黄海冬季大型底栖动物的群落结构特征❋
2023-05-22滕安康张蒙生杨湘君崔雯瑶邵聪聪于子山
滕安康, 张蒙生, 杨湘君, 崔雯瑶, 陈 晨, 祝 琳, 邵聪聪, 于子山❋❋
(1. 中国海洋大学海洋生命学院, 山东 青岛 266003; 2. 上海勘测设计研究院有限公司, 上海 200050; 3. 山东省海洋科学研究院, 山东 青岛 266100; 4. 海昌(中国)投资有限公司, 上海 200050)
根据分选筛网孔径的大小,海洋底栖动物可以分成大型、小型和微型[1-3]。大型底栖动物是指不能通过孔径为0.5 mm网筛的底栖动物[2-3],其对海洋底层生态系统的物质循环和能量流动过程有重要的作用。对大型底栖动物的研究,有助于加深人类对海洋底层生态系统的了解[3]。
黄海是大陆架上的一个半封闭浅海,面积约38万km2,平均深度约44 m,蕴藏着丰富的海洋生物资源。黄海是中国海洋生态系统最重要的海域之一,也是中国开展大型底栖动物生态学研究最早、最多、最深入的海域[3]。早在20世纪30年代,张玺等[4-7]调查了山东半岛附近海域的大型底栖动物,并先后发表了4期采集报告。20世纪50—70年代,研究主要集中于积累本底资料并开展分类学工作,从而了解黄海大型底栖动物区系的基本特点[8-9]。1980年代至今,依托于973项目、908专项等大型海洋综合项目的开展[3],对黄海大型底栖动物研究所涉及的方面更加广泛,如生物多样性[9-14]、群落结构及其优势种的变化[15-24]、次级生产力[25-27]和功能群[28-29]等。
有关黄海大型底栖动物群落结构方面的研究,最早见于刘瑞玉[8]的报道,其划分群落的方法属于优势种学派。随着计算机技术的发展,20世纪70年代后的群落研究基本采用统计学派的多元统计分析方法[30],研究海域也从近岸扩展到远海。目前已发表的基于多元统计方法划分群落的文献中,对黄海大型底栖动物群落结构的研究所涉及的区域多是黄海局部海域,如:山东半岛沿岸[16-18]、北黄海[19-20]和南黄海[21-22],尚未见有对整个黄海海域进行的报道。本文通过分析在整个黄海海域设置的22个站位的调查资料,探究了黄海冬季大型底栖动物群落结构特征及其变化,并分析了影响群落结构的环境因子,以期为黄海大型底栖动物生态学的深入研究提供参考。
1 材料和方法
1.1 调查海域
于2016年1月(冬季)对黄海海域22个站位的大型底栖动物进行调查取样,调查船为“东方红2号”,站位分布如图1所示。
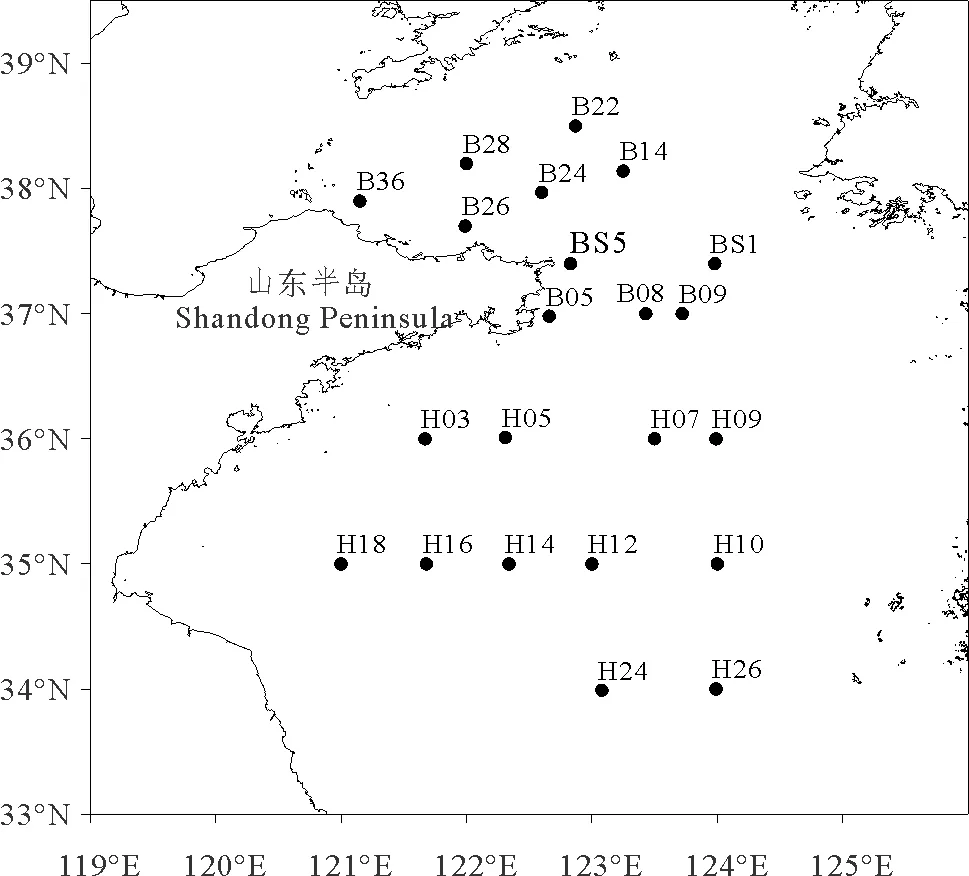
图1 黄海海域大型底栖动物采样站位
1.2 样品采集与处理
使用采样面积为0.1 m2的箱式采泥器,每站取样2次,并合为一个样品,现场用涡旋器冲洗泥样,并过0.5 mm孔径的网筛,将留在网筛上的样品和残渣收集装瓶,用浓度为10%的福尔马林溶液固定。室内样品的处理、鉴定、计数和称量等均按照《海洋调查规范 第6部分: 海洋生物调查》[31]进行。
1.3 环境因子的测定
现场取沉积物表层样品,-20 ℃保存,用于沉积物粒度、叶绿素a(Chla)含量、脱镁叶绿素a(Pha)含量、有机质含量和含水率的测定。其中:粒度使用Mastersizer 2000型激光粒度分析仪测定;叶绿素a含量和脱镁叶绿素a含量采用荧光光度法测定;有机质含量使用重铬酸钾-硫酸氧化法测定;含水率测定参照《海洋调查规范》的方法;水深、底温和底盐数据由CTD测定仪随船现场测得。
1.4 数据处理与分析
1.4.1 优势种 采用相对重要性指数(Index of Relative Importance,IRI) 作为评定大型底栖动物优势种的标准,公式如下:
IRI=(W+N)×F×104。
(1)
式中:IRI为相对重要性指数;W为每种的生物量占总生物量的百分比;N为每种的丰度占总丰度的百分比;F为该种的出现频率。
1.4.2 多样性指数 本文采用了Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(d)以及Pielou均匀度指数(J),公式如下:
(2)
(3)
(4)
式中:S为大型底栖动物的总种数;N为总丰度;ni为第i种的丰度。
1.4.3 群落分析 使用PRIMER 6.0软件对大型底栖动物丰度数据进行CLUSTR分析(划分群落)、ANOSIM分析(群落间差异显著性检验)以及SIMPER分析(计算群落组内相似性)[30]。使用SPSS 17.0软件对生物数据和环境因子进行Pearson相关性分析。
2 结果
2.1 沉积环境
本研究各站位环境数据见表1。调查海域各站位的平均水深为54.18 m,分布趋势为西浅东深、近岸浅远岸深;底温平均值为7.36 ℃,分布趋势为北低南高;底盐平均值为32.26,分布趋势为近岸低远岸高。
调查海域沉积物中Chla的高值站位分布于北黄海,均高于0.80 μg/g;南黄海各站位Chla的范围为0.08~0.91 μg/g,分布不均匀。Pha最高值和最低值的站位均位于南黄海,分布不均匀。有机质和含水率的分布趋势一致,为北低南高。中值粒径的高值站位分布于北黄海;南黄海除近岸H03站的中值粒径高达0.051 mm外,其余各站均低于0.013 mm。北黄海沉积物类型以砂质粉砂为主;南黄海沉积物类型以黏土质粉砂为主。
2.2 大型底栖动物的种类组成与优势种
本研究共发现大型底栖动物202种,隶属于11个门类,其中多毛类104种,占总种数的51.4%;甲壳类48种,占总种数的23.7%;软体动物27种,占总种数的13.4%;棘皮动物8种,占总种数的4.0%;腔肠动物5种,占总种类数的2.5%;尾索动物4种,占总种类数的2.0%;星虫动物2种,占总种类数的1.0%;纽形动物、毛颚动物、半索动物和鱼类各为1种,分别占总种数的0.5%。多毛类、甲壳类和软体动物是该海域冬季大型底栖动物的主要类群。
表2列出了调查海域IRI排名前10位的种,其中IRI大于1 000的优势种为中蚓虫(Mediomastuscaliforniensis)、薄壳索足蛤(Thyasiratokunagai)和浅水萨氏真蛇尾(Ophiurasarsiivadicola)。

表2 调查海域中相对重要性指数排名前十的物种
2.3 大型底栖动物的丰度和生物量及其分布
如表3所示,调查海域冬季大型底栖动物的总平均丰度为1 313 ind/m2,丰度最高值出现在山东半岛东北海域的B28站位,为3 875 ind/m2,该站出现了丰度很高的M.californiensis(2 135 ind/m2);丰度最低值出现在山东半岛东南海域的H05站位,为245 ind/m2。丰度的高值区位于北黄海海域(见图2)。
调查海域冬季大型底栖动物的总平均生物量为25.527 9 g/m2,生物量最高值出现在山东半岛东端近岸的BS5站位,为70.593 5 g/m2,该站采集到1个砂海星(Luidiaquinaria),其生物量达44.808 5 g/m2,存在一定的偶然性;生物量最低值出现在南黄海中部的H26站位,仅为4.374 5 g/m2。生物量的高值区位于黄海冷水团海域(见图2)。
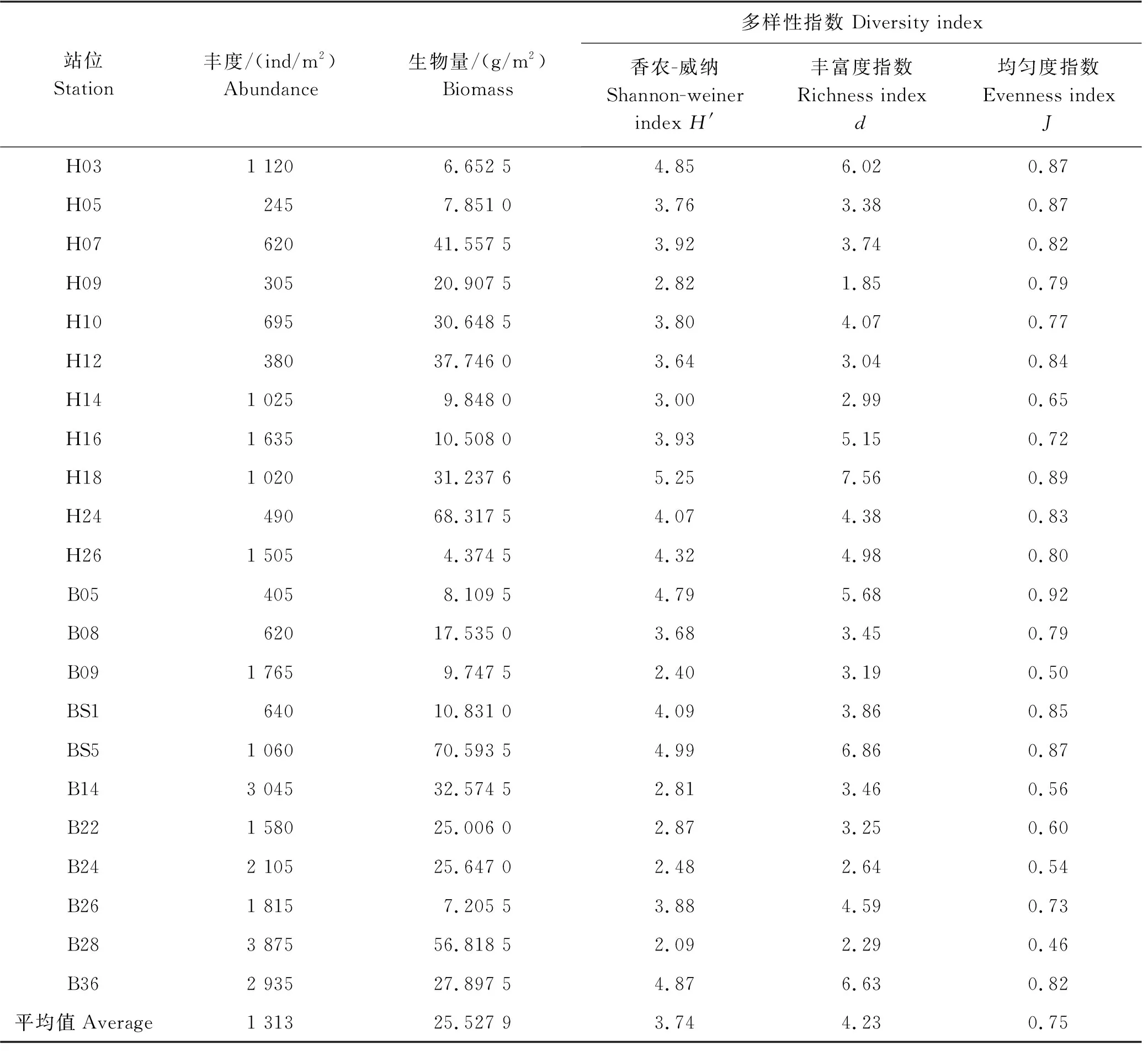
表3 调查海域大型底栖动物的丰度、生物量和多样性指数

图2 调查海域大型底栖动物丰度和生物量分布
2.4 大型底栖动物的多样性
调查海域冬季各站位多样性指数见表3。多样性指数(H′)的平均值为3.74,其中:高值站位分布于近岸海域,最高值出现在山东半岛南岸的H18站(5.25);位于北黄海中部4个站位的H′都低于3.00,最低为B28站(2.09)。丰富度指数(d)的平均值为4.23,高值站位分布同H′,最高值也出现在H18站(7.56),最低值则出现在南黄海中部的H09站(1.85)。均匀度指数(J)平均值为0.75,山东半岛东岸的B05站J值最高(0.92),北黄海中部的B28站J值最低(0.46)。
2.5 大型底栖动物的群落
本研究依据调查冬季海域大型底栖动物的丰度矩阵,对各站位进行了等级聚类分析(CLUSTER),使用组平均法,在22%相似度水平上将22个站位划分为4个群落(见图3),经单因素相似性分析(ANOSIM),各群落之间存在显著差异(R=0.78,P<0.01)。

图3 调查海域大型底栖动物聚类分析图(组平均法)
图4是根据聚类分析结果划分的4个群落分布图。

图4 调查海域大型底栖动物群落分布
群落Ⅰ: 位于北黄海中部,组内平均相似性为39.83%,包括5个站位,群落的优势种为薄壳索足蛤(T.tokunagai)、中蚓虫(M.californiensis)、太平洋方甲涟虫(Eudorellapacifica)和浅水萨氏真蛇尾(O.sarsiivadicola)。
群落Ⅱ: 位于南黄海中部,组内平均相似性为33.12%,包括10个站位,群落的优势种为太平洋方甲涟虫(E.pacifica)、薄壳索足蛤(T.tokunagai)、纽虫(Nemertinea spp.)、中蚓虫(M.californiensis)和浅水萨氏真蛇尾(O.sarsiivadicola)。
群落Ⅲ: 位于山东半岛东部沿岸,仅B05一个站位,群落的优势种为黄岛长吻虫(Saccoglossushwangtauensis、中蚓虫(M.californiensis)和拟特须虫(Paralacydoniaparadoxa)。
群落Ⅳ: 位于山东半岛近岸海域,组内平均相似性为28.60%,包括6个站位,群落的优势种为背毛背蚓虫(Notomastuscf.aberans)、纽虫(Nemertineaspp.)、寡节甘吻沙蚕(Glycindegurjanovae)和巴氏钩毛虫(Sigambrabassi)。
2.6 大型底栖动物的丰度与环境因子的关系
Pearson相关性分析是通过线性回归分析来确定2组变量间相互依赖的定量关系。本研究中大型底栖动物丰度与环境因子数据的相关性分析结果见表4,环境因子包括水深、底温、底盐、Chla含量、Pha含量、有机质含量、含水率和中值粒径。结果表明,调查海域冬季大型底栖动物丰度同Chla含量呈显著正相关,同底温呈显著负相关,同其余环境因子无显著相关性。

表4 调查海域大型底栖动物丰度与环境因子的Pearson相关性
2.7 大型底栖动物的群落结构与环境因子的关系
生物数据与环境因子最佳匹配分析(BIOENV)旨在通过计算环境因子和生物样品两者矩阵间的等级相关系数,测得同生物群落最佳匹配的环境变量子集[30]。本研究中BIOENV分析结果表明,同群落结构关系最为密切的单个环境因子为水深(秩相关系数rs=0.483);同群落结构关系最为密切的环境因子组合为水深+底温(秩相关系数rs=0.525)。
3 讨论
3.1 环境因子对调查海域大型底栖动物分布的影响
本研究中Pearson相关性分析结果(见表4)表明调查海域大型底栖动物丰度同Chla含量呈显著正相关,同底温呈显著负相关。
有研究[32]表明沉积物中Chla含量是底栖初级生产较为可靠的指示因子,即Chla含量的高低反映了底栖硅藻的多寡,通过硅藻→小型底栖动物→大型底栖动物的食物链传递营养和能量,进而影响大型底栖动物的丰度分布。
本研究调查海域底温的分布趋势为北低南高(见表1),丰度分布则刚好相反。李荣冠[33]分析了1998—2000年黄海近40个站位的数据发现,北黄海大型底栖动物的丰度高于南黄海,冬、春季尤为明显。孙昕雨[34]对2011年黄海大型底栖动物的研究发现,冬季航次的丰度高值区域出现在北黄海冷水团附近,丰度分布为北高南低。李新正[3]也指出,北黄海大型底栖动物的丰度比南黄海高出一个数量级。这些研究都与本研究得到的丰度分布趋势相一致,造成该现象的原因有待深入研究。
本研究中BIOENV分析结果表明,与群落结构关系最为密切的单个环境因子为水深,与群落结构关系最为密切的环境因子组合为水深+底温。
黄海海域的水深分布为西浅东深,近岸浅远岸深(见表1),沿此梯度对应的群落依次为广温性群落、边缘性群落和冷水性群落,这种对应关系同刘瑞玉等[8]、刘卫霞[20]、徐勇等[21]的研究结果一致。本研究中广温性群落与边缘性群落分界不明显,可能与近岸站位设置较少有关。
山东半岛沿岸流、黄海冷水团以及黄海暖流的相互作用,影响着黄海的底温分布,决定了黄海大型底栖动物的区系性质和组成分布[8]。本研究中,近岸的群落Ⅲ、Ⅳ受山东半岛沿岸流的影响较大,优势种以广温性种为主;远岸的群落Ⅰ、Ⅱ受黄海冷水团的影响较大,优势种以冷水性种为主;黄海暖流的作用加剧了远岸海域底温南高北低的趋势等[8],因此刘瑞玉等[8]与本研究的冷水性群落明显分成南北2个群落(见表5)。
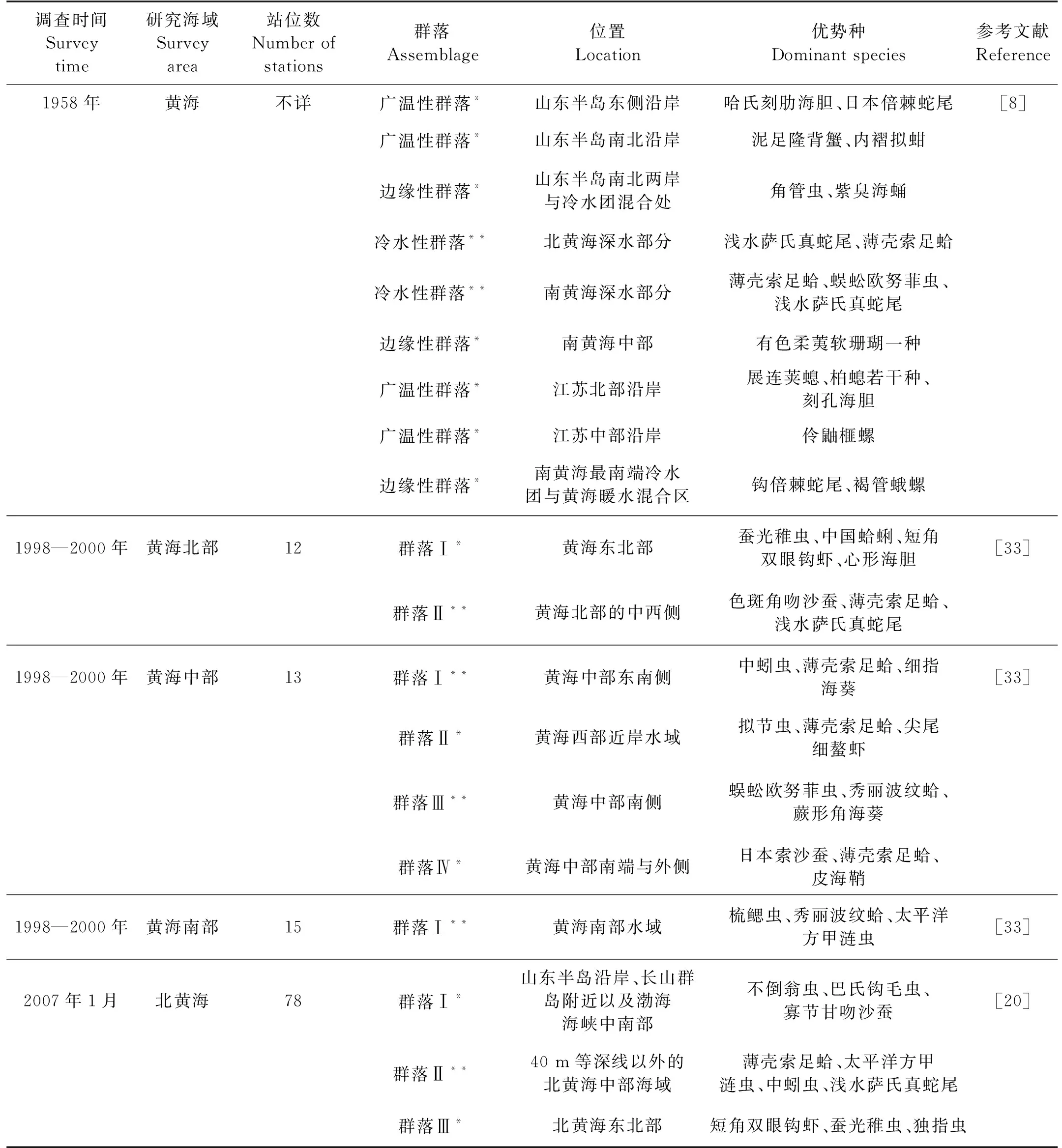
表5 本研究群落划分结果与相关研究的对比Table 5 Comparison of assemblage division results in this study with correlated studies

续表5
环境中各种理化因子对底栖动物群落结构有很大的影响[35],水深+底温的组合可以较好的解释本研究的群落结构和分布。
3.2 调查海域大型底栖动物群落结构的变化
表5为本研究与历史文献对黄海海域大型底栖动物群落划分的对比。刘瑞玉等[8]于上世纪中叶,首次将黄海的大型底栖动物划分为冷水性群落、广温性群落和边缘性群落。近20年来的有关研究[20-21,33]和本研究,对黄海大型底栖动物群落的划分与刘瑞玉[8]的划分基本一致。
近60年以来,黄海远岸的冷水性群落的优势种组成变化不大,仍为大个体冷水性种,如薄壳索足蛤(T.tokunagai)、浅水萨氏真蛇尾(O.sarsiivadicola)等。20世纪90年代之后的研究[20,33]和本研究中远岸群落的优势种组成中,出现了中蚓虫(M.californiensis)、太平洋方甲涟虫(E.pacifica)等小型个体种,这可能与网筛孔径的选择有关[36],1991年的《海洋调查规范》将网筛孔径的规定由1.0 mm改为0.5 mm[37]。因原有的大个体冷水性种仍存在,故可认为远岸群落结构变化较小。这可能与黄海冷水团常年低温的特性有关[38],有研究表明黄海冷水团底层水温、盐的年际变化很小[39-40],为大型底栖动物提供了稳定的生存环境[41]。
与远岸群落相比,黄海近岸群落的优势种类群组成变化较大。刘瑞玉等[8]研究中的近岸群落优势种,包括了腔肠动物、多毛类、软体动物、甲壳类和棘皮动物等类群(见表5)。张均龙[12]分析了1992年南黄海的资料,发现近岸群落的优势种组成已呈现以多毛类为主的现象,原有的大个体优势种数量显著降低甚至消失;2000年后的研究[20-21,42]及本研究的近岸群落,优势种也呈现单一类群化现象,即多毛类占优,且多为小型个体种,如中蚓虫(M.californiensis)、背毛背蚓虫(N.cf.aberans)、巴氏钩毛虫(S.bassi)、拟特须虫(P.paradoxa)和寡节甘吻沙蚕(G.gurjanovae)等(见表5)。出现这种优势种类群转变的现象,可能与后来航运、捕捞、排污等人类活动增加有关[20]。
4 结论
(1)近60年以来,黄海远岸群落的优势种组成仍以冷水性种为主;近岸群落的优势种组成呈现单一类群化现象,多毛类增多,且多为小型个体种。
(2)与大型底栖动物丰度显著相关的环境因子为Chla含量和底温,与群落结构关系最为密切的环境因子组合为水深+底温。
