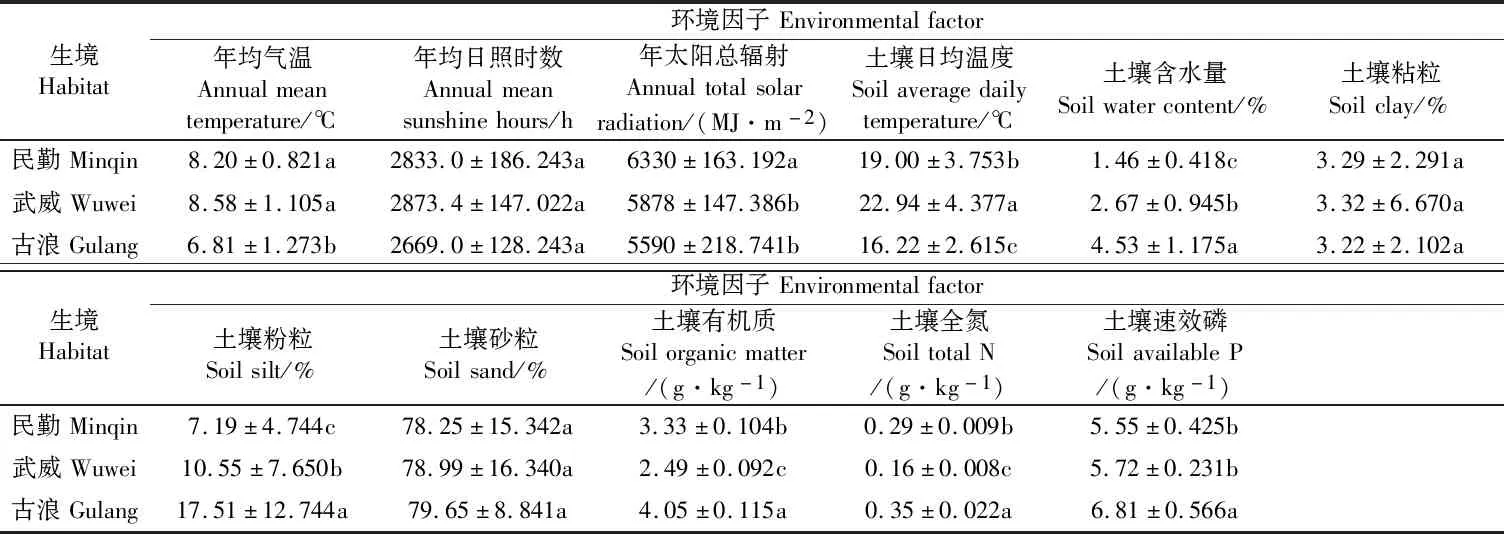
表1 不同生境中主要环境因子Table 1 The main environmental factors in different habitats
2.2 不同生境下沙米的形态特征
由表2可以看出,不同生境沙米株高、冠幅之间均存在显著性差异(P<0.05),古浪株高最大(70.63 cm)、武威次之(67.59 cm)、民勤最小(62.40 cm),冠幅和株高的变化趋势具有一致性,株高越大,冠幅越大;基径在3个生境中无显著性差异(P>0.05);古浪沙米根长最大(96.15 cm)、武威次之(90.75 cm)、民勤最小(86.31 cm),3个生境之间均存在显著性差异(P<0.05)。由此说明,古浪生境下沙米生长状态良好。
2.3 不同生境下沙米生物量及其分配特征
由表3可以看出,古浪沙米根生物量最大(7.56 g),显著高于武威和民勤(P<0.05),比武威和民勤分别高出41.67%和41.40%,武威和民勤之间无显著性差异(P>0.05);古浪沙米具有较高的根冠比(0.291),武威次之(0.199)、民勤最小(0.198);武威沙米叶生物量较小(19.80 g),和民勤之间无显著性差异(P>0.05),但二者均显著小于古浪(P<0.05);茎和花生物量在3个生境之间均无显著性差异(P>0.05);总生物量古浪最高(88.07 g)、民勤次之(81.40 g)、武威最小(77.25 g)。

表3 不同生境条件下沙米各器官生物量、总生物量及根冠比Table 3 Biomass of different organs,total biomass and root/shoot ration of A. squarrosum in different habitats
同一生境下,民勤和武威各器官生物量质量分数均表现为茎>花>叶>根,古浪表现为茎>叶>花>根。茎的质量分数均最高,民勤、武威和古浪茎生物量分配比分别为43.21%、40.67%和33.68%;根生物量分配比古浪最高(15.98%)、武威次之(5.71%)、民勤最小(5.44%);不同生境中叶生物量分配比相近;武威花生物量分配比为27.98%,民勤和古浪分别为25.77%和24.01%(图1)。

图1 不同生境中沙米各器官生物量百分比Fig.1 Biomass partitioning of A. squarrosum organs in different habitats
2.4 不同生境下沙米器官生物量间相关生长关系
由图2和表4(见253 页)可以看出,3个生境沙米根-茎、根-叶和根-花生物量均呈α<1的异速生长关系,即根生物量积累速率大于茎、叶和花生物量的积累速率,进一步等速生长检验表明,沙米在不同生境中根-茎生物量α有显著性差异(P=0.0248),不存在共同斜率,根-叶生物量α无显著性差异(P=0.1708),存在共同斜率(α=0.0788),根-花生物量α无显著性差异(P=0.0997),存在共同斜率(α=0.074);3个生境沙米叶-茎生物量(P=0.2476)及花-茎生物量(P=0.6259)α均无显著性差异,均存在共同斜率(α=0.1501和α=0.0784),且均是α<1的异速生长关系,即叶和花生物量积累速率均大于茎;不同生境沙米花-叶生物量α有显著性差异(P=0.0373),不存在共同斜率,且均是α<1的异速生长关系,即花的生物量积累速率大于叶的生物量积累速率。由此说明,沙米生长过程中地下部生物量积累速率大于地上部生物量,在地上部生物量中,生殖器官生物量积累速率大于营养器官生物量。
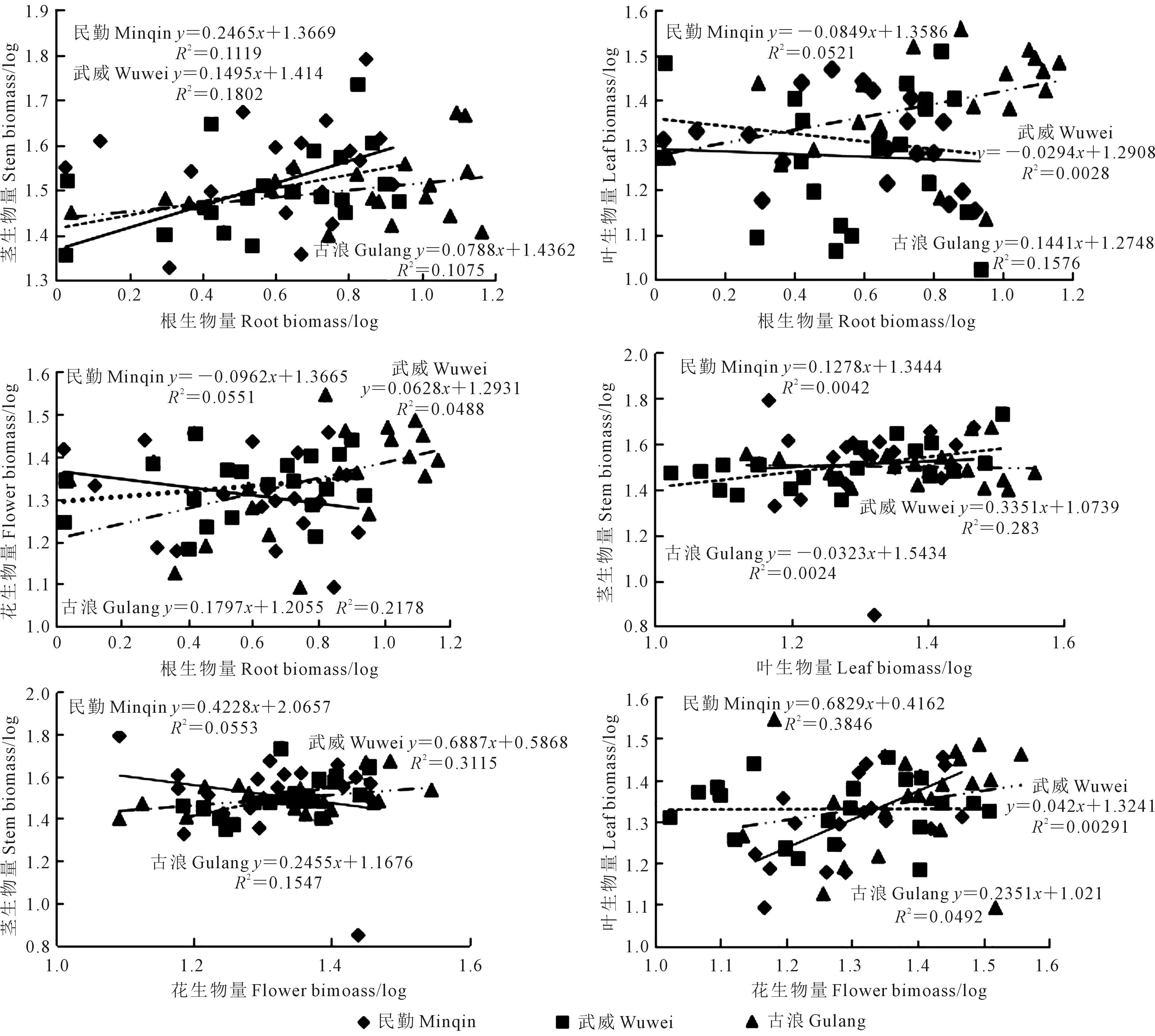
图2 不同生境沙米叶、茎、根、花生物量之间的异速生长关系Fig.2 Allometric relationships between leaf,stem,root and flower biomass of A. squarrosum in different habitats
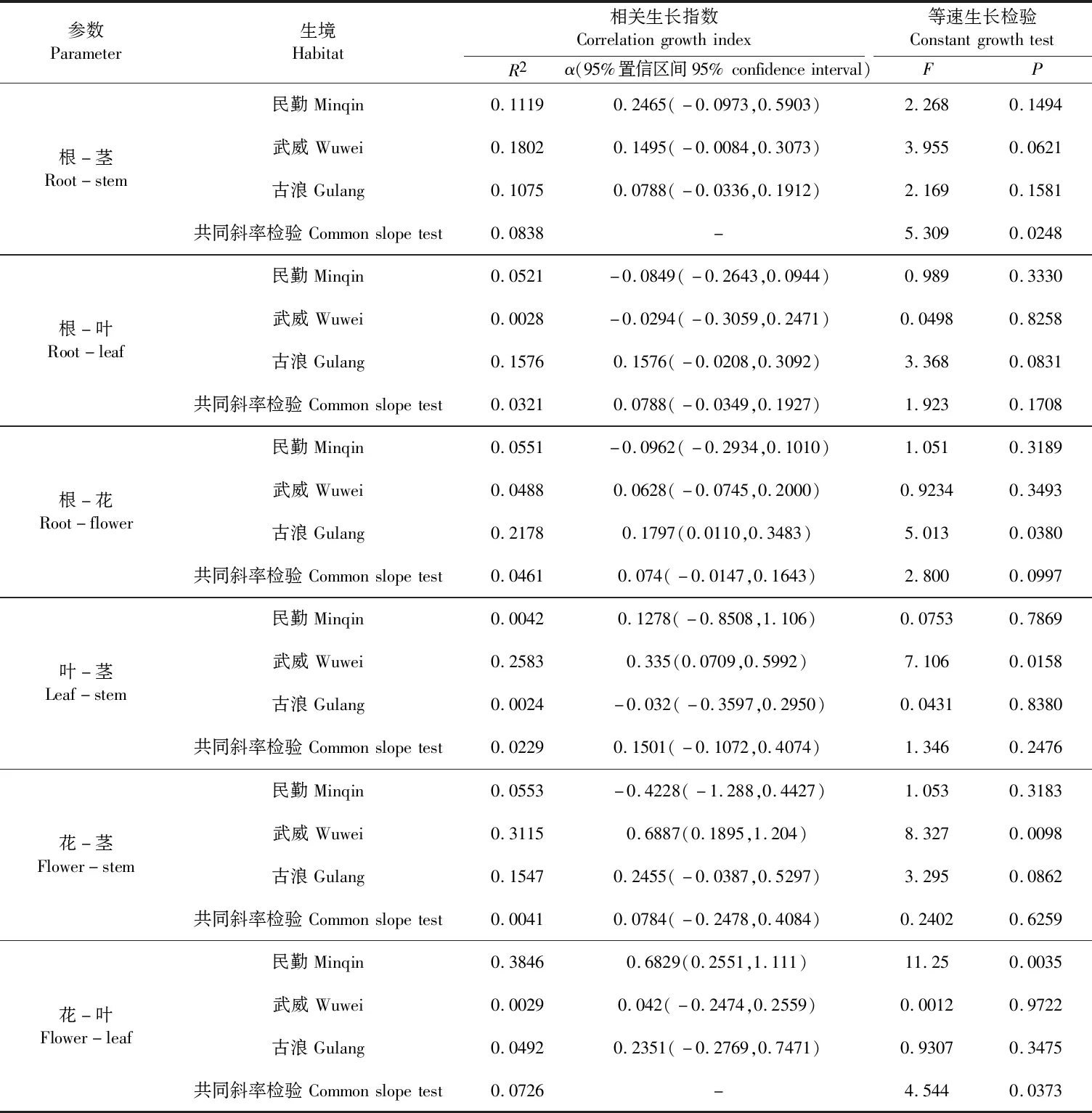
表4 不同生境沙米叶、茎、根、花生物量间的拟合参数、等速生长检验及共同斜率检验Table 4 Fitting parameters,isometric growth test and common slope test of leaf,stem,root and flower biomass of A. squarrosum in different habitats
2.5 不同生境下沙米形态与个体生物量间的相关性
由表5(见253 页)可以看出,株高和根、茎生物量在3个生境中均呈显著正相关(P<0.05),和叶、花生物量在民勤和武威均呈显著正相关(P<0.05),在古浪呈极显著正相关(P<0.01),民勤(r=-0.742)和武威株高与根冠比呈显著负相关(r=-0.698),古浪呈极显著负相关(r=-0.852,P<0.01),表明随着沙米植株的增大根冠比呈减小的趋势;根长与根生物量在民勤呈极显著正相关(r=0.902,P<0.01),在武威(r=0.752)和古浪呈显著正相关(r=0.635),3个生境中根长与茎、叶、花生物量之间均无显著相关性,与根冠比仅在古浪呈显著负相关(r=-0.772,P<0.05);冠幅与根、茎生物量在3个生境中相关性均不显著,而与叶生物量及根冠比均呈显著正相关(P<0.05),与花生物量均呈极显著正相关(P<0.01)。由此说明,不同生境沙米的形态特征与个体生物量间具有协同变化的趋势。
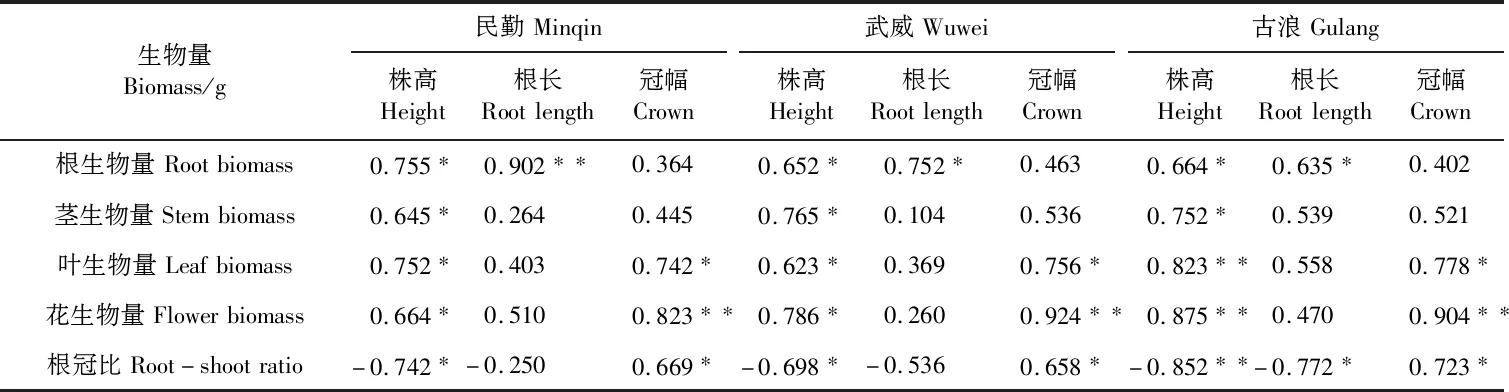
表5 不同生境沙米株高、冠幅、根长与个体生物量间的相关性分析(样本量n=9)Table 5 Correlation between plant height,crown width,root length and individual biomass of A. squarrosum in different habitats(Sample size n=9)
2.6 器官生物量与环境因子的相关性
逐步回归分析表明(表6,见254页),年均气温和土壤日均温度均极显著影响沙米根生物量和根冠比(负相关,P<0.01),均显著影响茎生物量(正相关,P<0.05),即气温和土壤温度越低,沙米植株根生物量和根冠比越大,茎生物量就越小;年均日照时数、年太阳总辐射和叶生物量之间均呈显著负相关(P<80.05),和花生物量间均呈极显著负相关(P<0.01)。土壤含水量与总生物量、根生物量及根冠比均呈显著正相关(P<0.05);土壤有机质对总生物量、叶和花生物量具有显著影响(正相关,P<0.05)、对根生物量和根冠比具有极显著影响(正相关,P<0.01);土壤全氮和总生物量、根和叶生物量及根冠比之间均呈显著正相关(P<0.05);土壤粉粒与根、叶生物量及根冠比之间均呈显著正相关(P<0.05);除此之外,土壤速效磷、土壤粘粒和土壤砂粒对沙米各器官生物量均不产生显著影响。
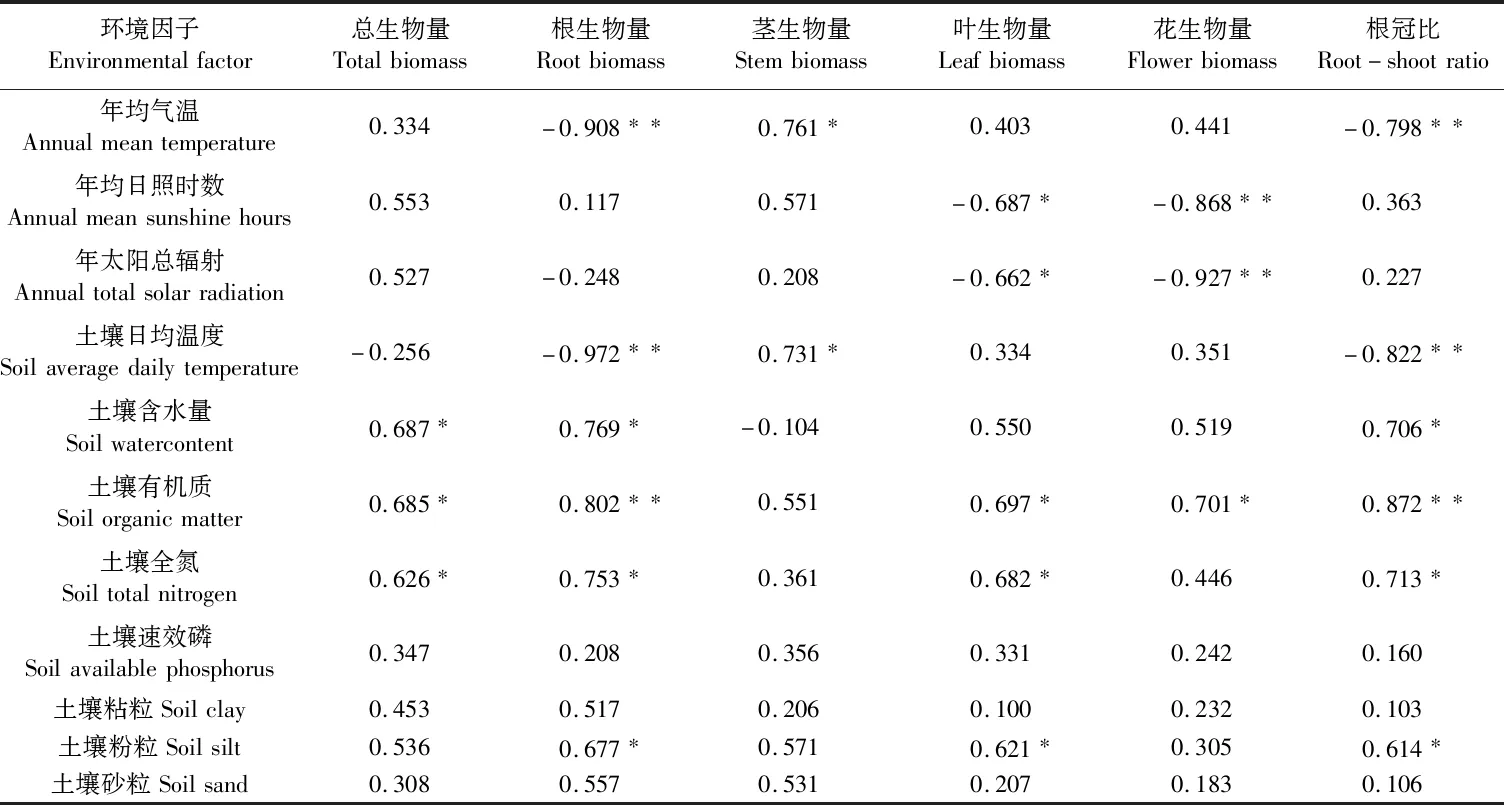
表6 器官生物量与环境因子的相关性分析(样本量n=9)Table 6 Correlation analysis between organ biomass and environmental factors (Sample size n=9)
3 讨论与结论
3.1 不同生境沙米形态特征差异
植物经过长期的历史进化,逐渐形成了独特的生长发育及繁殖策略,以增强其对异质生境的适应性[16]。在甘肃民勤、武威和古浪3个生境中,光照、气温、土壤水分、养分和温度等环境因子存在较大的差异,对植物的生长动态产生了较大的影响。不同生境条件下沙米株高、冠幅与根长均表现为古浪>武威>民勤,基径在3个生境中无显著性差异(P>0.05),结合3个生境的环境因子来看,古浪土壤含水量(4.53%)、土壤粉粒(17.51%)、土壤有机质(4.05 g·kg-1)和土壤全氮(0.35 g·kg-1)及土壤速效磷(6.81 g·kg-1)均最高,武威土壤含水量(2.67%)、土壤粉粒(10.55%)及土壤速效磷(5.72 g·kg-1)均高于民勤(1.46%、7.19%和5.55 g·kg-1),间接表明了古浪土壤肥力最好,土壤有机质分解及矿化过程更快,能够较快地促进植物生长发育;另外,土壤养分和水分共同作用影响沙米植株的生长。沙米的株高也与其对光的竞争有关,在本研究中,年均日照时数和年太阳总辐射古浪最低(2 669 h和5 590 MJ·m-2),沙米通过增加其高度使其对光的捕获能力增强[17]。有研究表明,植物在外部形态结构及生长特性方面都能表现出对环境的适应性。洪岚等[18]研究发现,薇甘菊(Mikaniamicrantha)在光照充分的开阔地中能获得更高的生长量。黄迎新等[19]研究表明,土壤养分和水分的增加有利于沙米的生长,并且土壤养分对沙米生长的影响最大。王艳莉等[20]发现,不同生境中砂蓝刺头(Echinopsgmelinii)的生长不仅受水分和养分条件的影响,还与群落中植物水分利用竞争相关。上述结论和本研究结果一致。
3.2 不同生境下沙米生物量分配特征及其与形态、环境因子的相关性
植物生物量体现了其在生长发育过程中对外界养分的吸收、积累、利用的能力[21]。影响生物量分配的生物和非生物因素有降水量、养分含量、土壤质地、植被类型等,其中温度是影响植物生物量分配非常重要的环境因子[22]。根系作为植物个体的重要组成部分,具有吸收水分、营养物质和储存的作用[15]。本研究发现,3个生境中根生物量在古浪最高(7.56 g)、民勤次之(4.43 g)、武威最小(4.41 g),且古浪沙米具有较高的根冠比(0.291),而年均气温和土壤日均温度古浪均最低(6.81℃和16.22℃),武威均最高(8.58℃和22.94℃)。马冰等[23]研究发现,降温处理过的幼苗株高、叶面积、地上部生物量分配相对较小,而表现出将更多的资源分配给根部,这与本研究结果类似。经逐步回归分析发现,年均气温和土壤日均温度均极显著影响沙米根生物量和根冠比(负相关,P<0.01),这是因为植物为了适应低温环境,通过对地上部分生长一定程度的弱化,将更多光合产物分配到地下部分,使得古浪沙米有更高的根冠比,提高了植物根系对水分及养分的吸收能力,另一方面,在低温环境中,植物根系生物量的损失速率相对较低,这也同样有利于根系生物量的累积,导致较高的根冠比[24]。3个生境中茎的生物量古浪(32.27 g)略小于武威和民勤(32.42 g和35.17 g),这可能是因为古浪温度较低,植物蒸腾作用较小,减小茎的生物量可以更好地适应较低的蒸腾拉力下的水分和养分需求。逐步回归分析也表明,年均气温和土壤日均温度与茎生物量呈显著正相关(P<0.05)。叶生物量古浪最大(25.23 g)、民勤次之(20.82 g)、武威最小(19.80 g),花生物量古浪最大(23.01 g)、武威次之(21.62 g)、民勤最小(20.98 g),这是因为年均日照时数和年太阳总辐射古浪均最小(2 669 h和5 590 MJ·m-2),武威年均日照时数最大(2 873.4 h),民勤年太阳总辐射最大(6 330 MJ·m-2)。有研究表明,当植物幼苗生长受到光照条件的限制,植物将更多的资源分配到地上部分,通过增加其高度、叶分配和比叶面积来获取更多的光照资源用以提高其光竞争能力和生存适合度[25],而强光和高温对花序的生长发育具有阻碍作用,造成花序发育中止或者败育[18]。逐步回归分析也表明,年均日照时数和年太阳总辐射与花生物量呈极显著负相关(P<0.01)、与叶生物量呈显著负相关(P<0.05)。
在本研究中,土壤粉粒(0.002~0.05 mm)与根、叶生物量及根冠比之间均呈显著正相关(P<0.05),土壤粘粒和土壤砂粒对沙米各器官生物量均不产生显著影响。Wang 等[22]研究发现,在草地生态系统中,植物在土壤砂粒 (0.05~2 mm)含量较高的地方其根冠比也较高,而在粘粒(<0.002 mm)含量较高的地方根冠比较低,后者的土壤含水量和养分含量较低而孔隙度较高而使得氧气含量相对充足,植物为了获取更多水分及养分而促进根系生长,这与本研究结果并不完全一致。Muller等[26]研究表明,根冠比与土壤养分呈负相关,而王艳莉等[20]研究发现,土壤水分和养分与根冠比呈正相关。本研究表明,土壤含水量与总生物量、根生物量及根冠比均呈显著正相关(P<0.05),土壤有机质对总生物量、叶和花生物量具有显著影响(正相关,P<0.05)、对根生物量和根冠比具有极显著影响(正相关,P<0.01),土壤全氮和总生物量、根和叶生物量及根冠比之间均呈显著正相关(P<0.05),这就说明当土壤养分或水分吸收受到限制时,将更多的物质能量分配给根系生长。
除了环境因素外,植物大小对沙米的器官生物量分配也产生了显著影响。在不同生境条件下,株高和根、茎、叶、花生物量间均呈显著或极显著正相关,表明这些指标间具有明显的协同生长关系。株高与根冠比呈显著或极显著负相关,说明地下部生物量的分配比例随地上部形态指标增大有减小的趋势,这与陶冶等[27]关于短命植物生物量分配研究结果一致。
3.3 不同生境沙米叶、茎、根和花生物量的异速生长关系
当植物的光合产物向其不同部分的分配关系成非线性比例时,即表现为植物的异速生长关系,它能反映出植物在不同环境下的生长策略[24]。在本研究中,3个生境沙米根-茎、根-叶、根-花、叶-茎、花-茎和花-叶生物量均是α<1的异速生长关系,说明沙米生长过程中根生物量积累速率大于茎、叶、花,叶、花生物量积累速率大于茎,花生物量积累速率大于叶,即沙米倾向于将更多的生物量分配到根部,提高对营养物质和水分的吸收速率以更好地适应干旱荒漠环境。营养器官与生殖器官的生物量分配反映了植物在有性繁殖和无性繁殖之间的资源分配策略[18],在地上生物量中,生殖器官生物量积累速率大于营养器官生物量积累速率,有利于其繁殖生长,这也是其对干旱环境适应的具体体现。根-茎和花-叶生物量α有显著性差异,均不存在共同斜率,根-叶、根-花、叶-茎、花-茎生物量α均无显著性差异,均存在共同斜率,表现出同一植物在不同环境下的趋同适应。
