蔷薇科经济林液流特征及环境因子对其的影响1)
2023-05-23郑家银李绣宏鲁绍伟赵娜徐晓天李少宁
郑家银 李绣宏 鲁绍伟 赵娜 徐晓天 李少宁
(河北农业大学,保定,071000) (北京市农林科学院林业果树研究所)
蒸腾是林木生长发育必不可少的生理代谢过程。蒸腾作用将土壤、植物、大气的水分紧密联系在一起,因此,准确掌握各树种蒸腾特征及其对环境变化的响应规律,对于高效利用水资源具有重要意义[1-2]。干旱缺水是生态建设、农业生产和国民经济发展的主要限制因子之一[3]。北京市被列为世界上最缺水的城市之一[4]。北京地区是蔷薇科经济树种集中栽培区,但受水资源和水利设施的限制与影响,经济林灌溉问题长期以来得不到有效解决,少部分有灌溉条件的经济林区,因人们对经济林需水和耗水规律缺乏了解或对其不合理灌溉,所以限制了经济林生态功能、经济效益的发展[5-6]。提高经济林树种水分利用效率对改善生态环境质量、提高经济收益、缓解北京市用水紧缺现状具有重要意义。
目前,林木蒸腾耗水的研究对象多为园林绿化树种,对于蔷薇科经济林树种的研究较少[7-8]。现阶段研究表明,部分经济林树种液流启动时间基本为日出前后,液流日变化一般呈现出“昼高夜低”的趋势,液流变化曲线多呈“几”字单峰型、双峰型、多峰型等类型。正午时太阳辐射较强,蒸腾消耗的水分大于根系吸收的水分,气孔关闭,导致林木液流变化趋势呈双峰型甚至多峰型[9-10]。受太阳辐射的影响,晴天液流速率均值和峰值均高于阴天和雨天[11]。桑玉强等[12]研究发现,核桃(Juglansregia)液流具有明显的时间变化特征,液流速率日峰值(0.629 L·h-1)出现在16:30,最大液流量出现在5、6月份,2008—2010年生长季日平均液流量分别为4.96、4.75、4.68 L,液流季节变化总体呈夏季液流速率均值最高,春、秋次之,冬季最小的变化趋势。李自豪等[13]研究发现,樟子松(Pinussylvestris)的季节平均树干液流速率由大到小依次为夏季、春季、秋季,树干液流月平均速率在8月份最高(870.97 g·h-1),10月份最小(216.44 g·h-1)。有研究表明,树干液流易受环境因子(太阳辐射、空气温湿度、土壤水势等)的强烈影响,但由于各环境因子不是独立作用于液流,且环境因子与液流的关系有黑箱性,所以液流与环境因子之间存在时滞效应[11,14]。气象因子和土壤因子对林木水分运输影响的部位不同,前者主要通过影响叶片气孔开闭程度改变个体水分蒸发,后者通过改变根系和根际土壤界面间的水分导度从而影响水分吸收[15-16]。
为探明北京市蔷薇科经济林树种液流特征,试验以4种蔷薇科树种为研究对象,利用热扩散探针式茎流仪连续测定各树种液流变化,结合同步监测的环境因子,探究各蔷薇科经济林树种液流与环境因子的相关性及响应规律,为北京地区合理构建经济林配置,解决水资源短缺和经济林树种灌溉管理之间的矛盾提供参考。
1 研究区概况
研究地位于北京市顺义区高丽营镇,地理位置为116°29′41″E,40°11′8″N,属北温带大陆性季风气候,年均气温为11.5 ℃,春季雨量少,夏季炎热多雨,冬季寒冷干燥,年均相对湿度50%,全年降水约80%集中在夏季,近10 a年均降水量约563.65 mm,年日照时间为2 000~2 800 h。2018年—2020年北京市总用水量中超33.2%来自地下水,2020年全市总用水量为40.6亿m3,农业用水3.2亿m3,占总用水量的7.8%。截至2018年,北京市经济林面积超15万hm2,经济林面积约占北京市森林资源面积的30%。研究区各类经济果树多种多样,其中最具代表性的品种有苹果(Malusdomestica)、梨(Pyruspyrifolia)、桃(Amygdaluspersica)、李(Prunussalicina)、杏(Armeniacavulgaris)等。
2 材料与方法
2.1 树种选择
本研究以桃树(瑞光)、李树(沸腾)、杏树(凯特)、梨树(早红考密斯)等北京市常见4种蔷薇科经济林树种为试验材料,在栽植密度为3 m×3 m的样地上进行每木检尺。每个树种分别选择3棵标准样树(生长状况良好、胸径差别不大且无病虫害),应用热扩散式液流测定仪进行连续测定。由于冬季各树种处于休眠状态,试验时间为2021年4月1日—2021年11月3日。

表1 测量液流样树基本情况
2.2 环境因子测定
样地内布设vantage pro 2全自动气象站(Davis Instruments,美国)对环境因子数据进行采集,频率为5 min 1次。监测指标包括:太阳辐射(Rs)、风速(W)、风向、大气温度(Ta)、大气相对湿度(RH)、降水量(P)、土壤温度(Ts)、土壤水势(SWP)等。水汽压亏缺(VPD)由空气相对温度和相对湿度计算得到,公式如下:
VPD=a·eb×Ta/(Ta+c)·(1-RH)。
式中:Ta为空气相对温度(℃);RH为空气相对湿度(%);a、b、c为常数,数值分别为0.611、17.502、240.97。
2.3 树干液流测定
利用热扩散探针式茎流仪(Campbell Scientific,美国)测定树干边材液流密度,将20 mm探针刺入茎内的木质边材,并接通恒定电流以测定边材的导热率,数据采集频率为10 min 1次。通过测量2个探针间的温度差,用Granier经验公式计算树干液流密度,公式如下:
式中:Js为液流密度(cm3·cm-2·s-1);ΔTmax为24 h内上下探针的最大温差值(℃);ΔT为某时刻瞬时温差值(℃),即当时测定的温差值。
2.4 数据处理
将测得数据通过Excel 2013和Sigmaplot 13.0进行整理,再通过SPSS 23.0进行相关性分析、显著性分析、主成分分析。由于液流数据采集频率与环境因子数据釆集频率不同,因此,以1 h测定数据的均值进行分析。
3 结果与分析
3.1 经济林树种液流变化特征
3.1.1 不同季节液流日变化
试验选取各树种生长季内,每个月不连续10 d的晴天下林木液流数据进行日平均(图1)。蔷薇科树种液流昼夜差异明显,白天液流密度较大(2.98~15.04 cm3·cm-2·h-1),夜间液流密度较小(0~3.36 cm3·cm-2·h-1)。液流增强时间段为春季(4月1日—5月4日)的05:00—08:30、夏季(5月5日—7月7日)的05:00—07:00、秋季(7月8日—11月6日)的07:00—09:30;液流减弱时间段则为春季的18:00—21:00、夏季的19:00—23:00、秋季的17:30—20:00。春季,李树的液流密度变化呈双峰型,有晨间冲高回落的趋势,在08:00、12:00左右液流密度达到峰值,分别为14.14、13.64 cm3·cm-2·h-1;杏树、梨树、桃树液流变化趋势呈单峰型,杏树、梨树在09:00左右液流密度达到峰值,分别为15.03、8.01 cm3·cm-2·h-1,桃树在12:00左右达到峰值,为6.48 cm3·cm-2·h-1。夏季,杏树、李树、梨树液流变化趋势呈单峰型,杏树、梨树的峰值出现在11:00,分别为13.74、5.97 cm3·cm-2·h-1,李树的峰值出现在13:00,为14.06 cm3·cm-2·h-1;桃树液流趋势呈双峰型,在07:00、13:00出现峰值,分别为3.46、7.12 cm3·cm-2·h-1。秋季,各树种液流趋势均呈单峰型,杏树、梨树的峰值均在12:00左右出现,分别为5.71、4.74 cm3·cm-2·h-1;李树、桃树的峰值在13:00左右出现,分别为9.02、2.99 cm3·cm-2·h-1。生长季内夜间各树种整体液流密度均为前半夜(1.66 cm3·cm-2·h-1)强于后半夜(0.87 cm3·cm-2·h-1)。

图1 各季节晴天树种液流日变化
3.1.2 不同月份液流日变化
试验将各树种生长季内液流数据进行日平均,同时结合环境因子的变化,其结果见图2。随着月份推移,各树种液流密度整体呈先增加后减少趋势,且液流变化明显受树种生长状况的影响。杏树、梨树液流密度的峰值出现在6月份,分别为14.43、7.44 cm3·cm-2·h-1,李树、桃树的峰值出现在4月份,分别为14.05、6.92 cm3·cm-2·h-1。生长季内各树种整体平均液流密度由大到小依次为:李树(5.64 cm3·cm-2·h-1)、杏树(5.03 cm3·cm-2·h-1)、梨树(3.26 cm3·cm-2·h-1)、桃树(2.43 cm3·cm-2·h-1)。各树种在整个生长季内液流波动幅度最大是李树,为0.18~14.46 cm3·cm-2·h-1,最小的是桃树,为0.32~6.84 cm3·cm-2·h-1。各树种在不同月份内液流密度峰值波动幅度由大到小依次为:桃(0~534.99%)、杏(0~304.85%)、梨(0~160.34%)、李(0~53.00%)。梨树根系生长每年有2个生长高峰,第一次生长高峰出现在新梢停止生长时,第二次高峰出现在9—10月,在适宜条件下,梨树根系可周年生长,无休眠期[17]。李树、梨树由于树种自身生理特性,其落叶期晚于其他树种,表现为秋季液流密度远大于其他树种,其中,梨树10月份液流强度大于9月份。各树种的液流密度变化种间差异明显,即使同为蔷薇科树种,液流曲线相似,但液流峰值、启动时间也有明显差异。
3.2 环境因子对经济林树种液流的影响
3.2.1 气象因子变化对经济林树种液流的影响
选取2020年气象因子变化最剧烈的夏季和秋季,将各树种液流数据平均成24 h液流图,同步结合环境因子,得到图3。由图2、图3、表2可知,在整个生长季内,各树种树干液流均受到VPD、Rs、Ta、RH等微气象因子的极显著影响。梨树和桃树的树干液流与P呈显著负相关,各树种树干液流与VPD呈极显著正相关。夏季,除梨树外各树种液流峰值整体大致晚于VPD峰值(2.25 kPa);秋季,各树种液流峰值均早于VPD峰值(2.10 kPa)。各树种液流与Rs呈极显著正相关,各树种液流峰值基本早于Rs峰值(夏季710.96 W·m-2,秋季371.43 W·m-2),但夏季,杏树、桃树液流峰值晚于Rs峰值。各树种液流与Ta呈极显著正相关,各树种液流峰值均早于Ta峰值(夏季30.35 ℃,秋季18.99 ℃)。各树种液流密度随着Ta增加而增大,但当Ta大于各树种的最适温度时,各树种液流密度随着Ta增加而减少。林木液流与RH呈极显著负相关,各树种液流密度随着RH的减少而增大,各树种液流峰值均早于RH最小值(夏季46.95%,秋季34.30%)。生长季内,梨树、桃树的树干液流与P呈显著负相关,其液流峰值均早于P的最大值(夏季0.19 mm,秋季0.06 mm)。各树种间虽然存在差异,但对各气象因子的响应却始终保持相似性。各树种生长发育对水分的需求十分巨大,但是P的改变却并没有引起液流的实时变化,表明各树种树干液流对气象因子的响应存在时滞性。
3.2.2土壤温度、土壤水势变化对经济林树种液流的影响
试验选取2020年4月至10月各树种液流数据进行平均,其结果见图4。6月到7月,SWP急剧下降(73.58~2.36 kPa),之后一直保持在较低水平(0.86~3.58 kPa)。同时,Ts受SWP的强烈影响,自8月后下降较大(23.79~14.68 ℃),出现该现象的原因是2021年北京地区出现了大雨季,雨季从6月底开始,于7月中下旬达到峰值,雨量远超以往,延续到9月中旬减弱。由图4、表1可知,各树种树干液流在生长季内与SWP呈极显著正相关;梨树树干液流在生长季内与Ts呈显著负相关(R=-0.187),桃树树干液流与Ts呈极显著负相关(R=-0.458)。小尺度上,SWP波动较小,只有在降雨等特殊时期,SWP短时间内波动巨大。但由于土壤的理化性质,同时基于地膜影响,短时间内的降雨只有部分可以渗透到地表之下,大部分被排出林外。短期内SWP增高可增强各树种的蒸腾作用,促进林木液流昼夜波动加大。大尺度上,由于降雨量大,易导致林区内排水渠道堵塞,使大量雨水汇集在林区内,田间持水量饱和,形成涝灾,各树种生存环境恶劣,液流强度降低。
各树种液流密度随Ts的增加而减少,随SWP增加而增大。理论上,各树种的液流峰值应该出现在Ts最低的10月和SWP最高的6月,然而各树种液流峰值却集中在6月或7月。大雨季的出现,致使短时间内P过大,严重影响了VPD、Rs、RH、Ta,导致Ts、SWP出现特殊变化,从而打破各树种树干液流变化的整体趋势。这一现象进一步体现了影响各树种树干液流变化的不是单一环境因子作用,而是多因子的共同作用。
3.2.3 各环境因子与经济林树种液流的相关性
各经济林树种液流变化易受环境因子(微气象因子、土壤因子)的影响。在日尺度上将生长季内各树种液流与环境因子进行相关性分析,其结果见表2。各树种液流与VPD、Rs、Ta、SWP均呈极显著正相关,与RH呈极显著负相关。梨树液流与P、Ts呈显著负相关,相关系数分别为-0.14 5、-0.187;桃树液流与Ts呈极显著负相关(R=-0.458,P<0.01)。由此可知,不同树种对各环境因子的响应程度不一,在具体实践管理中需结合不同树种的生长习性及管理目的,在大时间尺度上把握各经济林的生产方向,小尺度精细调控灌溉方式,从而达到提高生产水平和节约用水的目的。
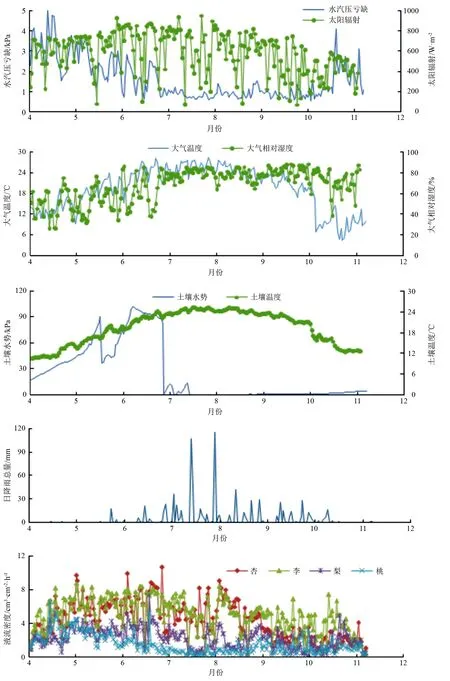
图2 各树种液流及环境因子月变化
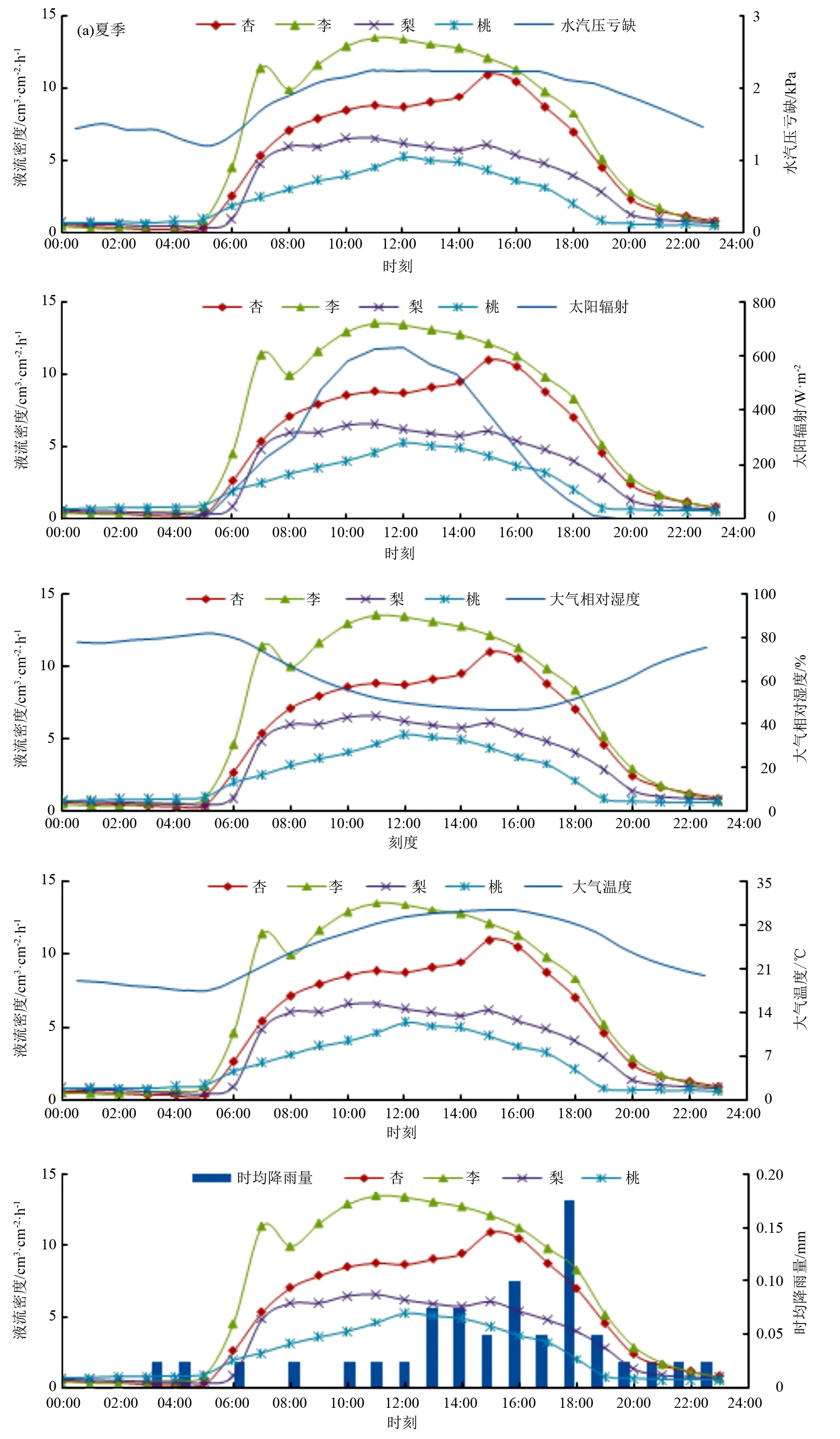
图3 气象因子与不同树种液流的关系曲线
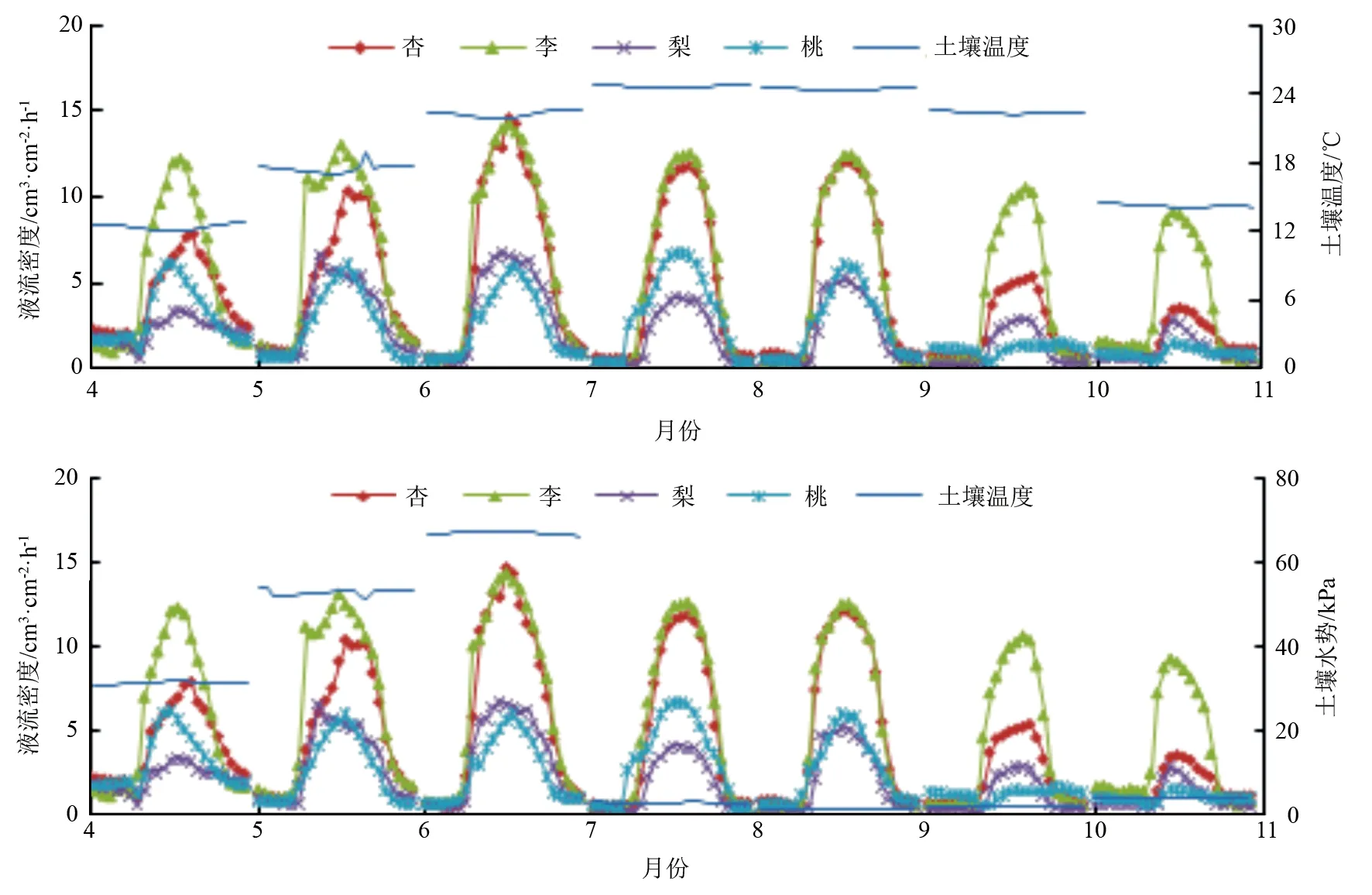
图4 土壤温度、土壤水势与不同树种液流的关系曲线
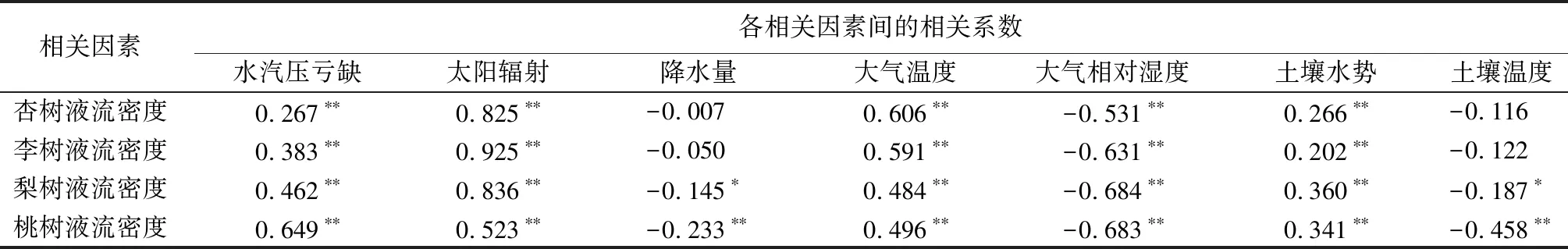
表2 环境因子和各树种液流密度相关性
3.2.4 树种液流变化对环境因子的时滞性
为探索各树种树干液流对不同环境因子响应的时滞效应,采用错位分析法,即将各树种树干液流数据与提前或延后3小时的环境因子数据进行相关性分析,结果见表3。杏树、李树、梨树树干液流密度与1 h前VPD的相关性最强,相关系数分别为0.286、0.421、0.478(P<0.01);桃树树干液流与2 h前VPD的相关性最强,相关系数为0.682(P<0.01)。杏树、李树、梨树树干液流密度均对1 h后Rs的响应程度最高,相关系数分别为0.853、0.945、0.863(P<0.01);桃树树干液流与实时Rs的相关性最强,相关系数为0.523(P<0.01)。杏树、李树、梨树、桃树树干液流密度对3 h后P的响应程度最高,相关系数分别为-0.164、0.179、-0.251、-0.265。杏树、李树、梨树树干液流密度对1 h前Ta的响应程度最高,相关系数分别为0.634、0.633、0.514(P<0.01);桃树树干液流密度与实时Ta的相关性最大,相关系数为0.496(P<0.01)。杏树、李树、梨树树干液流密度对1 h前RH的响应程度最高,相关系数分别为-0.562、-0.686、-0.716(P<0.01);桃树树干液流对2 h前RH的响应程度最高,相关系数为-0.743(P<0.01)。杏树、李树、梨树、桃树树干液流密度与3 h前SWP的相关性最大,相关系数分别为0.277、0.207、0.373、0.352(P<0.01)。梨树树干液流密度与实时Ts的相关性最大,相关系数为-0.187(P<0.05);桃树树干液流密度对3 h前Ts的响应程度最高,相关系数为-0.471(P<0.01)。由上可知,试验中生长季内各树种液流变化对VPD、Rs、Ta、RH等环境因子均有不同程度的时滞性,且由于天气、月份、季节、物候期的不同,同一树种对同一环境因子的时滞程度也不同。
3.2.5 主成分分析与回归模型
将环境因子对各树种液流变化的影响进行主成分分析,其结果如表4。提取3个主成分,即k=3,3个主成分方差累计贡献率分别为49.348%、76.674%、87.119%,大于85%。所提取主要因子的得分越高则说明各树种液流变化对其响应程度越高。因此VPD、Ta、RH3个主成分能够反映各环境因子的大部分信息,解释环境对各树种液流变化的影响程度,决定各树种液流变化的趋势。
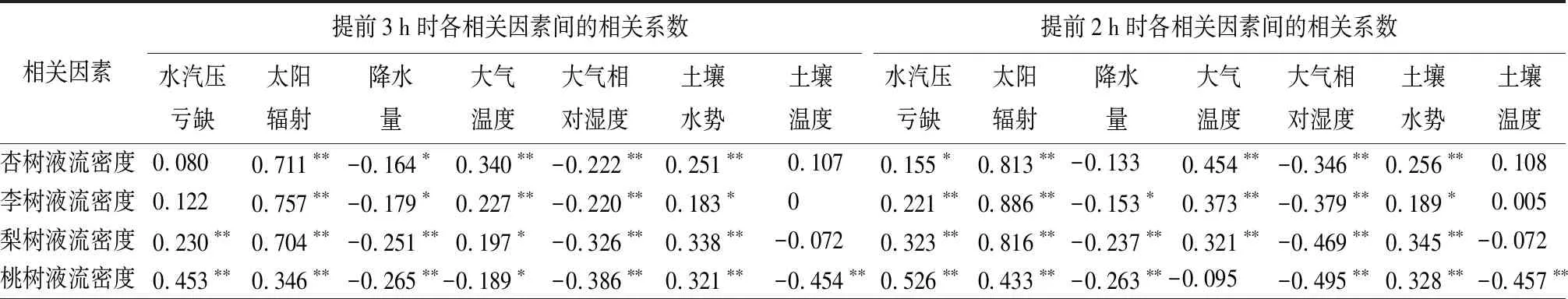
表3 环境因子和各树种液流密度错位相关性

表4 环境因子对各树种液流变化影响的主成分分析结果
将生长季内各树种树干液流(y)与环境因子进行回归分析,拟合多元回归模型,其结果见表5。

表5 各树种液流密度与环境因子的回归分析结果
结合表2、表3、表4可知,各环境因子对杏树树干液流的影响由大到小依次为:Rs、Ta、SWP,对李树树干液流变化影响最大的是Rs,对梨树树干液流变化影响最大的是Rs、SWP,对桃树树干液流变化影响最大的是Rs、VPD。表3与表4所反映的环境因子有所差异,原因是表3主要为大环境中对整个经济林产生重要影响的环境因子,而表4体现了不同树种对各环境因子的主要响应程度,二者并不冲突。为了避免数据过多造成散点图杂乱,现将观测期内每月数据平均成1 d数据,对各树种液流与对其影响较大的环境因子进行拟合(图5),各散点图和拟合方程反映了单个环境因子与液流的整体趋势,也从侧面验证了回归方程精确度。
4 讨论
杏、李、梨、桃等蔷薇科经济林树种液流变化具有明显的昼夜差异,晴天液流变化大体呈单峰型和双峰型,在12:00—13:00左右出现峰值波动(0.000 9~0.004 2 cm3·cm-2·h-1)。树种不同,液流变化趋势、峰值、启动时间皆不相同。周玉燕等[18]发现,山旱塬区苹果树干液流有明显的时间变化特征,晴天峰值在15:00左右,阴天在12:00、17:00左右。随着月份推移,各树种液流密度逐渐增加,在6、7月份达到峰值后缓慢减少,直至树种休眠期,液流活动趋近0,保持在树种最低生存范围,且液流变化明显受到树种生长状况影响。桑玉强等[12]研究发现,核桃液流的最大液流量出现在5、6月份,其次为7、8、9月份。各经济林树种液流变化在季节上的整体表现为夏季大于春季大于秋季。各树种在春季叶片、花芽、坐果等生长发育迅速,整体呈现液流速率增加,液流活动不断加强;夏季,叶片发育成熟,树木处于果实膨大期,外界持续高温,林木为维持蒸腾和繁殖,液流活动强烈,并保持在较高强度的波动范围;秋季,果实成熟,叶片衰老和脱落,液流减弱。马长明等[19]发现,核桃5月进入果实膨大期,生殖及营养生长均旺盛,液流速率进一步提升;8月进入果实成熟期,外界环境持续高温,树干液流速率达到峰值。对果树生育期不同阶段的水分利用研究可为人类社会活动及生产提供服务,且对经济林树种的栽培管理、提高经济及生态效益具有重要作用[20]。本研究对各树种液流变化特征的探究受地域、品种、种植养护管理的局限,未来还需打破这些局限,进一步深入研究。

图5 树种液流与重要环境因子散点图
林木液流受环境影响,具有较大复杂性,不同的地区、水热条件、气候对林木液流的影响均不相同[21]。本试验中,各蔷薇科树种液流受VPD、Rs、P、RH、Ta、SWP、Ts等环境因子的显著影响,其与VPD、Rs、Ta、SWP呈极显著正相关,与RH呈极显著负相关,且液流密度的变化与气象因子之间存在时滞性。王华等[22]发现,林木液流与VPD、Rs等呈正相关,与RH呈负相关;万发等[23]发现,引黄灌区苹果树白天液流的主要驱动因子为VPD、Rs、Ta等环境因子,而液流变化与气象因子间存在时滞性。各树种对不同环境因子的响应程度不一致,杏树液流对各环境因子响应程度由大到小依次为:Rs、Ta、SWP;桃树液流对各环境因子响应程度则是Rs大于VPD。该现象说明,影响林木液流的主要气象因子因树种差异而存在差别。Tognetti et al.[16]提出,土壤水分亏缺会增加土壤和根系等阻力,从而影响土壤到林木叶片的水分运输,造成气孔关闭,最终导致耗水减少。Cochard et al.[24]研究表明,当土壤温度改变时,较高的土壤温度更有利于植物的蒸腾耗水。本研究中,部分树种液流受到Ts、SWP显著或极显著影响,其中SWP对树种液流的影响程度相较于其他环境因子更大,这一结果与前人研究中环境因子对树种液流影响程度的排名有所差异[25-26]。原因是大雨季的出现严重促使了SWP改变,特大降雨量和积水导致SWP在7月后大部分时间段内接近0,此时各树种根系处于饱和田间持水量的土壤中。由于SWP对各树种液流的影响相较于其他环境因子的影响程度更大,因此打破了气候因子对各树种树干液流变化的整体影响趋势。有研究表明,林木体内会由于空穴化作用形成栓塞,导致导管中水柱中断,降低导管的输水能力,严重影响林木液流活动,阻碍林木生长[27-29]。因此,下一步应加强对于各树种内部因子的研究。
5 结论
杏树、李树、梨树、桃树等蔷薇科经济林树种液流昼高夜低,差异明显,晴天各树种液流变化呈单峰型和双峰型2种类型。液流密度随月份推移逐渐增加,6、7月份达到峰值,之后缓慢减少,直至树种休眠期,液流密度趋近0。生长季内各树种耗水强度由大到小依次为:李树、杏树、桃树、梨树,且液流变化明显受到树种生长状况影响。
蔷薇科树种液流受环境因子影响强烈,各树种种间虽然存在差异,但对各气象因子响应却始终保持相似性。树种液流密度的变化与气象因子之间存在时滞性,时滞程度受天气、季节、物候等因素的影响。各树种液流变化虽然受到多因子共同作用,但当某一环境因子的影响程度过大时,会打破各环境因子共同作用于液流变化的平衡,形成“独大”的局面。
