基因Cdca2敲除对小鼠精子发生和生育力无明显影响
2023-05-04邹定峰缪时英
柳 俊,邹定峰,缪时英,宋 伟,李 凯
中国医学科学院基础医学研究所 北京协和医学院基础学院 生物化学与分子生物学系医学分子生物学国家重点实验室,北京 100005
不孕不育症是一个全球性医学问题。研究报告表明40%~50%的不孕不育症的病例是由男性因素导致的,主要表现为精子发生障碍和系统性疾病[1]。精子发生是发生在动物睾丸及附睾内的高度复杂、协调、有序的动态过程,涉及复杂的调控机制。
研究表明部分RNA结合蛋白(RNA binding protein, RBP)在转录基因表达的调节中起重要作用,进而影响精子发生过程。如Nanos2编码一种锌指RNA结合蛋白,缺失将导致精原干细胞(spermatogonial stem cell, SSC)缺失,过表达则会导致精原干细胞堆积[2-3]。Lin28A作为精原干细胞的标志物之一,也是一种RNA结合蛋白,其具有促进精原干细胞增殖的功能[4-5]。
细胞分裂周期相关蛋白2(cell devision cycle associated 2,CDCA2) 是一种与细胞周期相关的RNA结合蛋白[6]。有研究表明在细胞的有丝分裂过程中,CDCA2 可与蛋白磷酸酶1(protein phosphatase 1,PP1) 结合,确保核膜重组过程顺利进行[7]。此外,CDCA2 还可参与染色质重塑和DNA损伤修复等过程[8-9]。本研究利用 CRISPR/Cas9技术结合Cre-loxP介导的重组系统对睾丸高表达的基因Cdca2进行敲除并获得了纯合的生殖细胞特异性敲除小鼠品系。通过对Cdca2组织特异性敲除小鼠进行雄性生殖功能的表型分析,为研究该编码基因的功能提供了参考。
1 材料与方法
1.1 材料
1.1.1 动物: SPF 级 C57BL/6J 雄鼠 12只,雌鼠12只,体质量20~25 g(斯贝福生物技术有限公司); SPF 级成年Cdca2floxed杂合子雄鼠3只,雌鼠3只(委托北京百奥赛图基因生物技术有限公司构建);Cdca2生殖细胞特异性敲除雄鼠12只(本实验所构建);上述动物均在中国医学科学院基础医学研究所动物中心饲养。饲养标准为 3~5只/笼,温度区间为20 ℃ ~24 ℃,湿度区间为50%~70%,光照时间为8∶00~ 20∶00,期间自由饮水摄食。
1.1.2 主要试剂:AG RNAex Pro RNA提取试剂(艾科瑞生物技术有限公司);互补DNA(cDNA)反转录试剂盒(北京全式金生物技术有限公司);动物睾丸组织固定液(武汉赛维尔生物科技有限公司);SDS-PAGE 快速凝胶试剂盒(上海雅酶生物科技有限公司);BCA蛋白定量试剂盒(Thermo Fisher Scientific公司); Anti-CDCA2一抗(艾比玛特医药科技上海有限公司,36263);Anti-beta-actin一抗(Proteintech公司, 66009); 山羊抗小鼠IgG/辣根酶标记(北京中杉金桥生物技术有限公司,ZB-2305);250 bp-DNA ladder(上海捷瑞生物工程有限公司);琼脂糖(Biowest公司);引物(北京天一辉远生物科技有限公司合成)(表1)。
1.2 方法
1.2.1 CDCA2系统发育树的构建:多蛋白序列比对由MAFFT进行比对,系统发育树由IQ-TREE软件构建。进化距离标注在分支线上,代表进化支变化的程度,越短代表差异越小,进化距离越近。
1.2.2 检测小鼠各部位组织mRNA和蛋白质:将8周龄的成年雄性小鼠用脱颈法处死,解剖小鼠,获得小鼠的肺、心、脾、肾、脑、肝、睾丸、附睾等组织,将组织放入离心管中,并加入1 mL Trizol试剂 (RNA提取液), 于冰上超声裂解组织5 min。随后将样本室温放置 5 min 使其得到充分裂解,加入 200 μL氯仿,充分震荡混匀、室温放置10 min后,于离心机中12 000 r/min,4 ℃离心 15 min,吸取离心管中的上层水相至新管中并加入等体积异丙醇,上下颠倒混匀,室温静置约10 min。12 000 r/min,4 ℃离心10 min后弃上清液留 RNA沉淀于管底。加入预冷过的1 mL DEPC水配置的75%乙醇洗涤 RNA沉淀, 7 500 r/min,4 ℃离心 10 min后弃上清液,再次重复洗涤。 打开离心管室温晾干,加入 DEPC水溶解,并测量 RNA浓度。将提取后的 RNA 按照反转录试剂盒TransScript First-Strand cDNA Synthesis SuperMix说明进行,每个样品反转录 1 μg RNA。Q-PCR体系(每孔20 μL,每个样品3个重复孔)配置如下: Q-PCR Mix 10μL,前、后向引物各 0.4 μL,cDNA0.5 μL, ddH2O 2 μL。 Q-PCR引物序列见 (表1)。 Q-PCR反应条件:预变性 95 ℃, 5 min, 扩增和荧光信号收集(95 ℃ 30 s,60 ℃ 30s)×40 cycle,熔解曲线 (60 ℃ 30 s,+0.3 ℃/s,90 ℃ 15 s)用上述方法取得小鼠各组织后,根据每个组织的大小向 离心管中加入适量SDS裂解液,添加蛋白酶及磷酸酶抑制剂,于冰上进行超声裂解。裂解后冰上静置5 min, 随后100 ℃电浴10 min,12 000 r/min离心5 min,取上清,部分BCA测定、部分制样(浓度:4 g/L)。于预制胶中80 V电泳30 min,120 V电泳1 h。320 mA转膜1 h 30 min。5%脱脂牛奶封闭1 h。使用anti-CDCA2和anti-Actin的一抗, 4 ℃过夜孵育。TBST洗3次,每次10 min。二抗孵育1 h, TBST洗3次,每次10 min。随后进行显影。
1.2.3 生殖细胞特异性敲除Cdca2小鼠的构建:使用CRISPR/Cas9技术构建Cdca2floxed杂合子小鼠,外显子5和7两侧有两个loxP位点和同源臂。为了生成Cdca2floxed纯合小鼠品系,将获得的杂合子小鼠,相互交配,得到Cdca2floxed纯合雌性小鼠。将Cdca2floxed纯合雌性小鼠与组织特异性Cre的雄性小鼠Ddx4-Cre(在胚胎第15天雄性生殖细胞中表达)交配,以获得Cdca2floxp/+-Cre雄性小鼠。最后将Cdca2floxed纯合雌性小鼠与Cdca2floxp/+-Cre雄性小鼠交配获得生殖细胞特异性敲除Cdca2的Cdca2floxp/floxp-Cre小鼠,方案及原理见(图2B)。
1.2.4 鉴定小鼠基因型:剪取小鼠尾尖约5 mm,置于1.5 mL 离心管中,加入50 μL碱性组织裂解液,100 ℃电浴40 min。加入50 μL酸性溶液中和,震荡混匀。12 000 r/min离心2 min去除鼠毛杂质, 将获得的鼠尾 DNA样品进行 PCR鉴定其基因型。 PCR体系(每个样品20 μL)配置如下:2×Taq PCR StarMix 10 μL,上下游引物各1 μL,鼠尾DNA样品6 μL,ddH2O 2 μL。PCR扩增 条件:95 ℃ 5 min,(95 ℃ 25 s,64.5 ℃ 30 s,72 ℃ 40 s)×35 cycle,72 ℃ 2 min,16 ℃保持。PCR 后进行琼脂糖凝胶电泳,根据DNAladder及电泳条带位置判断基因型。
1.2.5 小鼠睾丸石蜡切片苏木精-伊红(hematox-ylin-eosin dyeing,HE)染色:8周龄的野生型小鼠和Cdca2floxp/floxp-Cre小鼠用脱颈法处死,收集睾丸和附睾,在动物睾丸组织固定液中固定48 h,随后按照常规的苏木精-伊红(HE)染色流程制片,并用中性树脂封片,自然晾干后置于显微镜下拍照观察。
1.2.6 小鼠精子计数、活率、运动参数分析:取野生型和Cdca2floxp/floxp-Cre成年雄鼠各3只,脱颈法处死,收集一侧附睾精子于获能培养液(DMEM F12+10%胎牛血清) 中, 37 ℃ 5% CO2中培养30 min,进行体外获能。用精子全自动检测分析系统(computer assisted sperm analysis,CASA) 检测精子数量、运动性及精子活率。每组重复3次,取平均值比较差异有无统计学意义。
1.2.7 小鼠生育力测试:分别取8周龄的雄性Cdca2floxp/floxp-Cre和野生型雄性小鼠各3只,将其与12只野生型C57BL/6J成年雌鼠按1∶2的比例合笼,交配3个月。定期对雌鼠检栓,确保交配。记录每只孕鼠平均每胎所生的幼崽数量。
1.3 统计学分析
2 结果
2.1 CDCA2在小鼠睾丸中高表达
Cdca2位于小鼠14号染色体上,全长39.5 kb, 其编码的蛋白由982个氨基酸组成。首先通过公共数据库NCBI搜索了CDCA2在人类中的表达模式,发现其在睾丸组织中高表达(图1B)。通过多序列比对构建了CDCA2的蛋白系统发育树,结果表明CDCA2在哺乳类动物中具有较高的保守性(图1A)。为了证实其在小鼠中的表达,取小鼠的不同组织进行了Q-PCR和Western blot检测,结果显示其在睾丸组织高表达(图1C, D)。
2.2 构建生殖细胞特异性敲除Cdca2小鼠模型
成功构建了Cdca2floxed杂合子小鼠(图2A),经过与Ddx4-Cre工具鼠配繁,得到与Cdca2floxp/+-Cre雄性小鼠。将其与Cdca2floxed纯合雌性小鼠交配获得生殖细胞特异性敲除Cdca2的Cdca2floxp/floxp-Cre小鼠(图2B, C),并通过小鼠基因型鉴定证实。与野生型小鼠相比,Cdca2floxp/floxp-Cre小鼠生长、发育和行为均表现正常(图2D)。
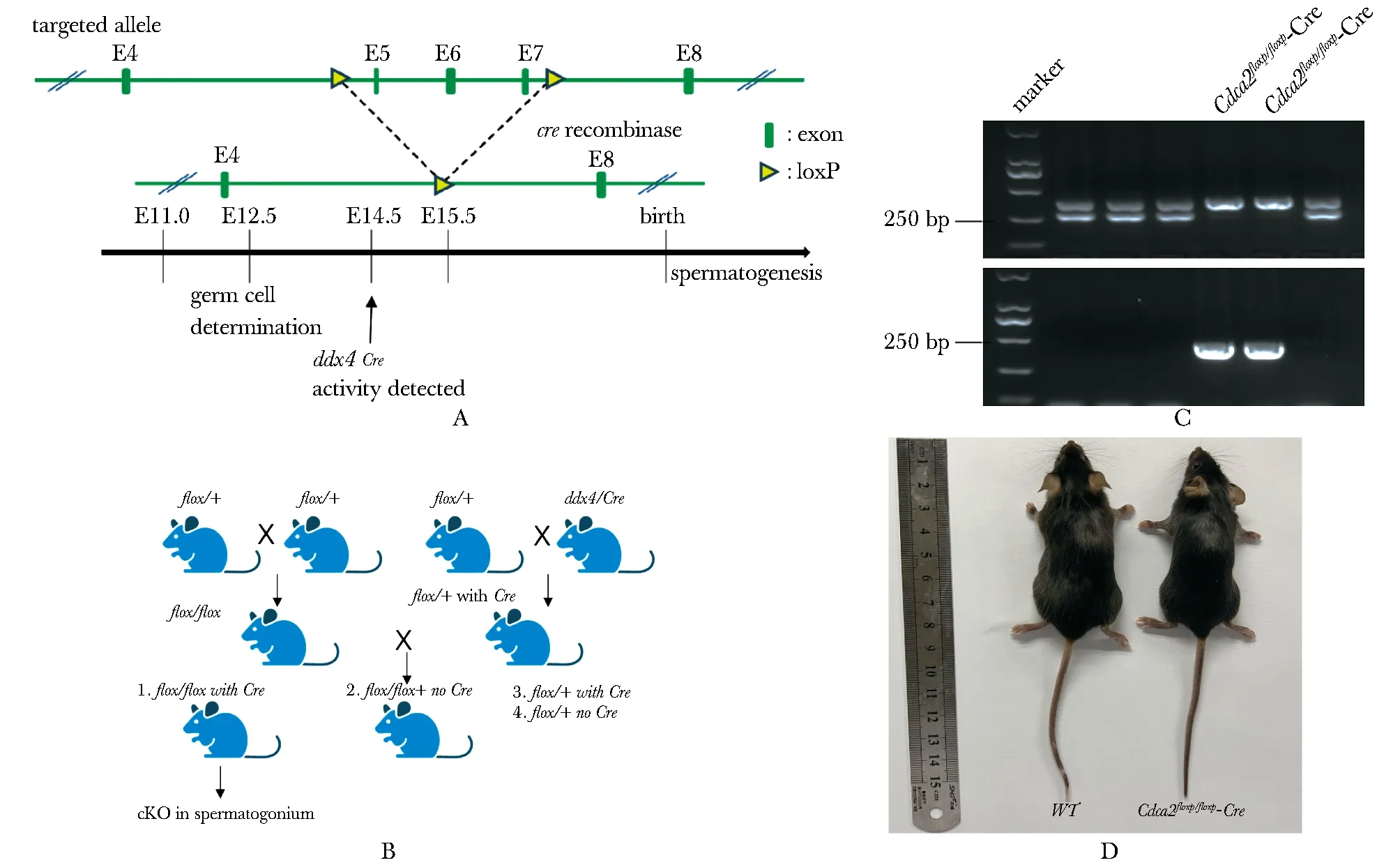
A.schematic diagram of Cdca2 conditional knockout mouse construction; B.mating diagram of germ cell conditional gene knockout mice; C.genotype identification of Cdca2 conditional gene knockout mice; D.photos of adult wild type(WT) mice and Cdca2 conditional knockout mice
2.3 CDCA2缺失对小鼠精子发生和生育力无明显影响
在成功构建生殖细胞特异性敲除Cdca2floxp/floxp-Cre小鼠后,对小鼠进行生育力测试,发现Cdca2floxp/floxp-Cre小鼠与野生型小鼠相比,繁育的小鼠数量几乎相同(图3E),并且睾丸大小和质量之间的差异无统计学意义(图3A,B)。随后比较了Cdca2floxp/floxp-Cre小鼠与野生型小鼠的精子浓度,发现Cdca2floxp/floxp-Cre小鼠的精子浓度仅下降了5%(图3C)。HE染色结果显示生殖细胞特异性敲除Cdca2小鼠的睾丸曲细精管具有正常的结构包括从精原干细胞到精子等各级生精细胞(图3D)。此外还利用计算机辅助精液分析系统测定了Cdca2floxp/floxp-Cre小鼠与野生型小鼠的精子活率及精子运动状况(图3E),包括精子正常形态与异常形态百分比(图4A)、精子运动直线性(图4B)、精子平均鞭打频率(图4C)、精子平均运动性(图4D)、精子进行性运动性(图4E)、以及精子头部平均侧摆幅值(图4F)等。各组精子数、精子活动能力和运动参数差异均无统计学意义。这些发现表明睾丸高表达基因Cdca2的缺失对小鼠的精子发生和生育能力没有明显影响。
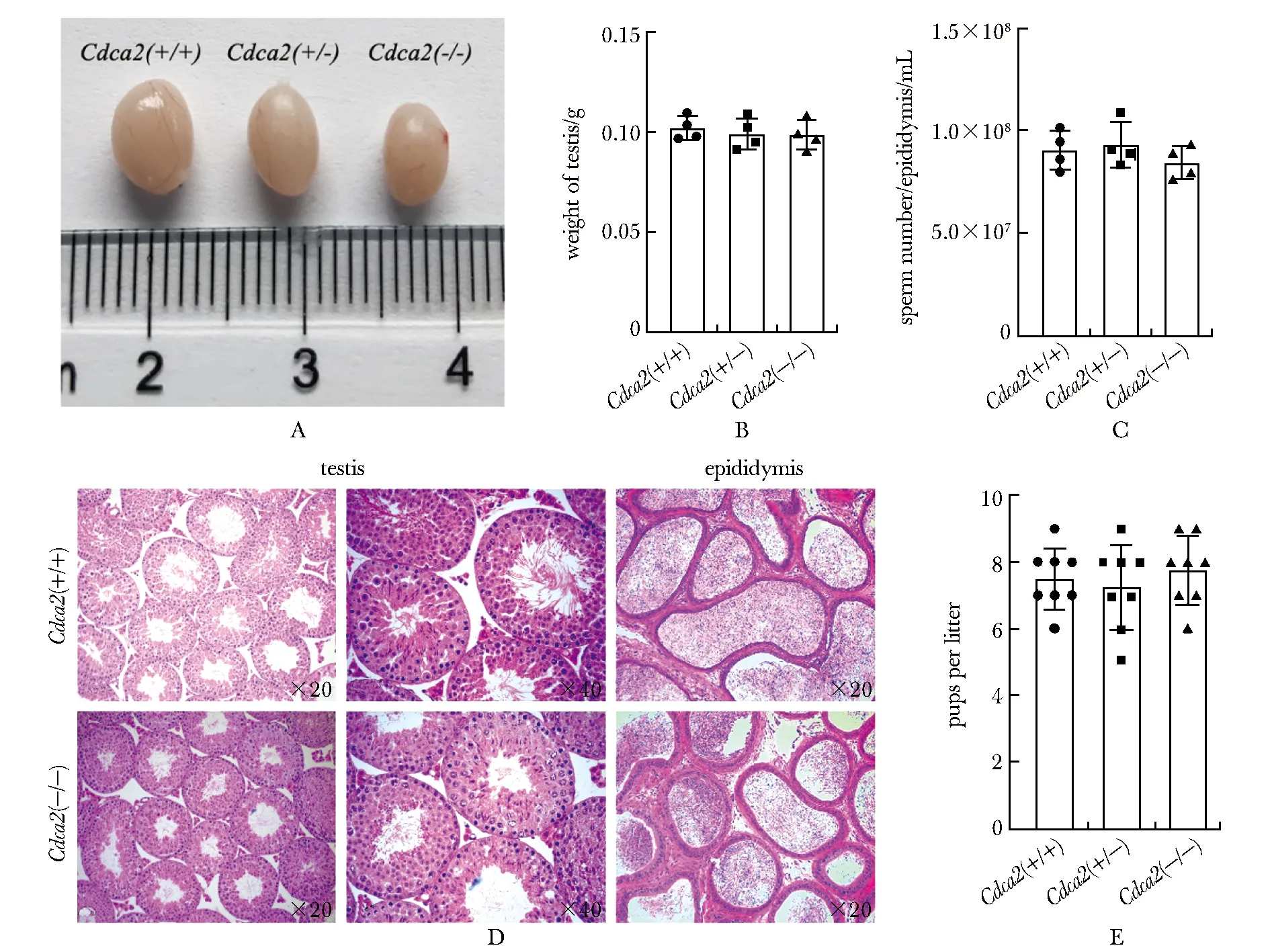
A.typical image of testis from wild type(WT) and Cdca2 germ cell specific knockout mice aged 8 weeks; B.testis weight of WT and Cdca2 germ cell specific knockout mice aged 8 weeks; C.sperm count of 8-week old WT and Cdca2 germ cell specific knockout mice; D.hematoxylin-eosin(HE) staining of testis and epididymis sections from WT and Cdca2 germ cell specific knockout mice aged 8 weeks; E.fertility test of 8-week old WT and Cdca2 germ cell specific knockout mice
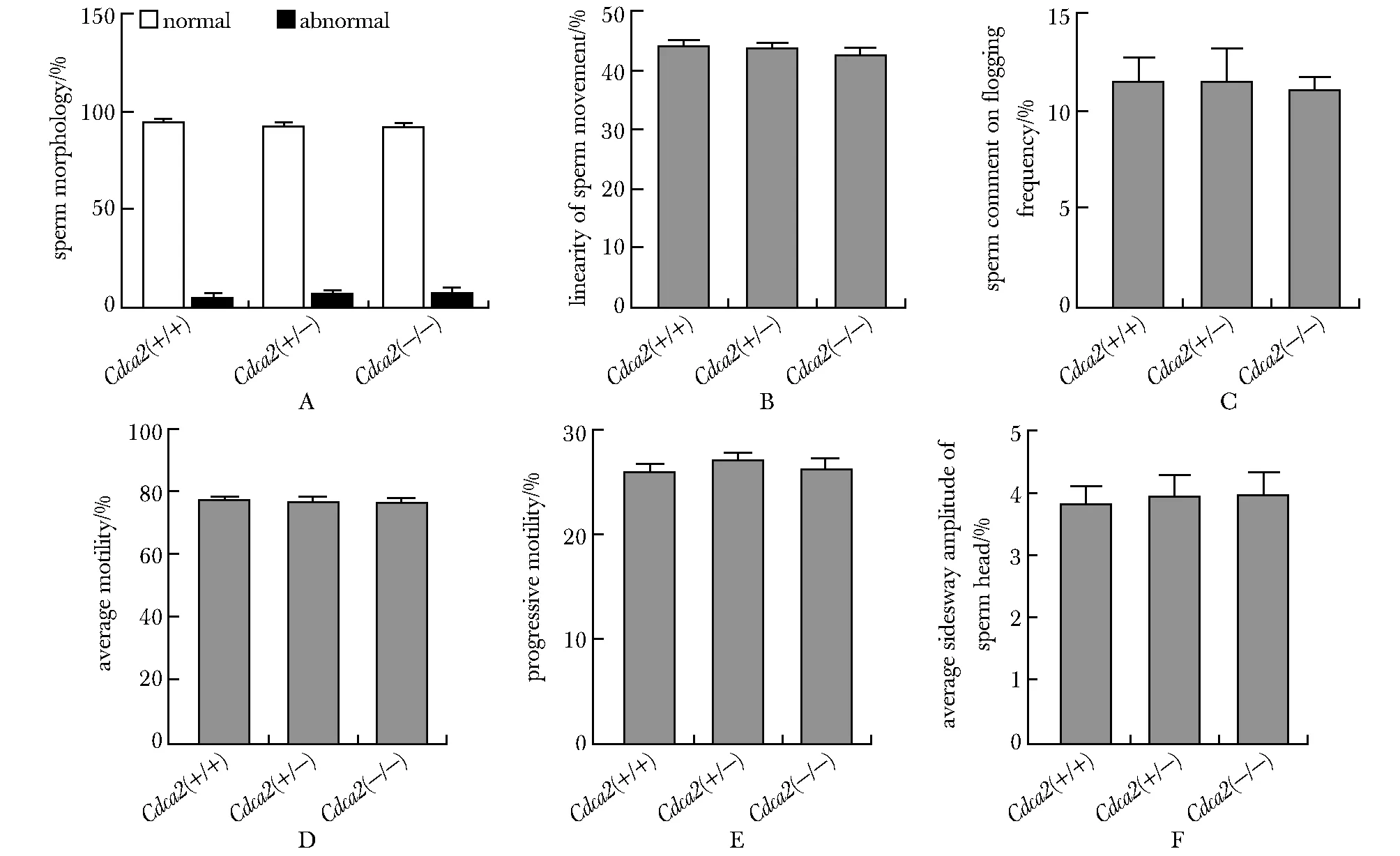
A.percentage of normal and abnormal sperm morphology; B.linearity of sperm movement; C.sperm comment on flogging frequency; D.average motility of sperm; E.progressive sperm movement; F.average sidesway amplitude of sperm head
3 讨论
目前有大量研究鉴定了睾丸特异表达基因在雄性生殖中的功能及机制。研究表明睾丸高表达基因数量约为1 000个,其中约400个睾丸高表达基因被研究人员证实对精子发生和雄性生殖具有重要意义[10-11]。仍有大量睾丸高表达基因未被证实是否在精子发生过程中起到重要作用。
Cre-loxP介导的重组系统是一种强大的基因编辑工具,可以实现组织特异性基因编辑,因其高效性、准确性、快速性、时空特异性的优势,已广泛应用于转基因动物的构建[12]。细胞分裂周期相关蛋白 (cell division cycle-asociated protein,CDCAs)家族共计 8 个成员(CDCA1~8)[13]。其中CDCA2在细胞周期中参与核膜的重组、调控 DNA 损伤,并在人和小鼠睾丸组织中高表达。本研究证明Cdca2在小鼠睾丸组织中高表达,再通过构建生殖细胞特异性敲除Cdca2小鼠模型,分析了Cdca2缺失后小鼠的精子发生和生育能力,发现Cdca2缺失后对小鼠精子发生和生育力并无明显影响,此外CASA分析表明Cdca2缺失后小鼠的精子活率和活动性与野生型小鼠相当。
RNA测序结果显示Cdca2在精母细胞及精子细胞阶段高表达,这表明CDCA2可能在精子减数分裂及精子形成时期发挥作用。但结果显示生殖细胞特异性敲除Cdca2后,小鼠的精原细胞增殖、精母细胞减数分裂及精子形成阶段均未观察到明显损伤。但这并不意味着生殖细胞缺失Cdca2在分子水平没有发生明显变化。后续可以结合转录组测序技术(RNA sequencing,RNA-seq)分析检测Cdca2缺失后生殖细胞在转录水平是否改变。本研究发现,尽管Cdca2在睾丸组织中高表达,但在雄性生育中Cdca2可能是其功能冗余,或CDCA家族的其他成员可能会补偿其功能。这些数据将有助于研究人员筛选对生育、精子发生至关重要的基因,并避免研究重复,为后续进一步确定CDCA2功能提供了一定的研究基础。
