蜡样芽孢杆菌NJSZ-13对松材线虫生理生化指标的影响1)
2023-05-23潘敏许嘉麟谈家金孙玉凤叶建仁
潘敏 许嘉麟 谈家金 孙玉凤 叶建仁
(南京林业大学,南京,210037)
松材线虫病是由松材线虫(Bursaphelenchusxylophilus)引起的一种可危害黑松(PinusthunbergiiParlatore)、马尾松(PinusmassonianaLamb.)、火炬松(PinustaedaL.)、黄松(PodocarpusimbricatusBl. Enum. Pl. Jav.)等多种松树的病害[1-3]。松材线虫主要由松墨天牛(Monochamusalternatus)、云杉花墨天牛(M.saltuarius)等为媒介昆虫进行远距离传播,在感病30~40 d后,寄主松树枯死[4]。松材线虫病在我国首次被发现是在1982年,自此,该病害侵害范围不断扩大,已经蔓延至我国多个省市地区[5]。由于松材线虫病具有快速传播性和高度危害性,导致其对全世界多种松树资源都存在着巨大威胁,该病的防控已成为世界性的难题[6]。
近年来,生物防治逐渐进入大众视野,其最突出的特点就是不污染环境,与化学防治相比,具有较强的环境友好性[7-8]。微生物是生物防治的重要组成部分,而细菌因易于繁殖、生产简单等优点,在生物防治中被广泛应用[9-11],其中又以芽孢杆菌菌剂应用较为广泛[12-14]。Li et al.[11]从马尾松中分离得到短小芽孢杆菌(Bacilluspumilus)LYMC-3,其代谢产物中的胍类化合物能对松材线虫产生明显抑杀作用。徐华潮等[15]研究发现2株苏云金芽孢杆菌(Bacillusthuringiensis)020和RBT-200701的胞外毒素对松材线虫表现出较强的杀线虫活性。
氧自由基的化学反应活性高,极不稳定,易与其他物质发生化学反应,直接或间接造成细胞结构的破坏,从而导致细胞凋亡[16]。而氧自由基能被松材线虫体内的过氧化物酶(POD)、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)清除,减少氧自由基对线虫的危害[17-18]。谷胱甘肽过氧化物酶(GSH-Px)能够催化谷胱甘肽(GSH)与活性氧反应,减轻活性氧对线虫的损害[19]。丙二醛(MDA)是常见的膜脂过氧化指标,MDA的质量摩尔浓度与线虫氧化损伤程度呈正相关[20-22]。因此,检测线虫体内MDA的质量摩尔浓度及SOD、POD、CAT、GSH-Px的活性可以反映机体被氧化损伤的程度。一般来说,乙酰胆碱酯酶(AchE)在突触或神经肌肉连接处,催化神经递质乙酰胆碱(ACh)的水解,同时终止神经冲动,是松材线虫不可或缺的酶[23-24]。
蜡样芽孢杆菌(Bacilluscereus)NJSZ-13是从松树体内分离得到的一株生防菌,已知松材线虫在NJSZ-13的菌悬液中处理48 h后,死亡率达到81.5%[25]。为了进一步明确菌株NJSZ-13的杀线机理,本研究在此基础上,探究NJSZ-13菌株代谢产物对松材线虫体内可溶性蛋白质质量浓度、MDA质量摩尔浓度、AchE和保护性酶(SOD、POD、CAT、GSH-Px)活性的影响,为研发微生物源杀线剂提供参考。
1 材料与方法
供试材料:蜡样芽孢杆菌NJSZ-13菌株和松材线虫AMA3虫株来源于南京林业大学森林病理实验室。
营养琼脂(NA)固体培养基:牛肉膏5 g/L,氯化钠5 g/L,蛋白胨10 g/L,琼脂20 g/L,pH为7.2~7.4。
营养肉汤(NB)液体培养基:牛肉膏5 g/L,氯化钠5 g/L,蛋白胨10 g/L,pH为7.2~7.4。
菌株NJSZ-13菌悬液和发酵滤液的获得:挑取单个NJSZ-13菌落接入NB液体培养基中,在28 ℃、200 r/min条件下摇培4 d,即得发酵液。将发酵液于4 ℃、10 000 r/min条件下离心15 min后,上清液用注射器经0.22 μm微孔滤膜过滤,即得发酵滤液,沉淀菌体用等体积的无菌水悬浮即得菌悬液(菌体浓度为3×108个/mL)。
菌株NJSZ-13处理松材线虫:采用浸渍法,将一定数量的松材线虫(每个处理30 000条)用蜡样芽胞杆菌NJSZ-13的发酵液、发酵滤液、菌悬液分别处理6、12、24、36 h后(对照用无菌水和NB液体培养基处理),于室温条件下,以3 500 r/min离心5 min去除浸渍液,之后用无菌水冲洗沉淀3次,每个处理重复3次。
线虫生理生化指标测定:将供试线虫液与可溶性蛋白提取液混合于1.5 mL离心管中,用低温预冷组织研磨仪(净信JXFSTPRP-CL)将其研磨180 s,4 ℃、3 500 r/min条件下离心10 min,上清液即为组织匀浆液。线虫可溶性蛋白质质量浓度测定采用Bradford法[26],利用紫外分光光度计(Perkin Elmer Lambda 365)在波长595 nm处测量吸光值。其余6种指标(SOD、POD、CAT、GSH-Px的活性、MDA质量摩尔浓度和AchE活性)的测定按照苏州科铭生物技术有限公司的试剂盒方法进行。
数据处理:采用Excel 2021进行数据整理,IBM SPSS Statistics 25软件进行数据统计和分析。
2 结果与分析
2.1 菌株NJSZ-13对松材线虫可溶性蛋白质质量浓度的影响
菌株NJSZ-13在不同处理时间对松材线虫体内蛋白质质量浓度的影响如表1所示,对照组与处理组在整个处理时间均呈下降趋势。前6 h,处理组松材线虫体内蛋白的质量浓度缓慢下降,处理组与对照组差异显著(p<0.05);6 h后,处理组松材线虫体内蛋白的质量浓度急剧下降,发酵液和发酵滤液处理组尤为明显,且极显著地低于对照组(p<0.05)。由数据可知,3组处理组中,线虫体内蛋白质质量浓度与对照组相比显著下降(p<0.05),这是造成菌株NJSZ-13杀死松材线虫的重要原因之一。
2.2 菌株NJSZ-13对松材线虫体内超氧化物歧化酶活性的影响
菌株NJSZ-13在不同处理时间对松材线虫体内超氧化物歧化酶(SOD)活性的影响如表2所示,在整个处理时间内,SOD活性呈现出先升高后降低的趋势。在0~12 h内,处理组松材线虫体内的超氧化物歧化酶活性均高于对照组。6 h时3组处理组高于对照组,差异显著(p<0.05);在12 h时各处理组和对照组结果相近,无显著性;在24、36 h时,3组处理组低于2组对照组,差异显著(p<0.05)。
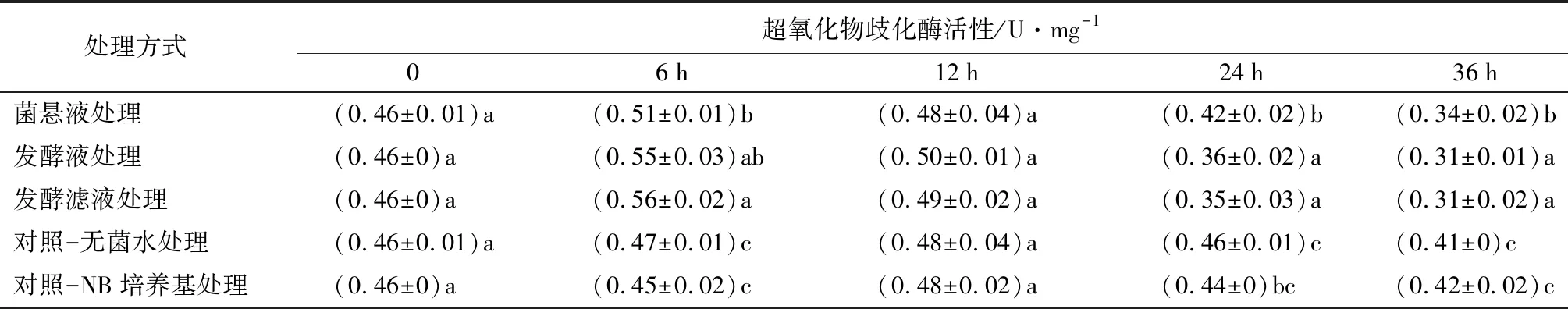
表2 不同处理的松材线虫超氧化物歧化酶活性
2.3 菌株NJSZ-13对松材线虫过氧化物酶活性的影响
菌株NJSZ-13在不同处理时间对松材线虫体内过氧化物酶(POD)活性的影响如表3所示,菌株NJSZ-13处理后,松材线虫体内的过氧化物酶活性在整个处理时间段均低于对照组。0~12 h时,菌悬液处理与对照组无显著差别,24、36 h时显著降低(p<0.05)。发酵液和发酵滤液处理后,与对照组相比,POD活性在处理后6、12 h存在显著差异,在24、36 h时有极显著差异(p<0.05)。
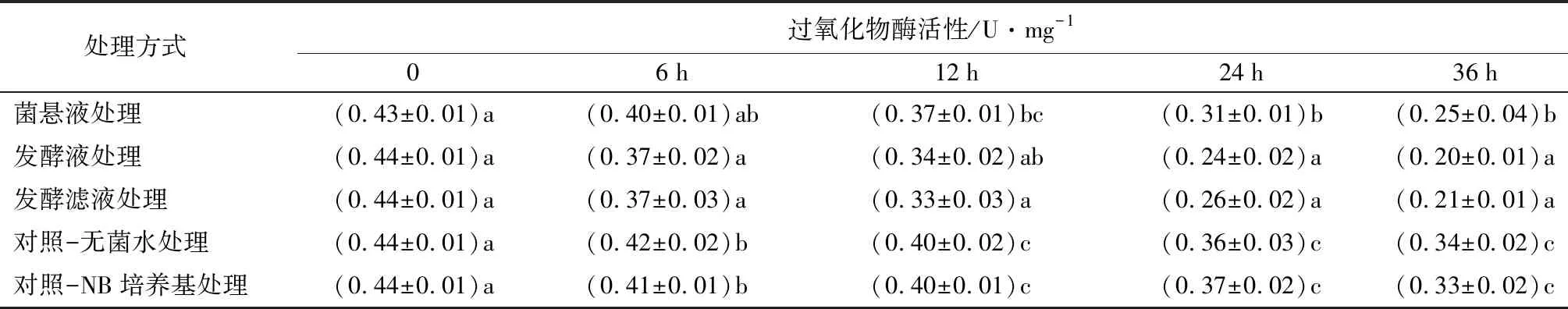
表3 不同处理的松材线虫过氧化物酶活性
2.4 菌株NJSZ-13对松材线虫过氧化氢酶活性的影响
菌株NJSZ-13在不同时间对松材线虫体内过氧化氢酶(CAT)活性的影响如表4所示,菌株NJSZ-13处理后,松材线虫体内的过氧化氢酶活性的变化趋势与POD相近,在处理6 h时,处理组与对照组有显著差异(p<0.05);6 h后,CAT活性急剧下降,与对照组形成显著差异(p<0.05)。

表4 不同处理的松材线虫过氧化氢酶活性
2.5 菌株NJSZ-13对松材线虫谷胱甘肽酶活性的影响
菌株NJSZ-13在不同处理时间对松材线虫体内谷胱甘肽酶(GSH-Px)活性的影响如表5所示,菌株NJSZ-13处理后,松材线虫体内谷胱甘肽酶活性表现出先升高后降低的趋势。菌悬液处理后,谷胱甘肽酶活性在6、12 h时,与对照差异不显著,24、36 h时则显著低于对照(p<0.05)。发酵液和发酵滤液处理后,谷胱甘肽酶活性在6 h时高于对照,但差异不显著,随后急剧下降,与对照组形成显著差异(p<0.05)。
2.6 菌株NJSZ-13对松材线虫丙二醛质量摩尔浓度的影响
菌株NJSZ-13在不同处理时间对松材线虫体内丙二醛(MDA)质量摩尔浓度的影响如表6所示,松材线虫体内丙二醛质量摩尔浓度在菌株NJSZ-13处理后的变化区别于其他几种物质,呈现出上升趋势。菌悬液处理后的松材线虫,体内MDA质量摩尔浓度在6、12、36 h时较对照略高,无显著差别,24 h时显著高于对照(p<0.05)。发酵液和发酵滤液处理后的松材线虫体内的MDA质量摩尔浓度在6、24 h时与对照有显著差异(p<0.05),在12 h与对照组均达到极显著差异水平(p<0.05),在36 h时发酵液处理与对照差异不显著,发酵滤液处理与对照极显著(p<0.05)。

表5 不同处理的松材线虫谷胱甘肽酶活性
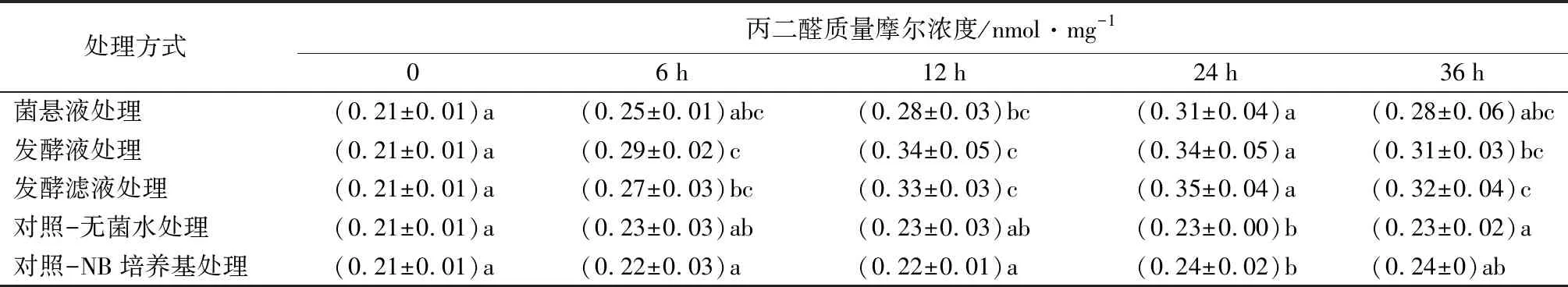
表6 不同处理的松材线虫丙二醛质量摩尔浓度
2.7 菌株NJSZ-13对松材线虫乙酰胆碱酯酶活性的影响
菌株NJSZ-13在不同处理时间对松材线虫体内乙酰胆碱酯酶(AchE)活性的影响如表7所示,其中对照组和3组处理组的线虫体内,AchE活性变化大体一致,均呈下降趋势。处理6 h时,菌悬液处理与对照无显著差异,发酵液和发酵滤液处理较对照显著降低(p<0.05);处理12 h时,菌悬液处理酶活性降低,与对照组差异不显著,发酵液和发酵滤液较对照极显著降低(p<0.05);处理24、36 h时,菌悬液酶活性显著降低,发酵液和发酵滤液均极显著降低(p<0.05)。
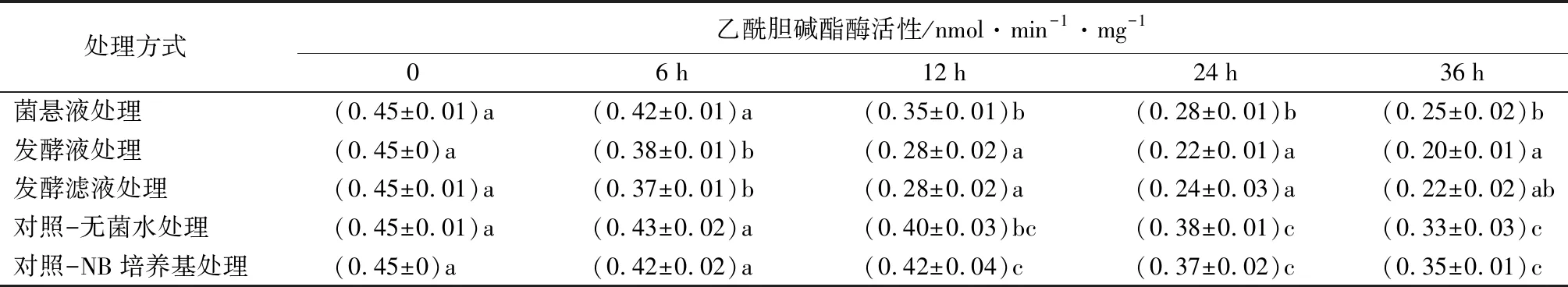
表7 不同处理的松材线虫乙酰胆碱酯酶活性
3 讨论与结论
经菌株NJSZ-13处理后,松材线虫SOD和GSH-Px的活性呈现先升高后降低的趋势。2种物质的活性在处理前期升高,表明细菌处理导致线虫启动防御机制,引起活性氧迸发,产生大量超氧自由基和过氧化物,机体以提高体内SOD和GSH-Px活性开启自身免疫反应。此后2种酶的活性下降,使线虫体内的氧自由基不能及时被清除,造成其抗氧化保护酶系统被破坏,最终导致线虫死亡。随着细菌处理时间的延长,线虫另外2种保护性酶POD和CAT活性受到抑制,进一步导致超氧自由基和过氧化物不断积累。蒋春艳[27]研究发现SOD、POD、CAT 3种保护酶在黄曲条跳甲抵抗绿僵菌引起的氧化胁迫中发挥了重要作用。李会平等[28]发现在桑天牛对白僵菌的免疫作用中,SOD和POD是首先发生作用的酶系,CAT是继上述两种酶之后发生作用的酶系,其中POD所起作用更为重要。经菌株NJSZ-13处理后,线虫体内MDA质量摩尔浓度不断上升,说明了线虫脂质过氧化程度不断加重,其细胞结构和功能持续受损,可能最终导致线虫的死亡。
线虫体内的大多数可溶性蛋白质为参与各种代谢的酶类,所以可溶性蛋白质质量浓度是一个重要的生理生化指标。3组处理的松材线虫体内,可溶性蛋白质质量浓度显著低于对照,表明菌株NJSZ-13使松材线虫体内蛋白代谢受到严重影响。AchE是生物神经传导中的关键性酶,在神经传导调节中起重要作用[29]。试验中发现,菌株NJSZ-13处理后,松材线虫行动迟缓,反应迟钝,一段时间后虫体僵直死亡。这与其体内的AchE活性降低,导致神经递质乙酰胆碱(Ach)无法正常水解,造成线虫神经传导受到影响有关。侯金丽等[30]发现,松材线虫体内神经系统关键酶(TChE)的活性受到Sr18菌代谢产物的显著抑制,造成线虫体内乙酰胆碱的大量积累,使突触后膜长期处于兴奋状态,导致神经传递中断,引起线虫的中毒反应。同时,翁群芳等[31]研究发现,骆驼蓬提取物能抑制松材线虫体内乙酰胆碱酯酶活性,从而导致线虫死亡。
在蜡样芽孢杆菌NJSZ-13处理松材线虫的过程中可以发现,发酵液对松材线虫的各项生理指标影响最为明显,这与前期试验中发酵液对线虫的致死率最高相一致。通过菌株处理后线虫的各个生理效应可以推测,菌株NJSZ-13对松材线虫的致死作用是综合性的,其中包括蛋白代谢、保护酶系统和神经系统等方面的共同作用。在今后的研究中,可考虑从代谢组和转录组的角度,研究菌株处理后松材线虫的重要代谢通路和关键酶基因的表达量变化,以期找出菌株NJSZ-13对松材线虫的关键致死机制。
