乌苏里鼠李茎叶的解剖结构及其生态适应性
2023-05-21吴青松刘英卉李盼盼张友民
吴青松 刘英卉 李 硕 李盼盼 张友民
(吉林农业大学园艺学院,长春 130118)
乌苏里鼠李(Rhamnus ussuriensis)隶属于鼠李科(Rhamnaceae)鼠李属(Rhamnus),分布在我国黑龙江、吉林、辽宁、内蒙古、河北北部和山东(昆嵛山),国外主要见于俄罗斯西伯利亚和远东地区,朝鲜和日本也有分布[1]。乌苏里鼠李是一类用途广泛的小乔木或灌木,种子含油量大,可制作润滑油,耐干旱和耐贫瘠,是重要的水土保持和生态修复灌木树种[2],在园林绿化中常用于庭院、公园、风景区的绿化[3-5]。胡海清等[6]研究发现,乌苏里鼠李具有较好的抗火性,在森林防火方面具有重要价值。迄今为止,对乌苏里鼠李的研究主要集中在乌苏里鼠李的分类、生理等方面[7-11]。然而,关于其植物形态解剖特征及其生态适应性方面的研究资料相对较少。枝叶营养器官的形态和结构特征反映了植物适应环境的能力和机制,可为评价植物的抗性提供理论依据[12]。李蒙蒙等[13]研究认为叶表皮有表皮毛覆盖,气孔密度越大的植物抗旱性越强;沈萩荻等[14]研究认为叶柄中维管束越发达,抗旱能力就越强;赵君茹等[15]研究认为植物叶片栅栏组织越发达,中脉越厚的植株抗旱性越强;陈模舜等[16]研究认为,幼茎皮层内维管束数量多,维管束木质部导管数量多,直径大,髓部薄壁细胞发达的植物抗旱性强;蒋迎红等[17]研究认为老茎的髓部面积大,木质部发达,导管数量多、直径大的植物抗旱性强。以上的研究结果表明,植物枝叶的内部结构与其抗旱性密切相关。
本研究对乌苏里鼠李叶片和茎的结构特征进行系统的研究,探讨其结构与所处生境的关系,为乌苏里鼠李抗旱性鉴定和今后可持续开发利用提供科学依据。
1 材料与方法
1.1 试验材料
供试材料取自吉林农业大学园林试验基地,于2021 年6 月选取3 株生境和长势相一致的乌苏里鼠李,取成熟叶片和3 年生枝及1 年生枝。从叶基部以上叶片1/3 处(保留主脉),将叶片切成1 cm×1 cm 的小块,茎切成1 cm 长的小段,切好后立即放入FAA 固定液(V(甲醛)∶V(冰乙酸)∶V(50%乙醇)=1∶1∶18)中固定,然后将多年生植株的茎用软化剂(V(95%乙醇)∶V(丙三醇)=1∶1)软化15~30 d。
1.2 试验方法
采用常规石蜡切片法切片,叶片和初生茎的切片厚度为12 µm,次生茎的切片厚度为16 µm,番红—固绿复染,中性树胶封片[18]。所得样片用生物显微镜(LEICA-DM2500,上海)观察、拍照和测量。
扫描电子显微镜样品的制备:参考王玉等[19]研究方法,将处理过的叶片经梯度乙醇脱水,放入电热鼓风干燥箱(101A-1ET,上海)60 ℃条件下干燥3.5 h 将材料放置在粘有双面胶的样品台上,后用离子溅射仪(ETD-2000,上海)导电处理,溅射镀膜厚7~8 nm,在扫描电子显微镜(Shimadzux-550,日本)下观察并拍照。
光学显微镜样品的制备:采用刀片分别刮去上下表皮叶肉细胞,滴入0.5%固绿染液,V(无水乙醇)∶V(二甲苯)=1∶1 溶液冲洗,中性树胶封片,放入恒温箱中35 ℃保存,在生物显微镜(LEICADM2500,上海)下观察、拍照与测量[20]。
1.3 数据及分析
每个样片在同倍数的光学显微镜下随机取5个视野,每个视野测量10 个数据,采用Excel 2019对所得数据进行整理,计算平均值和标准差。
2 结果与分析
2.1 叶表皮超微特征和叶片解剖结构
2.1.1 扫描电子显微镜下叶的表皮特征
扫描电镜下,乌苏里鼠李叶上、下表皮蜡质纹饰相似,为皱脊状(见图1:A~B)。气孔和表皮毛只存在叶下表皮(见图1:C~D),气孔形状为椭圆形,气孔两侧的蜡质纹饰向下凹陷(见图1D)。
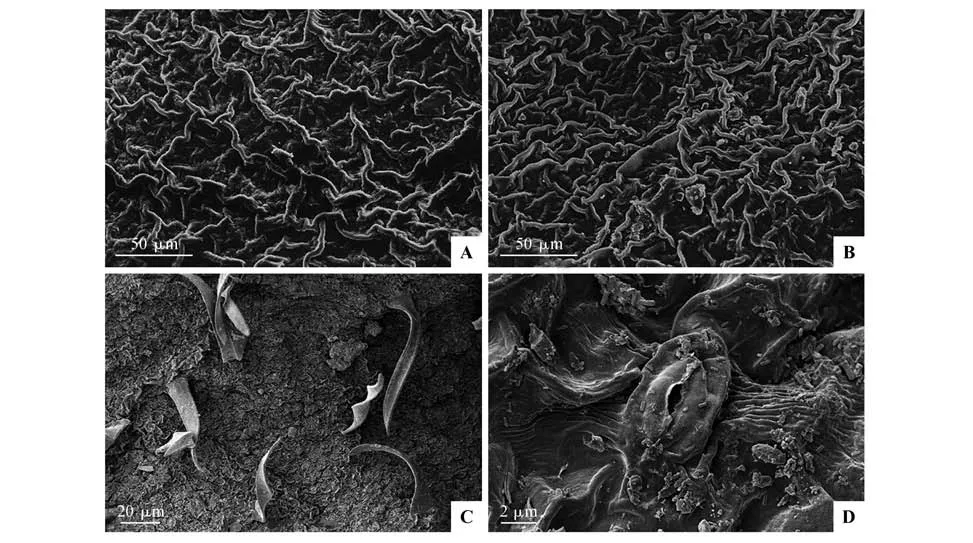
图1 扫描电镜下乌苏里鼠李叶片上(A)、下表皮(B~D)超微特征Fig.1 Ultrastructural characteristics of upper and lower epidermis of Rh.ussuriensis leaves under scanning electron microscope
2.1.2 光学显微镜下叶的表皮特征
光学显微镜下,乌苏里鼠李叶上表皮无气孔分布,表皮细胞形状不规则,表皮细胞垂周壁类型为深波状、A 型(见图2A)。叶下表皮分布气孔,为椭圆形,气孔器类型为无规则型,气孔器由2个肾型保卫细胞和气孔组成,无副卫细胞,表皮细胞形状不规则,表皮细胞垂周壁类型为浅波状、G型(见图2B)。
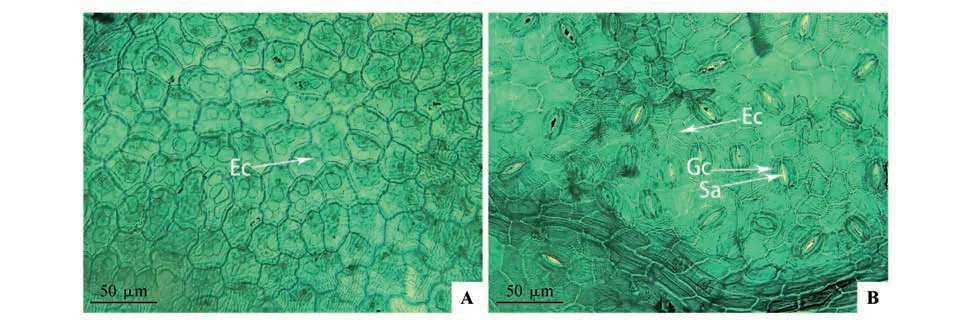
图2 光学显微镜下乌苏里鼠李叶片上(A)、下表皮(B)特征Ec.表皮细胞;Gc.保卫细胞;Sa.气孔Fig.2 The characteristics of the upper and lower epidermis of Rh.ussuriensis leaves under the optical microscopeEc.Epidermal cell;Gc.Guard cells;Sa.Stomata
2.1.3 叶的解剖结构
由图3 可知,乌苏里鼠李的叶片横切面由表皮、叶肉和叶脉3 部分组成,叶肉薄壁组织分化为栅栏组织和海绵组织,为典型的异面叶。
叶表皮结构特征:乌苏里鼠李其上、下表皮均由1 层排列紧密的椭圆形表皮细胞组成(见图3A),上表皮细胞的平均厚度31.63 µm,外覆盖厚5.15 µm 的角质层,无气孔分布。下表皮细胞平均厚度23.43 µm,外覆盖厚3.33 µm 的角质层,有气孔分布(见图3B)。
叶肉结构特征:栅栏组织由2层排列整齐而紧密的长柱形薄壁细胞组成,平均厚度为136.29 µm,海绵组织由2~3 层形状不规则的细胞组成,细胞间隙大,排列疏松,平均厚度为107.31 µm,栅海比为1.40,在栅栏组织与海绵组织之间有维管束分布,叶片平均厚度为337.86 µm(见图3B)。
主脉结构特征:乌苏里鼠李叶片主脉发达,由机械组织、基本组织和维管束组成。表皮由1层排列紧密的细胞组成,外覆较厚的角质层,叶脉维管束为无限外韧维管束,呈半月状。木质部靠近近轴面,韧皮部靠近远轴面,二者之间有微弱的形成层分布,木质部由11~13 列导管组成。在维管束周围环绕着大量的红色单宁的异细胞、晶簇及靠近远轴面的细胞之间存在细胞间隙,形成了通气组织(见图3:C~D)。
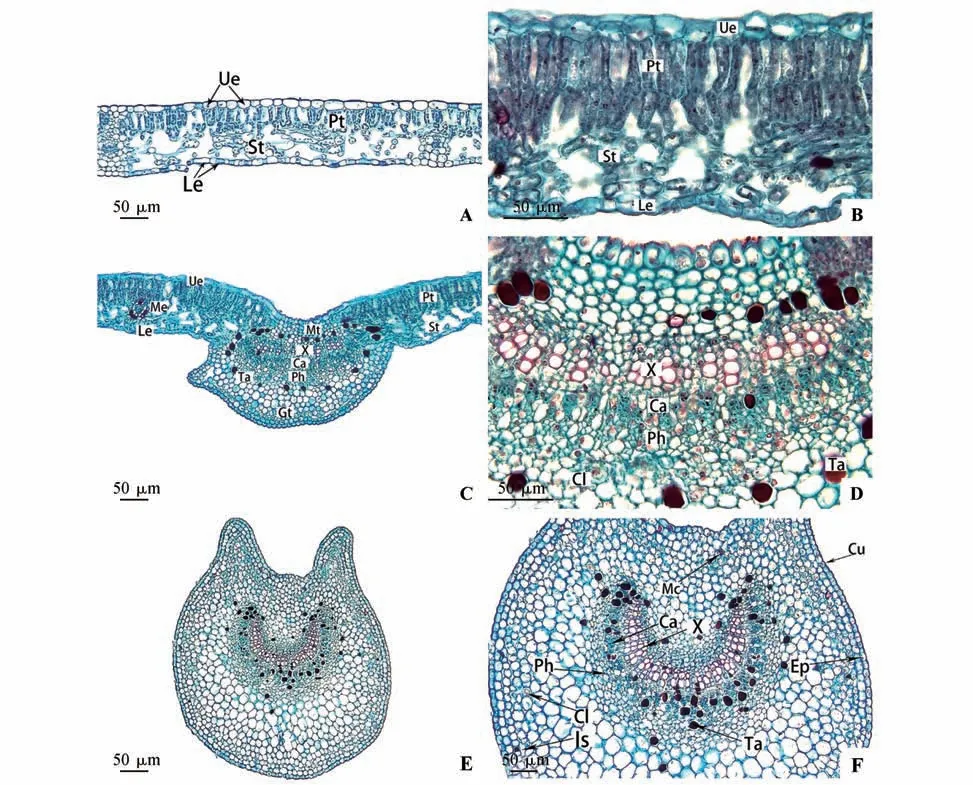
图3 乌苏里鼠李叶的解剖结构A.叶中脉两侧的解剖结构;B.叶中脉两侧的解剖结构;C.叶主脉处的解剖结构;D.叶主脉处的解剖结构;E.叶柄横切面轮廓;F.叶柄横切面维管束形态;Ue.上表皮;Pt.栅栏组织;St.海绵组织;Le.下表皮;Me.叶肉;Mt.机械组织;Gt.基本组织;X.木质部;Ca.形成层;Ph.韧皮部;Ta.单宁;Cl.晶簇;Cu.角质层;Ep.表皮;Mc.黏液细胞;Is.细胞间隙Fig.3 Anatomical structure of R.ussuriensis leafA.Anatomical structure on both sides of leaf midvein;B.Anatomical structure on both sides of leaf midvein;C.Anatomical structure of leaf main vein;D.Anatomical structure of leaf main vein;E.The outline of petiole transverse section;F.Vascular bundle morphology on petiole transverse section;Ue.Upper epidermis;Pt.Palisade tissue;St.Spongy tissue;Le.Lower epidermis;Me.Mesophyll;Mt.Mechanical tissue;Gt.Ground tissue;X.Xylem;Ca.Cambium;Ph.Phloem;Ta.Tannins;Cl.Cluster;Cu.Cuticles;Ep.Epidermis;Mc.Mucous cells;Is.Intercellular space
叶柄结构特征:乌苏里鼠李叶柄由表皮、基本组织、维管束3 部分组成。叶柄的横切面近圆形,中间有凹陷。表皮细胞为椭圆形,排列紧密,表皮细胞的平均厚度为37.23 µm,外覆盖4.59 µm 厚的角质层。维管束的类型为外韧无限维管束,由木质部、形成层和韧皮部组成,成半圆形排列,木质部导管分子数量众多且排列紧密,导管直径大,其平均直径为45.12 µm。叶柄内分布着大量的单宁异细胞和少量的黏液细胞及晶簇(见图3:E~F)。
2.2 茎的解剖结构特征
2.2.1 初生茎结构特征
乌苏里鼠李初生茎由表皮、皮层和维管柱3个部分组成(见图4A)。表皮由单层椭圆形细胞组成,排列紧密,外覆较厚的角质层,平均厚度为4.34 µm。皮层位于表皮的内方,介于表皮和维管柱之间,有多层细胞,细胞壁较厚,排列紧密。皮层外方为3~4 层的厚角组织,中皮层为8~10 层的薄壁组织细胞,细胞体积较大,呈圆形,含有晶簇,内皮层不明显(见图4:A~D)。乌苏里鼠李初生茎的维管柱包括维管束、髓和髓射线。维管束为外韧无限维管束,13~14 个维管束围绕髓排列1 轮,维管束由初生木质部、束中形成层和初生韧皮部组成(见图4:A~B)。初生木质部中导管3~4 个为1 列,导管的直径较大,每个维管束有2~6 列管孔链,初生木质部的发育方式为内始式。由初生茎的纵切结构可知,初生茎的导管主要有环纹导管和螺纹导管2种类型(见图4C)。初生茎的最中心是髓,较为发达,由近圆形的薄壁细胞组成,薄壁细胞由中心向四周逐渐变小,中心处细胞最大(见图4A)。
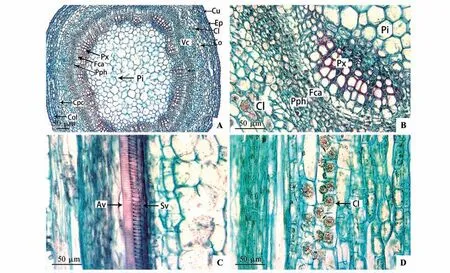
图4 乌苏里鼠李初生茎解剖结构A~B.初生茎横切面;C~D.初生茎纵切面;Cu.角质层;Ep.表皮;Co.皮层;Col.厚角组织;Cpc.皮层薄壁细胞;Vc.维管柱;Cl.晶簇;Pph.初生韧皮部;Fca.束中形成层;Px.初生木质部;Pi.髓;Av.环纹导管;Sv.螺纹导管Fig.4 Anatomy of primary stem of Rh.ussuriensisA-B.Transverse section of primary stem;C-D.Primary stem longitudinal section;Cu.Cuticles;Ep.Epidermis;Co.Cortex;Col.Collenchyma;Cpc.Cortical parenchyma cells;Vc.Vascular cylinder;Cl.Cluster;Pph.Primary phloem;Fca.Fascicular cambium;Px.Primary xylem;Pi.Pith;Av.Annular vessel;Sv.Spiral vessel
2.2.2 次生茎结构特征
乌苏里鼠李次生茎主要由周皮、次生维管组织和髓组成,周皮呈褐色,由木栓层、木栓形成层和栓内层组成,周皮上分布皮孔(见图5A)。次生维管组织由次生韧皮部、维管形成层、次生木质部和维管射线组成(见图5A)。次生木质部中导管多为复管孔和孔团,导管类型主要以孔纹导管和螺纹导管为主,管壁上存在具缘纹孔,多为互列式(见图5:B~D)。导管分子的平均长度为489.86 µm,平均直径为52.68 µm。木射线由薄壁细胞组成,维管形成层由2~3 层排列紧密的薄壁组织细胞组成。韧皮部较厚,细胞排列紧密。木质部年轮显明,是典型的环孔材(见图5A)。髓位于最中心,由排列紧密的多边形薄壁细胞组成。乌苏里鼠李木射线属于异形Ⅱ型,主要由单列射线和双列射线组成,单列射线多,双列射线少,由直立细胞和横卧细胞组成,直立细胞有3 层,横卧细胞有2 层(见图5:E~F)。

图5 乌苏里鼠李次生茎解剖结构A~B.次生茎横切面;C~D.次生茎纵切面;E.次生茎纵切面;F.次生茎弦切面;Le.皮孔;Pd.周皮;Phf.韧皮纤维;Sx.次生木质部;Ve.导管;Wr.木射线;Px.初生木质部;Pi.髓;Far.第1 年年轮;Sar.第2 年年轮;Tar.第3 年年轮;Wpc.木薄壁细胞;Pv.孔纹导管;Sv.螺纹导管;Prc.横卧射线细胞;Urc.直立射线细胞;Dr.双列射线;Sr.单列射线Fig.5 Anatomical structure of secondary stems of Rh.ussuriensisA-B.Transverse section of secondary stem;C-D.Longitudinal section of secondary stem;E.Longitudinal section of secondary stem;F.Secondary stem tangential section;Le.Lenticel;Pd.Periderm;Phf.Phloem fiber;Sx.Secondary xylem;Ve.Vessel;Wr.Wood ray;Px.Primary xylem;Pi.Pith;Far.First annual ring;Sar.Second annual ring;Tar.Third annual ring;Wpc.Wood parenchyma cells;Pv.Pitted vessel;Sv.Spiral vessel;Prc.Procumbent ray cell;Urc.Upright ray cell;Dr.Double ray;Sr.Single ray
3 讨论与结论
3.1 叶片结构特征对环境的适应
植物与其生长的环境关系密切,其形态结构特征最能体现对环境的适应性,为了适应周围环境和满足生长需要,形成了不同的表型特征[21-22]。叶片作为应对环境变化较敏感的器官,其外部形态和解剖结构特征的变化能较好地反映出植物对干旱等逆境的适应性[23]。任哲等[24]和王爽等[25]研究中发现叶片越厚其储水能力越强,蒸腾速率越低,抗旱性也越强。潘昕等[26]发现叶片栅栏组织厚,海绵组织薄,栅海比大的植物更加耐旱。张金玲等[27]发现叶片主脉越厚,木质部越发达,叶片的水分输导效率也越高,叶片越耐旱。许少祺等[28]研究指出海绵组织疏松的植物对湿生环境适应性好,耐寒性强。本研究中乌苏里鼠李叶片较厚,栅栏组织发达,栅栏组织由2 层薄壁组织细胞组成,结构紧密,海绵组织排列疏松,栅栏组织的厚度厚于海绵组织,栅海比达到1.4。乌苏里鼠李叶片主脉较厚,主脉维管束发达,木质部导管数量多,直径大,输水效率高。乌苏里鼠李这些结构特征均可以说明乌苏里鼠李耐旱性和耐寒性较强。陈燕等[29]和陈健辉等[30]在研究中发现叶片内富含单宁不仅可以阻碍水分流动,避免过度蒸腾引致生理干旱,还可以在盐渍环境中更好地生长。在本研究中发现乌苏里鼠李叶片维管束周围环绕着大量的单宁,因此可以推断乌苏里鼠李具有很强的耐旱性,同时还对盐渍环境有一定的适应能力。
管东旭等[31]研究发现叶片较厚的角质层可以降低蒸腾,提高抗旱能力。高艳等[32]研究认为气孔器集中于下表皮,可以使气孔不易受到直接的机械伤害,避免阳光直射而减少气孔蒸腾。王孟珂等[33]研究认为高光合效率的植物一般气孔小而多,密度大,以减少水分散失。尤凤丽等[34]认为表皮毛可以反射强光照射,减少水分蒸发。本研究中乌苏里鼠李叶上下表皮都有较厚的角质层,气孔集中分布在下表皮,气孔直径较小,密度大,在下表皮上附着着表皮毛,由此可以说明乌苏里鼠李的叶片具有抵御干旱环境的能力。
3.2 茎的结构特征对环境的适应
茎作为植物运输水分和营养物质的重要器官,其内部结构会影响植物的抗旱能力。胡云等[35]研究认为茎表皮外被角质层可以减少水分蒸腾、增强抗旱性。张般般等[36]研究发现,茎皮层较厚的植物保水性和抗旱性更强。钱雪等[37]指出发达的髓可以增强茎的贮水能力,滕丽等[38]发现维管束发达,维管束导管数量多的植物更能适应干旱环境。乌苏里鼠李茎的初生结构中表皮外被角质层,皮层较厚,皮层细胞排列紧密,维管束发达,维管束导管数量多,直径大,髓部所占面积大,细胞排列紧密,较为发达,这些结构增强了乌苏里鼠李的抗旱性,使其更好的适应干旱环境。
乌苏里鼠李茎的次生结构中,木质部发达,导管数量多,个体较大,形状近圆形。与黄萍等[39]研究中茎的结构相似,这些结构说明乌苏里鼠李的茎具有较高的水分输送效率,进而提高植株的抗旱能力。上官方京等[40]研究认为木质部中直径大的导管,具有更高的输水效率和导水率,来满足干旱环境下蒸发需求的加剧,乌苏里鼠李木质部导管的直径大,这表明乌苏里鼠李能够适应干旱环境。
综上所述,乌苏里鼠李叶片和茎的形态结构特征表明,乌苏里鼠李能够很好的适应干旱环境,这是其长期适应环境而进化的结果。
