雷州湾海域网衣污损生物群落的周年变化特征
2023-04-25周家丽傅建融曾嘉维陈志劼王学锋
周家丽,傅建融,曾嘉维,陈志劼,王学锋*,刘 丽
[1.广东海洋大学水产学院,广东 湛江 524088;2.南方海洋科学与工程广东省实验室(湛江),广东 湛江 524025]
网衣污损生物(亦称附着生物)是指固着或者栖息在养殖网衣部位、对人类经济活动产生不利影响的所有海洋生物的总称[1]。中国水产养殖产量多年居世界首位[2],这为民众提供了优质动物蛋白,但网衣因多缝隙、表面积大而成为污损生物的理想附着基,受其影响产生的养殖环境污染与动物病害等问题应引起广泛关注[3]。同时,随着渔业资源衰退,海洋渔业发生了由捕捞业向养殖业为主的巨大转变,1998年我国开始引进国外深水网箱技术,随后在全国沿海地区迅速发展,使深蓝渔业和渔业高质量发展也成为水产养殖可持续发展亟待解决的问题[4]。此外,养殖过程中投饵损失的饲料以及生物排泄所产生的丰富营养物质,进一步促进了养殖网衣附近水域的污损生物的生长[5]。污损生物随时间推移大量附着在网衣上而堵塞网孔,不仅影响养殖水体的交换和养殖对象的健康生长,也降低了网衣的使用年限[6]。污损生物种类组成复杂、防治困难,是生态学研究和健康养殖中面临的巨大挑战之一[7]。已有研究表明,污损生物具有种类多样、繁殖能力强、生活史短等特点,而且其生长具有规律性和阶段性[8],因此摸清污损生物的这些变化规律对网箱防污具有重要意义。
雷州湾位于我国粤西近岸海域,西沿雷州半岛,北有东海岛,东有硇洲岛,南至徐闻外罗港,湾口面向东南,水域辽阔,具有非正规半日潮特征,属于典型的热带亚热带季风气候,海域生物群落类型丰富,是我国污损生物生态调查研究的典型海区[9]。已有学者在该海域对原有的人工设施上的污损生物进行过取样[10-12],但尚无针对网衣污损生物群落的长时间变化及其与环境因子关系的研究报道。本文在雷州湾东南码头附近海域开展网衣污损生物周年挂网实验,旨在了解该海域网衣污损生物群落的变化规律,以期为网衣清洗、更换的时间和频次等防治措施的确定提供参考依据。
1 材料与方法
1.1 实验时间、地点
2020年6月至2021年6月,在雷州湾东南码头附近[13-14]网箱养殖集中区(N 20°55′、E 110°30′,图1)开展周年挂网附着实验,收集污损生物。

1.2 实验设计
实验网衣由网目30 mm的25 cm×25 cm聚乙烯(Polyethylene,PE)编制而成。实验共设2组,按不同的观测时间周期,每组均设季节周期网、半年周期网(上半年为12月至次年5月,下半年为6至11月)、全年周期网,分上、下两层悬挂,每个样本设4个平行样,上层距水面0.5 m,下层距水面4.5 m(图2)。网衣顶部固定于养殖平台上,底部用重锤绑缚,使网身与海底垂直。
1.3 取样与鉴定
回收的网衣样品保存在无水乙醇中,将网衣上的5 cm×5 cm生物全部取下,软体动物等大个体生物全部分析,端足类等小个体生物按每份样品1/10的量进行镜检,在光学显微镜下进行分类和计数,用吸水纸吸去生物体表的水分后,在电子天平上进行称重。分类单元名称依据《中国动物志》[15-18]及世界海洋物种登记网[19]进行交叉核对。
1.4 环境因子测定
每月初在挂网站点附近海域选取3个点进行环境因子检测,每次调查时间为13∶00~17∶00,由调查人员现场采用 YSI 水质检测仪测定溶解氧、pH、盐度和温度。依据GB/T 12763.6—2007《海洋调查规范 第6部分:海洋生物调查》[20]进行污损生物调查。
1.5 数据处理与分析
1.5.1 优势种
利用相对重要性指数(Index of relative importance,IRI)分析海洋污损生物群落的优势种。
IRI=(W+N)×F×104
(1)
式(1)中:W为某一特定物种的生物量占总生物量的百分比;N为某一特定物种的丰度占总丰度的百分比;F为某一物种的频率。IRI>25设为优势种。
海洋污损生物分为3种生活方式:运动(Motile,M)、固着(Sessiler,SE)、附着(Attached,A);按功能区分为6个功能性群组:初级生产者(Primary producers,PP)、悬浮饲养者(Suspension feeders,S)、草食动物(Herbivores,H)、食肉动物(Carnivorous,C)、杂食动物(Omnivores,O)和沉积物饲养者(Deposit feeders,D)[21]。
1.5.2 物种多样性
利用Excel对上、下水层不同时段观测周期网衣的污损生物丰度和生物量进行统计分析,群落多样性采用Shannon-Wiener 多样性指数(H′)、Pielou均匀度指数(J)和Margalef丰富度指数(d)进行分析。
H′=-∑(nt/N)log2(nt/N)
(2)
J=H′/log2S
(3)
d=(S-l)/lnN
(4)
式(2)~(4)中:ni为第i种个体数;N为所有种个体数;S为样品中总种类数。

1.5.3 多元统计分析
应用Excel和SPSS分别对样本进行多样性和单因素分析。应用GraphPad Prism 8.0软件对网衣污损生物群落多样性、丰富度、均匀度等指数作箱图对比。应用CANOCO 4.5软件对污损生物和水环境因子进行去趋势对应分析(Decentred correspondence analysis,DCA),当排序梯度最大值大于4.0时,选择典范对应分析(Canonical correspondence analysis,CCA)进行排序分析和作图;若在3.0~4.0之间,选择CCA或冗余分析(Redundancy analysis,RDA);若小于3.0,RDA的结果要好于CCA。
2 结果与分析
2.1 污损生物群落组成
2.1.1 种类组成
所有网衣共鉴定出污损生物35种(表1),分属11门28科35属,其中节肢动物门10种,占比28.57%;软体动物门8种,占比22.86%;扁形动物门、刺胞动物门、多孔动物门、红藻门、环节动物门、棘皮动物门、脊椎动物门、绿藻门、纽形动物门共计17种,总占比48.57%。

表1 东南码头污损生物种类组成Tab.1 Composition of fouling organisms in the Southeast Wharf

续表1

续表1
在时间上,春季(16种)、夏季(20种)、秋季(15种)、上半年(18种)、下半年(15种)、全年(26种)均高于冬季(7种)。各个时期的网衣中,节肢动物门均有显著优势,主要包括巴西地钩虾(Podocerusbrasiliensis)、加尔板钩虾(Stenothoegallensis)、指拟钩虾(Gammaropsisdigitata)等物种;其次为软体动物门,主要出现在夏、秋季,主要包括翡翠贻贝(Pernavirids)、斑马纹贻贝(Dreissenapolymorpha)等物种;再者为藻类,主要出现在春季,主要包括浒苔(Enteromorphaprolifera)、石莼(Ulvalactuca)等物种。
在空间上,下水层(32种)稍多于上水层(29种),污损生物种类主要包括巴西地钩虾、加尔板钩虾、指拟钩虾等物种。
2.1.2 优势种
全年网衣污损生物的优势种共有15种(表2),其中上水层12种、下水层15种。节肢动物占有绝对优势,有7种:指拟钩虾上、下水层IRI分别为4 648和3 645,网纹藤壶(Amphibalanusreticulatus)上、下水层IRI分别为3 948和2 425,加尔板钩虾上、下水层IRI分别为1 961和2 652,以及巴西地钩虾、夏威夷明钩虾、莫顿蜾蠃蜚(Corophiummortonii)、红巨藤壶(Megabalanusrosa);软体动物有4种:斑马纹贻贝、翡翠贻贝、变化短齿蛤(Brachidontesvariabilis)、厚壳贻贝(Mytiluscoruscus);藻类有3种:浒苔、石莼、异枝江蓠(Gracilariabailinae);刺胞动物1种:笔螅(Pennariadisticha)。
本次研究优势种根据污损生物的活动和生活方式,将其分为3类:固着型、附着型和运动型,结果表明固着生物和附着生物在污损生物中占优势,再根据功能性群体分类,确定了3个功能性群体,分别为S、D、PP群。在大多数实验网衣上,S群占优势,主要代表物种为贻贝;D群是第二大优势组,主要代表物种为钩虾;而PP群主要代表物种为浒苔。根据本研究确定的优势种功能群,共有2个营养水平:PP群代表第一营养级;S和D构成第二营养级(表2)。
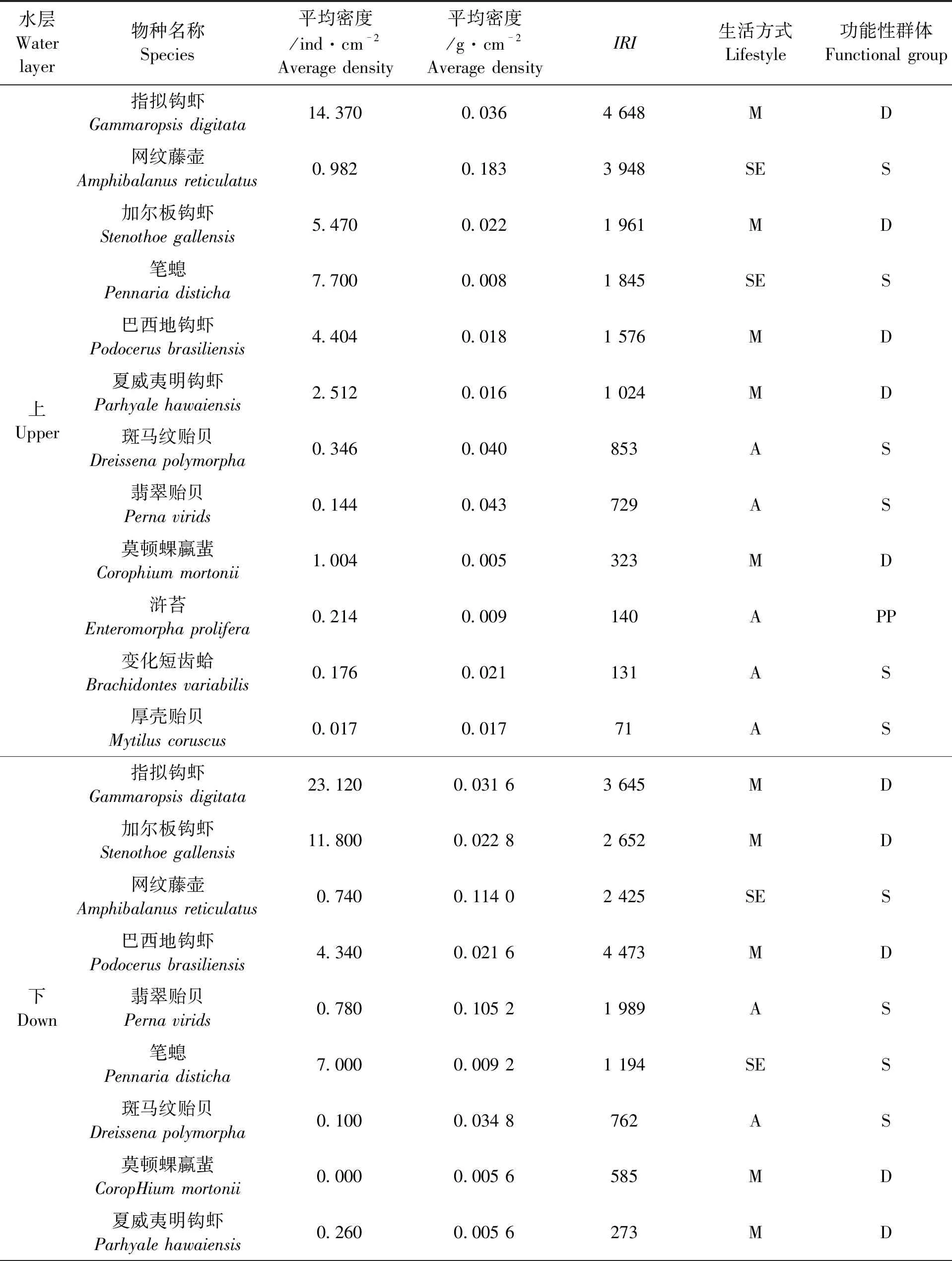
表2 网衣污损生物优势种Tab.2 Fouling biological dominant species of net

续表2
2.2 丰度和生物量
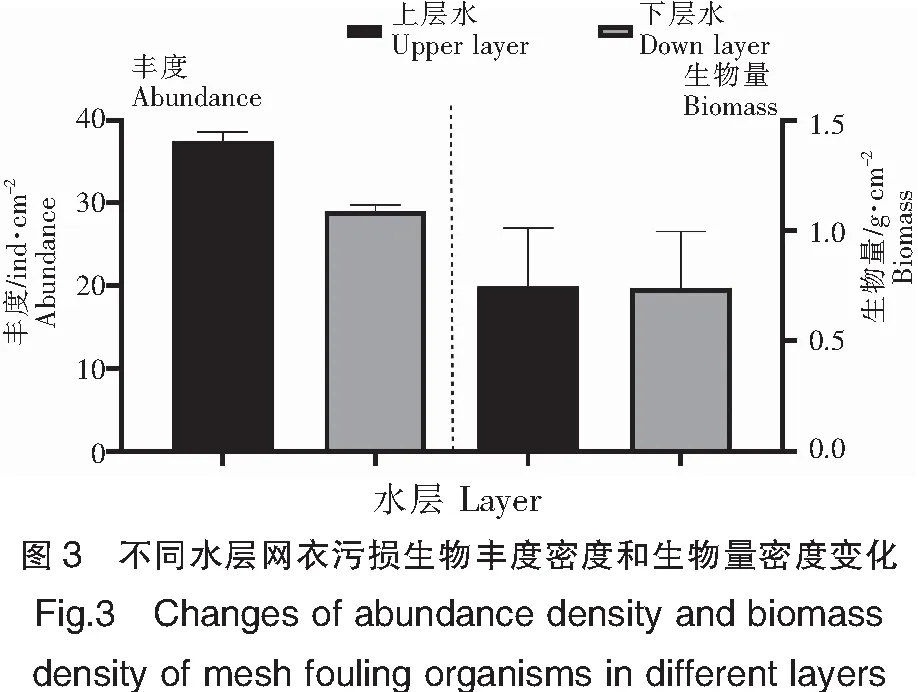
从空间上看,东南码头上水层的网衣污损生物丰度(37.53 ind·cm-2)显著高于下水层(29.03 ind·cm-2)(P<0.05);生物量(0.75 g·cm-2)高于下水层平均生物量(0.74 g·cm-2),但无显著性差异(P>0.05)(图3)。
从时间观测周期看,网衣污损生物的丰度和生物量分布存在明显差异,上、下水层的变化趋势均表现为夏季(月份)、半年、全年高于其他时间段,冬季最低,且时间观测周期越长,丰度和生物量数值越大(图4)。
2.3 多样性指数
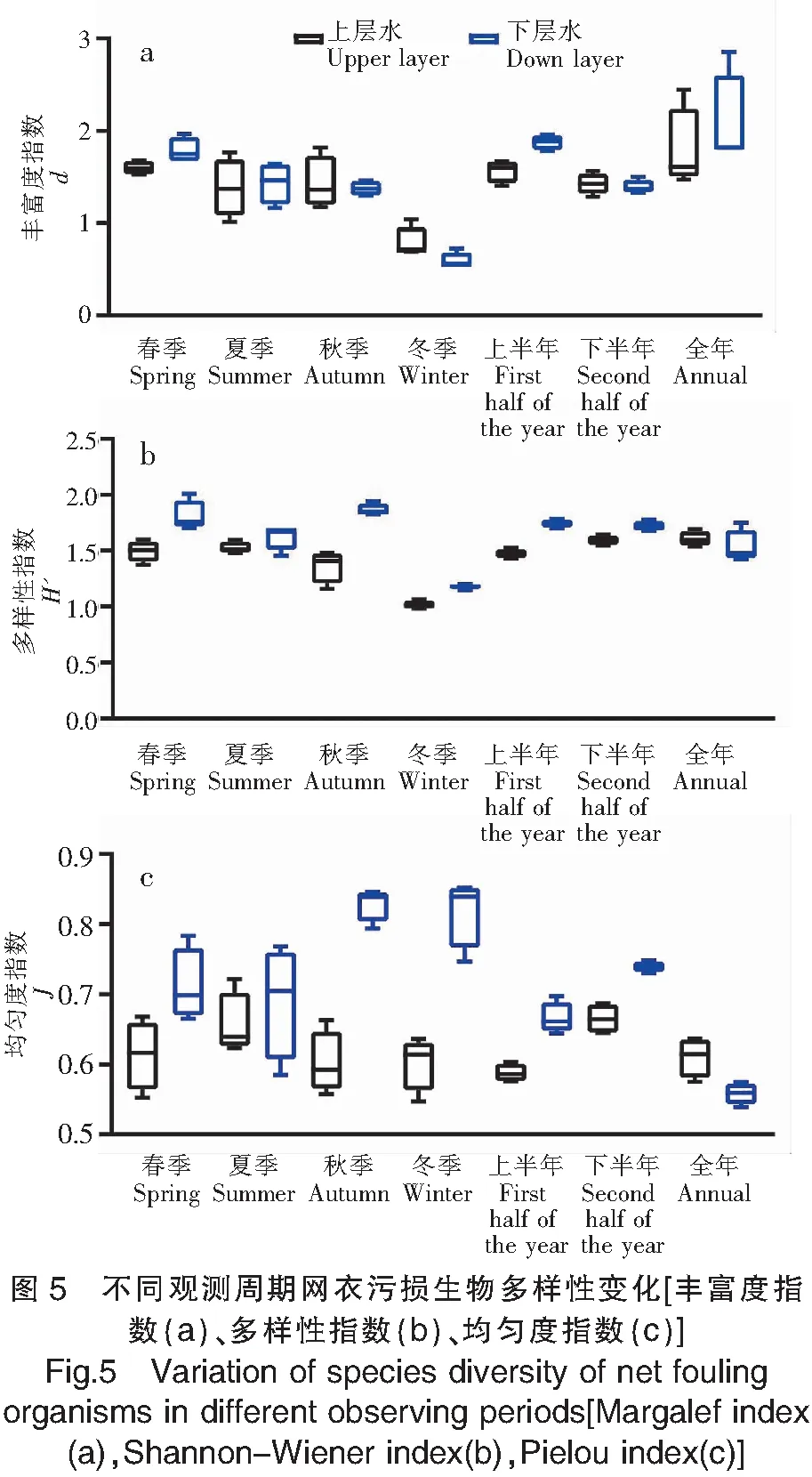
上、下水层的各个观测周期网衣多样性指数(0.985~2.010)、均匀度指数(0.539~0.852)、丰富度指数(0.539~2.856)数值相近。

网衣污损生物上、下水层7个观测周期网衣各指数的对比见图5,在空间上表现为下水层(1.149~2.010)多样性指数(H′)大体上高于上水层(0.985~1.693),丰富度指数(d)(上水层:0.688~2.443;下水层:0.539~2.856)、均匀度指数(J)(上水层:0.547~0.721;下水层:0.539~0.852)无显著变化规律;在时间上表现为冬季网衣污损生物群落多样性指数(H′)(春季:1.373~2.010;夏季:1.454~1.689;秋季:1.159~1.942;冬季:0.985~1.201;上半年:1.431~1.787;下半年:1.546~1.779;全年:1.423~1.751)、丰富度指数(d)(春季:1.527~1.966;夏季:1.014~1.769;秋季:1.173~1.820;冬季:0.539~1.040;上半年:1.405~1.958;下半年:1.289~1.567;全年:1.474~2.856)低于其他周期网衣,但均匀度指数(J)(春季:0.553~0.784;夏季:0.585~0.768;秋季:0.557~0.846;冬季:0.547~0.852;上半年:0.576~0.697;下半年:0.645~0.748;全年:0.539~0.636)各个观测周期无显著变化规律。
2.4 影响网衣污损生物群落分布的环境因子
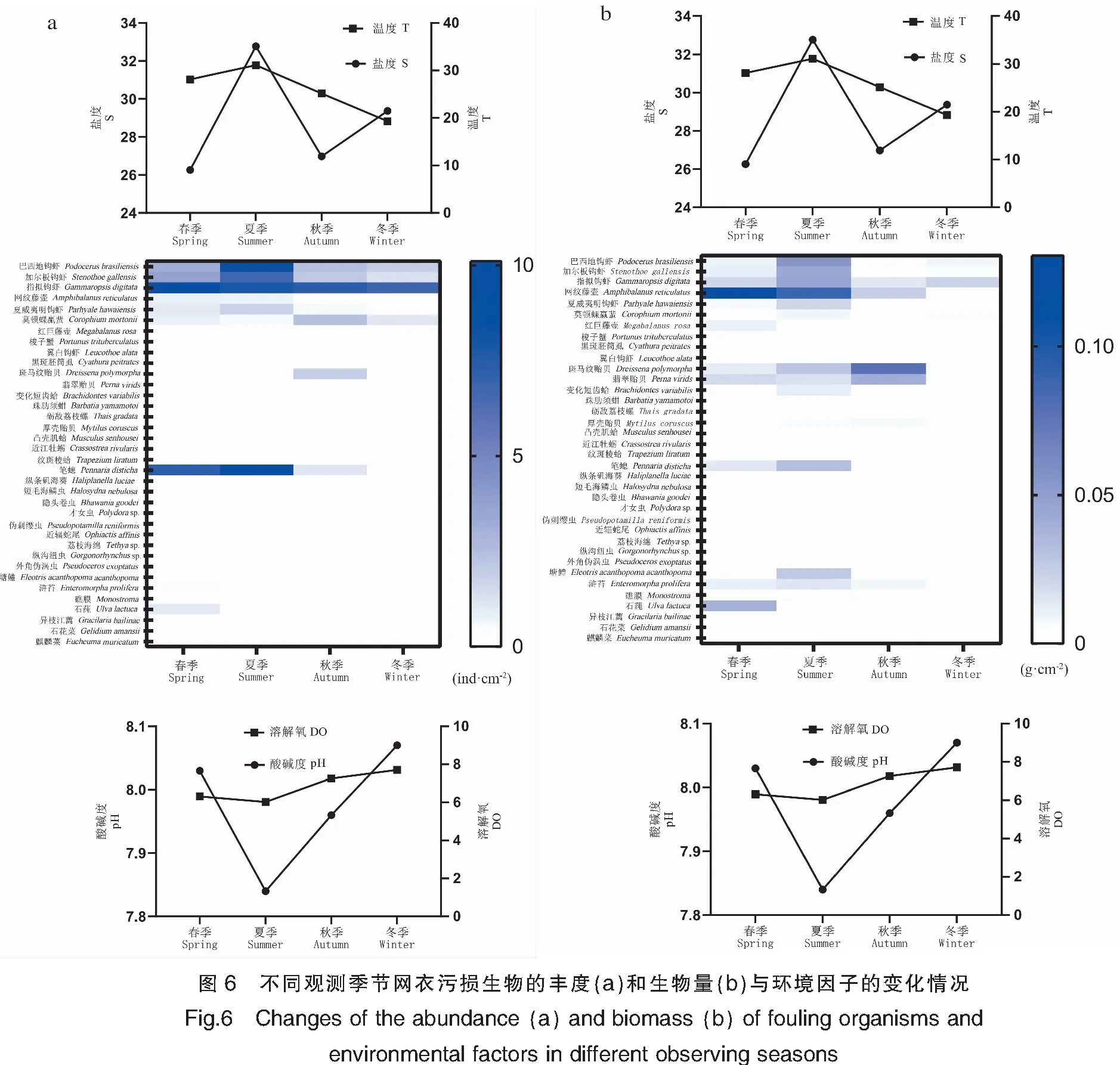
对4个季节时期网衣污损生物群落丰度和生物量进行热图结合环境指标分析,结果显示春、夏季的优势种丰度和生物量较其他季节均占优势地位,其优势种丰度和生物量与温度的关系基本呈正相关。优势种节肢动物钩虾类物种的生物量和丰度显示为春、夏季占据重要优势地位,其中指拟钩虾一年四季均有分布,春季的丰度和生物量更显著;藤壶物种的丰度和生物量显示为春夏季较显著;软体动物类斑马纹贻贝和翡翠贻贝的生物量在夏、秋季较显著;藻类石莼、浒苔、石花菜(Gelidiumamansii)等物种均显示为春季较显著;刺胞动物笔螅生物量和丰度显示为春夏季较显著(图6)。
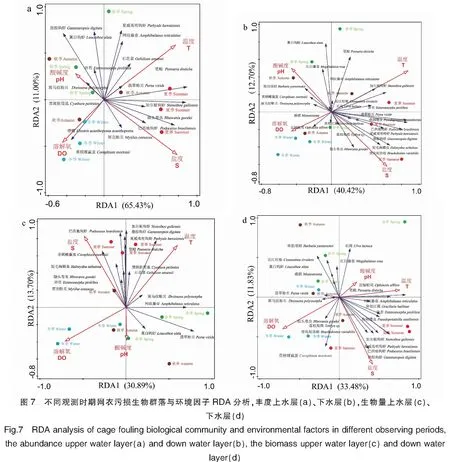
对4个季节观测时期网衣污损生物群落及东南码头的污损生物群落丰度和生物量进行DCA分析,排序结果显示上、下水层丰度和生物量 4 个轴最大长度分别为1.2、1.8、1.7、2.3,属于线性模型,因此再使用RDA分析环境因子对4个季节网衣污损生物群落的影响。在不同季节的上、下水层污损生物丰度、生物量-环境相关性在RDA1轴、RDA2轴相关性分别为30.89%~65.43%、11.00%~13.70%(图7)。1)上、下水层各环境因子对污损生物群落结构(种类组成丰度密度)的贡献率分别为63.6%、32.8%。分析结果均显示,盐度(上、下水层贡献率分别为40.1%、28.2%)和温度(上、下水层贡献率分别为57.4%、59.0%)为主要影响因子,盐度对夏季损生物分布影响较大,呈负相关;温度对春季、秋季、夏季的污损生物分布有极显著影响,呈正相关。2)上水层各环境因子对污损生物群落结构(种类组成生物量密度)贡献率为18.3%,分析结果均显示,盐度(贡献率为38.3%)和温度(贡献率为39.1%)为主要影响因子,温度对夏季污损生物分布有显著影响,呈正相关;盐度对夏、秋季影响较大,呈负相关。下水层各环境因子对污损生物群落结构(种类组成生物量密度)贡献率为32.7%,分析结果均显示,盐度(贡献率为21.0%)和温度(贡献率为24.3%)为主要影响因子,温度对春、夏季污损生物分布有极显著影响,呈正相关。
温度是影响污损生物群落结构的关键环境因子(平均贡献率为44.95%);盐度是影响污损生物群落结构的主要环境因子(平均贡献率为31.90%);溶解氧与网衣污损生物丰度、生物量呈负相关,但相关性较低,pH规律性不显著。
3 讨论
3.1 网衣污损生物群落组成
根据本研究结果,雷州湾网衣污损生物物种组成存在时空差异。1)从时间尺度看,节肢动物门、软体动物门及刺胞动物门在各个季节均有分布,其中节肢动物门端足目占据重要优势地位,冬季的网衣污损生物种类数较其他季节少。不同养殖海域污损生物的优势种类有很大的不同,如美国缅因州西部海洋水产养殖的软体动物贻贝为优势种[22];舟山海域水螅虫(Hydravulgaris)、礁膜(Monostroma)、孔石莼(Ulvapertusa)、贻贝(Mytilusedulis)等为优势种[23];地中海西南部养殖场水螅和藻类为优势种[24];克罗地亚养鱼场藻类为优势种[25]。也有研究学者对海域沿岸进行污损生物现场调查,如中国东海西南沿岸海域污损生物研究调查的优势种为麦秆虫(Caprellaequilibra)、中胚花筒螅(Ectopleuracrocea)、海藻等[26];中国北部湾优势种为牡蛎和藤壶[27];波斯湾哈尔克岛优势种为节肢动物门和软体动物门[28]。综上结果,软体动物、节肢动物、藻类、刺胞动物出现频率高,具有显著优势,与挂网材质相关性不大,本文研究结果也符合这一生态规律。网衣污损生物附着基底为网衣,有别于船底、浮标,其中端足目因个体小在挂网早期易附着于网衣上,软体动物在幼虫阶段也易附着网衣上,并且生长迅速,在挂网后期形成体型大、生物量占优势的物种,但本研究中藻类出现频率并不为最显著,考虑其与季风气候有关,高温不利于藻类生存,但具体原因还有待进一步深入探究。2)从空间尺度看,上层29种,下层32种,表明两水层种类数相差不大,与Amini N等[28]研究指出的污损生物生物量随时间变化、不随空间变化的规律相一致。Ramadan S E等[29]对亚历山大港东部港口的4个10年(1960—1999)进行了比较研究,表明海洋污损生物多样性范围在19~35个分类单元之间,报告了El-Dekhaila港19 种海洋污染物种[30],与Ramadan S E等[29]研究结果相比,本研究的污损生物种类数接近,而与黄桂芳等[31]研究的污损生物总数154种相距甚远,但主要门类较一致,考虑本次所选取的实验材料较黄桂芳等[31]的少,具体的种类鉴定可能受表型可塑性影响,因而存在差异。综合时空生物多样性特征,本次实验表明雷州湾海域网衣污损生物生多样性高且具有一般水域的普遍特征,日后应注重对网衣的节肢动物门及软体动物门的防除探索。
3.2 网衣污损生物群落多样性
本研究还发现,从时间尺度看,春、夏、秋季的多样性指数(H′)、丰富度指数(d)差异不显著,且这3个季节平均值均显著高于冬季,但均匀度指数(J)各个季节差异不显著,表明春、夏、秋季的生物多样性较冬季更为复杂。从空间尺度看,上下水层网衣污损生物群落多样性指数(H′)、丰富度指数(d)、均匀度指数(J)差异不显著,表明本次取样水深0.5 m、4.5 m的污损生物生长状况差异不大。在日后深水养殖探究中,可对更深层海域做网衣污损生物多样性探究,为深水网箱养殖提供更多的参考数据。
3.3 环境因子对网衣污损生物群落分布的影响
网衣污损生物群落结构及其变化是多种环境因子在时间和空间序列上共同作用的结果,环境变化必然导致网衣污损生物群落结构和优势种的变化,而在众多环境因子中,存在起主导作用的因子[32]。本研究RDA分析结果显示,温度和盐度是影响网衣污损生物群落季节分布的主要因素,温度变化直接影响网衣污损生物的附着生长,多样性指数分析显示,春、夏、秋季较冬季更适合污损生物的生长,钩虾类和藤壶等物种生物量和丰度显示为春、夏季占据重要优势地位,斑马纹贻贝和翡翠贻贝的生物量在夏、秋季为优势种,藻类石莼、浒苔、石花菜等均显示春季较显著,这与严涛等[33]对黄渤海污损生物生态的研究结果相一致,但本文雷州湾礁膜、石莼等海藻在冬春季旺盛附着,而黄渤海礁膜、石莼等海藻一般都在温度比较高的月份旺盛附着,这可能与两地纬度差异有关。此外,Dziubińska A等[34]研究也指出污损生物群落演替受季节性影响,表明高温季节更适宜污损生物生长。RDA和丰度、生物量与环境因子的变化情况分析均显示,加尔板钩虾、笔螅、巴西地钩虾等附着生物与温度呈正相关,再进一步说明了温度对污损生物的生长响应显著。盐度是影响污损生物分布的另一关键因素,不同盐度区域污损生物的种类组成有很大的差别[35]。泥藤壶(Fistulobalanuskondakovi)适应河口低盐水域,而网纹藤壶等则在东海和南海的高盐海域出现,若水体盐度降至18甚至更低,则会妨碍网纹藤壶幼虫的发育,甚至导致其死亡[36]。在本研究中,网纹藤壶附着量较大且没有泥藤壶出现,这可能与所处海域的盐度较高密切相关;春、秋季盐度较低,受盐度和温度的复合影响,这两季的生物多样性也较为相似。
3.4 网衣污损生物管理
根据污损生物的分布特点,合理管理网衣更替,可最大限度地降低养殖成本,保证生产安全、高效。为此,建议在春、夏、秋季养殖期间提高换网频率,冬季则可适当地降低换网频率;更换的网衣应注重清理节肢动物门和软体动物门污损生物,可采用浸泡法清除节肢动物、物理冲刷法加暴晒敲打法清除软体动物,建议积极开展探索低温环保材质的网衣制作工艺,以期为绿色养殖中污损生物的防治提供参考依据。
