小麦源库优异种质的鉴定与源库类型的划分
2023-04-11南瑞杨玉存石芳慧张礼宁米彤茜张立强李春艳孙风丽奚亚军张超
南瑞,杨玉存,石芳慧,张礼宁,米彤茜,张立强,李春艳,孙风丽,奚亚军,张超
小麦源库优异种质的鉴定与源库类型的划分
南瑞1,杨玉存1,石芳慧1,张礼宁1,米彤茜1,张立强2,李春艳2,孙风丽1,奚亚军1,张超1
1西北农林科技大学农学院/农业农村部西北地区小麦生物学与遗传育种重点实验室,陕西杨凌 712100;2宝鸡市陇县种子工作站,陕西宝鸡 721200
【目的】筛选源库评价指标,划分试验材料的源库类型,探究源库关系对小麦农艺、产量及品质性状的影响,为小麦源库代谢研究和遗传育种应用提供参考。【方法】以国内外190份小麦种质资源为试验材料,测定其源代谢、库代谢相关性状,利用主成分分析对小麦的源库代谢能力进行评价,根据综合得分对优异小麦材料进行筛选。以源活性综合得分、源大小(叶面积)、库活性综合得分、库数目(穗粒数)为指标进行层次聚类,根据聚类结果总结小麦材料的源库类型,分析不同区域小麦源库特征差异,比较不同源库类群间小麦的农艺性状、产量性状、品质性状的差异。【结果】通过主成分分析将6个与小麦源活性密切相关的指标转化为3个独立指标(光化学淬灭系数、最大光合潜力和叶绿素含量),将5个与库活性密切相关的指标转化为2个独立指标(最大灌浆速率和灌浆持续期),累计贡献率分别为82.80%和92.90%,筛选出源活性、源大小(叶面积)、库活性、库数目(穗粒数)排名前十的小麦品种。根据源库关系,将供试的190份小麦材料划分为三大类八小类:源足库乏型(源中库弱型、源强库中型)、源乏库足型(源中库强型、源弱库中型)和源库平衡型(受限于库活性的源弱库弱型、受限于源活性和穗粒数的源弱库弱型、源中库中型、源强库强型),其中,受限于源活性与穗粒数的源弱库弱型、源中库强型、源强库强型包含了大多数品种,占所有材料的76.84%。国内多数小麦品种源库关系相近,源活性、叶面积、穗粒数处在中等水平,库活性较高,长江中下游冬麦区品种库活性较低。不同类群间,株高、穗下节长、小穗数表现为源足库乏型>源库平衡型>源乏库足型,籽粒蛋白质干基、湿面筋干基、沉降值总体表现为源足库乏型>源库平衡型>源乏库足型,籽粒吸水率表现为库活性越强吸水率越高,单株产量在不同源库类群间表现不一,但在品种较多的三类中单株产量与源活性和穗粒数皆呈正相关。【结论】光化学淬灭系数、最大光合潜力、叶绿素含量可作为评价小麦源活性的主要指标,最大灌浆速率、灌浆持续期可作为评价小麦库活性的主要指标。在实际生产中可通过提高穗粒数和源活性的方式提高小麦产量,源的供应能力相对强于库的吸收能力将促进小麦株高、穗下节长、小穗数、蛋白质干基、湿面筋干基、沉降值的增加,较强的库活性有助于提高小麦籽粒的吸水率。
小麦;评价指标;源库类型;农艺性状;产量性状;品质性状
0 引言
【研究意义】近年来,人口的增加和极端气候条件的频发引发了人们对未来世界粮食安全的担忧[1-2],小麦作为人类碳水化合物和蛋白质的主要来源[3],其产量和品质的提高是当前研究的重点,有学者认为,未来作物生产力提高的关键在于研究源和库对作物生长的限制程度及内在机理[4-5],而作物源库类型的划分与优异材料的筛选则是相关研究的基础。因此,对不同小麦品种的源库代谢能力进行评价,筛选源库鉴定指标,对品种的源库类型进行划分将为小麦新品种选育和源库理论的完善提供种质基础与理论参考,对小麦产量和品质的提高具有重要意义。【前人研究进展】“源库”理论概念在1928年被Mason和Maskell首次提出,这一理论为作物高产提供了新的思路和方法。对于小麦而言,源主要指绿色的叶、茎、鞘以及根系[6],库主要指新生的组织和籽粒[7],在生育后期,旗叶是最关键的源[8-9],籽粒是最主要的库,不同小麦品种间源库特征差异显著[10-12],表明适合进行品种的筛选与分类研究。近年来,国内小麦综合评价研究及品种筛选主要集中在抗逆性[13-15]、抗病性[16-17]、资源利用效率[18-19]和品质[20-21]等方面,对小麦源库能力的评价以及源库类群划分的研究相对较少。20世纪80—90年代,曾有学者进行过小麦源库类型区分的研究,如:有学者依据源库结构和开花前后光合积累速率的变化将小麦划分为前期优势型、均势型和后期优势型[22];或根据源流库的协调状况,将麦田区划分为受限于流的类型、受限于源的类型和受限于库的类型[23],也有学者认为凡穗花数较多、不结实小花数较多的为源限制型,穗花数较少,不结实小花数较少的为库限制型[24],这对当代小麦源库类型的划分具有一定的借鉴作用。【本研究切入点】目前,对小麦源库关系的研究多集中在非生物胁迫以及人为改变源库特征对源库关系的影响[25-28],且涉及的品种数目偏少,未见大批量作物品种源库类型划分的研究。近年来,国内外育成了很多优良小麦品种,但其源库关系尚不明确,且在当前育种条件下,小麦产量与品质的提升逐渐无法满足人们的需求,重新从源库代谢的角度进行品种筛选和相关研究变得尤为必要。【拟解决的关键问题】本研究以国内外190份小麦种质资源为试验材料,筛选源、库评价主要指标,划分供试材料源库类型,探究源库关系对小麦农艺、产量及品质性状的影响,为小麦源库代谢研究和遗传育种应用提供参考。
1 材料与方法
1.1 供试材料
190份国内外小麦种质资源,国内品种114份,国外品种76份。国内品种根据其种植省份或审定情况进行麦区划分,长江中下游麦区品种8份,黄淮冬麦区品种74份,西南春麦区品种6份,西南冬麦区品种1份,北部冬麦区品种4份,新疆冬麦区品种1份,西藏麦区品种1份,种植麦区信息不明确的其余国内品种19份。种子由农业农村部西北地区小麦生物学与遗传育种重点实验室提供,其名称、生育期、冬春性、来源地、系谱等具体信息见电子附表1。
1.2 试验设计
试验于陕西省咸阳市杨凌区国家(杨凌)旱区植物品种权交易中心进行,试验场地为钢架旱棚,通风性良好,前茬作物为玉米,秸秆粉碎后旋耕用作基肥,施底肥小麦复合肥(N-P2O5-K2O)752 kg·hm-2,所有材料于2021年10月20日种植,采用人工点播的方式,每份材料种植2行,2次重复,行长1 m,行距0.25 m,株距6.6 cm,在小麦生长期内,根据田间土壤水分情况及时浇水,其余田间管理与高产田一致,2022年6月17日收获。
1.3 测定项目与方法
1.3.1 田间表型测定 以50%以上麦穗中上部小花的内外颖张开为标准,准时记录其开花期,根据公式(5)统计小麦成熟期。于小麦开花后15—20 d,采用SPAD-502型叶绿素计(Minolta Camera,日本)测定小麦SPAD值,测量时选取长势一致且无病虫害植株的小麦旗叶,每个品种测量5个叶片,分别测量叶中、叶尖和叶基SPAD值,取平均值表示该叶片的相对叶绿素含量。在测定完叶片SPAD值后的夜间,采用FluorPen FP110手持式叶绿素荧光仪(Photon Systems Instruments, Brno, Czech Republic)测定叶绿素荧光参数,选取健康植株的旗叶,每个品种测量3个叶片,测量叶片中部,使用该设备中的NPQ1程序进行测量,可以得到v/m(qy)、v’/m’(qy_lss)、NPQ、qP(qp_lss)、m’(fm_lss)等重要参数,取平均值用于表示该指标。灌浆期测定小麦旗叶叶面积,每个品种选取10株测量叶长、叶宽,根据公式(1)计算叶面积。
1.3.2 室内性状测定 小麦开花时,每个品种选择花期一致,无病虫害的单穗挂牌标记,开花10 d后开始取样, 此后每5天取样1次,直至小麦完熟,每次取样3穗,全部取出籽粒,调查每穗粒数,以每次取样的穗粒数平均值表示穗粒数。籽粒于105℃处理15 min后,80℃烘干至恒重,称量干重,换算成千粒重,计算灌浆速率。小麦成熟后每个品种取5株进行室内考种,分别调查株高、穗下节长、有效穗数、千粒重和单株产量。用DA7250型近红外漫反射谷物品质分析仪(Perton,瑞典)分析小麦品质性状,用万深SC-G自动考种及千粒重仪(杭州万深检测科技有限公司)测定籽粒面积。
1.3.3 数据计算
叶面积=叶长×叶宽×0.75 (1)
灌浆持续期(grain filling duration,GFD)=2-1(2)
平均灌浆速率(average filling rate,AFR)=W/GFD(3)
最大灌浆速率(maximal filling rate,MFR)=b1+2b2tmax+3b3tmax2(4)
成熟期=开花期+2(5)
具体计算过程如下:小麦籽粒生长动态符合Logistic方程以及三次多项式方程,有学者认为多项式方程比Logistic方程在拟合籽粒灌浆过程中具有更好的拟合优先度[29-30],因此,采用多项式方程对籽粒生
长动态进行拟合。三次多项式方程的数学表达式为
()=b1+b2t+b3t(6)
式中,()为粒重,为小麦开花后的天数,b1、b2、b3分别为模拟方程的系数,由于品种过多,部分品种拟合度较差,为防止部分品种灌浆参数产生过大误差,对传统方法进行一定的改动,主要对灌浆起始时间的计算进行了修正,根据小麦灌浆起始时间在开花后3—5 d,在进行数据拟合时设定当=0、0.5、1、1.5、2时,()=0。
GFD:令()=0,求得3个解,选择数值处在中间的解t1作为灌浆起始时间,令()=0,求得2个解,选择数值较大的解作为灌浆终止时间,GFD=2-1。
AFR:将灌浆终止时间2代入(6)式,求得理论最大粒重W,AFR=W/GFD。
MFR:对公式(6)求导即得灌浆速率方程(7):
()=b1+2b2+3b32(7)
对(6)式求导并令导数为0,求得灌浆速率极大值出现时间tmax=-b2/3b3将tmax代入(7)式求得MFR= b1+2b2tmax+3b3tmax2。
1.3.4 数据处理 采用Microsoft Excel软件计算品种间各指标最大值、最小值、平均值、标准差、变异系数以及遗传多样性指数(Shannon-Weaver多样性指数)[31],采用SPSS 26.0软件进行主成分分析和方差分析,采用Tbtools[32]进行热图聚类分析。
2 结果
2.1 供试小麦品种源库指标变异幅度及遗传多样性
将小麦的源库指标分为源性状和库性状,计算190份参试小麦材料各源库指标的最小值、最大值、变异系数和遗传多样性指数(表1),多数源性状的变异系数大于0.26,多样性指数为1.83—2.05,库性状的变异系数为0.1—0.19,多样性指数为2.02—2.09,整体而言,除v/m外,源性状的变异系数大于库性状,而多样性指数则小于库性状,表明小麦源性状变异幅度更大,而库性状在不同等级内的分布更加均匀,可能是由于在育种的过程中育种家更加倾向于对库性状进行选择[33],将其规定在一定的范围内,而忽略了对源性状的选择所造成的。v/m的变异范围为0.81—0.87,变异系数仅为0.01,多样性指数也较小,表明不同品种小麦在灌浆期内旗叶的最大光合潜力基本稳定,其余源库指标的变异系数及遗传多样性指数均较大,表明所选小麦材料源库性状变异程度大且遗传多样性丰富,适合进行品种的筛选与相关分析。
2.2 主成分分析
2.2.1 源活性主成分分析 主成分分析可通过对多个指标进行降维的方式,防止在整体分析时由于多个指标的信息重叠而造成结果失真,当指标反映问题不在同一维度时则没有必要进行主成分分析。源性状中,SPAD值、v’/m’、v/m、NPQ、qP、m’等6个指标可反映小麦源活性的不同特征,叶面积主要反映小麦的源大小,因此,对SPAD值等6个与小麦源活性密切相关的指标进行主成分分析,在特征值大于1的条件下,可将原本的6个单项指标转换为3个新的主成分。第一主成分特征值为2.44,贡献率为40.62%,第二主成分特征值为1.52,贡献率为25.37%,第三主成分特征值为1.01,贡献率为16.81%,累积贡献率达到82.80%,反映了原始数据的大多数信息(表2),因此,可以选取这3个指标对小麦源活性进行综合分析。第一主成分中qP的特征向量值最大,第二主成分中v/m特征向量值最大,第三主成分中SPAD特征向量值最大(表3),qP反映的是PSⅡ天线色素吸收的光能用于光化学电子传递的份额,可将第一主成分概括为有效光合因子,v/m反映植物最大光合潜力,可将第二主成分概括为光合潜力因子,SPAD反映植物叶绿素含量,可将第三主成分概括为叶绿素因子。因此,将小麦源活性的主成分因子概括为有效光合因子、光合潜力因子、叶绿素因子。
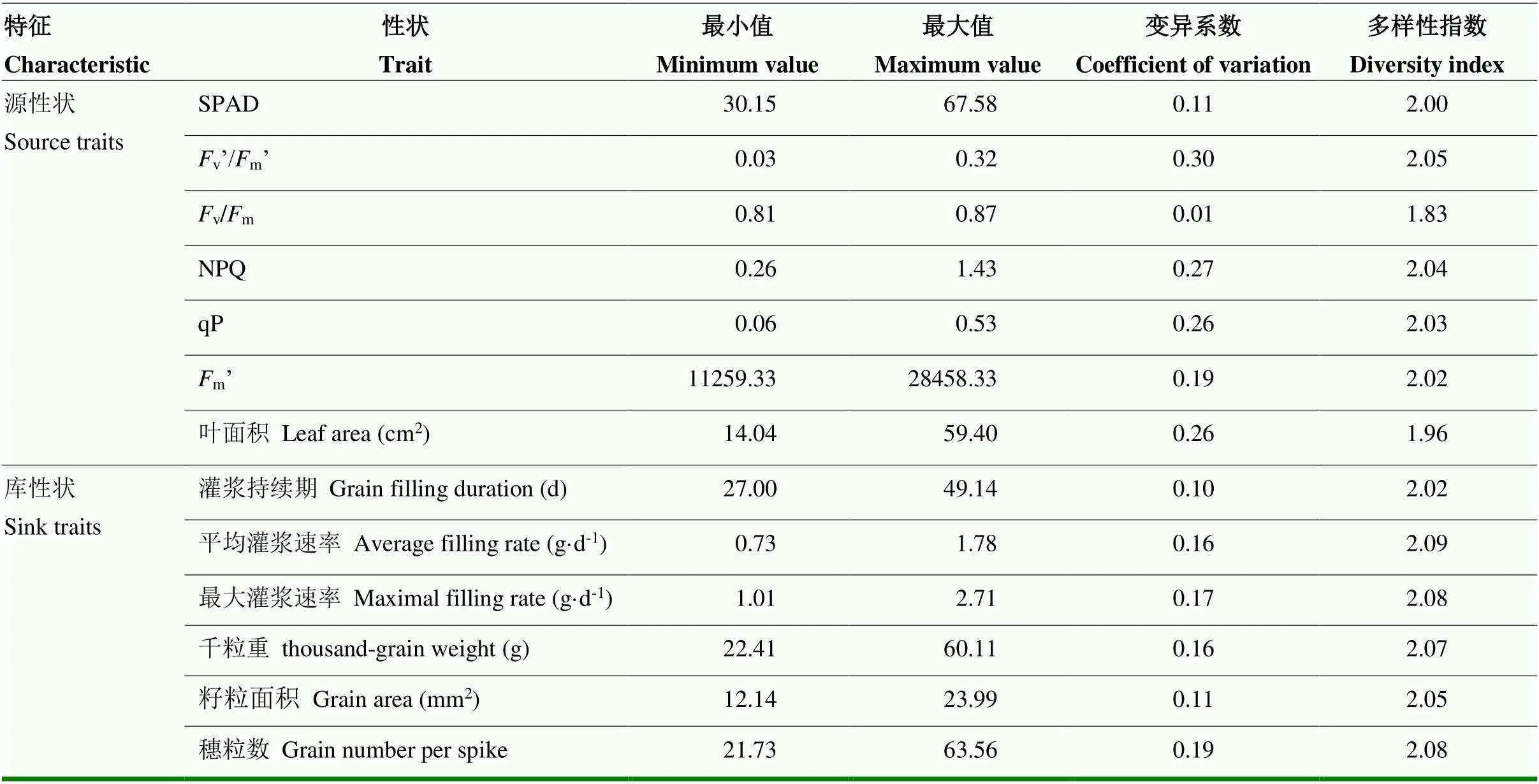
表1 小麦源库指标变异范围及遗传多样性指数
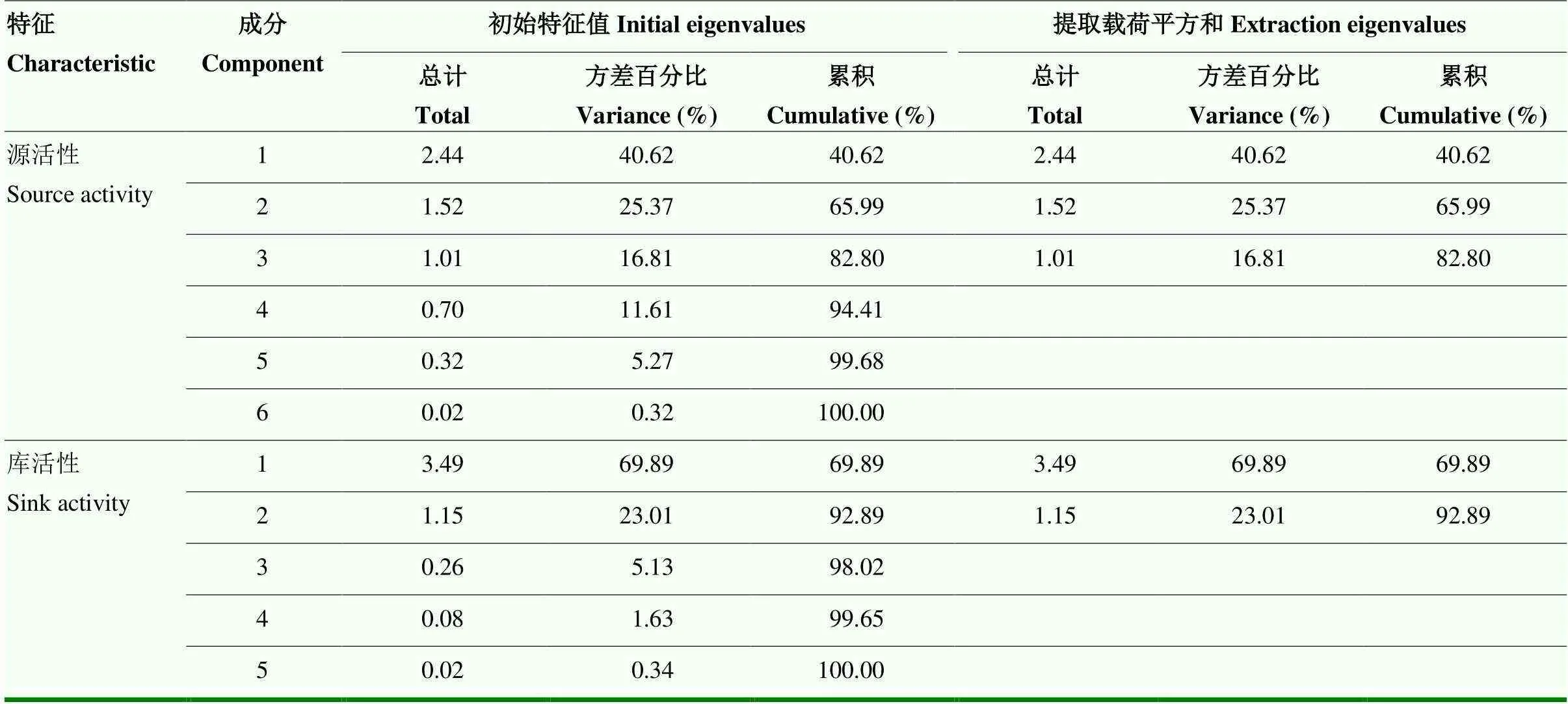
表2 小麦源活性、库活性主成分初始特征值、方差贡献率与累积贡献率

表3 小麦源活性、库活性的主成分载荷矩阵
利用各主成分计算综合得分,以综合得分评价小麦源活性,综合得分越高表明源活性越强。以3个主成分对应的方差贡献率为权重,得到综合得分计算公式为:C-Source=40.62%C1+25.37%C2+16.81%C3,综合得分排名前十的品种分别为:IR39、红粒无名、西农1376、印度蓝粒、MK186、豫麦54、SIRMIONE、西农100、开麦21和疙绉麦(表4),灌浆期小麦旗叶叶面积与小麦的源大小息息相关,因此,以旗叶叶面积来表示小麦源大小,叶面积排名前十的品种分别为:GRIFONE 235、LAGODEKHIS-GRDZEL TAVTAVA、H 798、LOHARI Y91-92 NO.123、西昌76-9、RED-BOBS-222、中国春、Claudia、MK186和SENMARQ(表5),上述品种可作为小麦育种的种质材料。
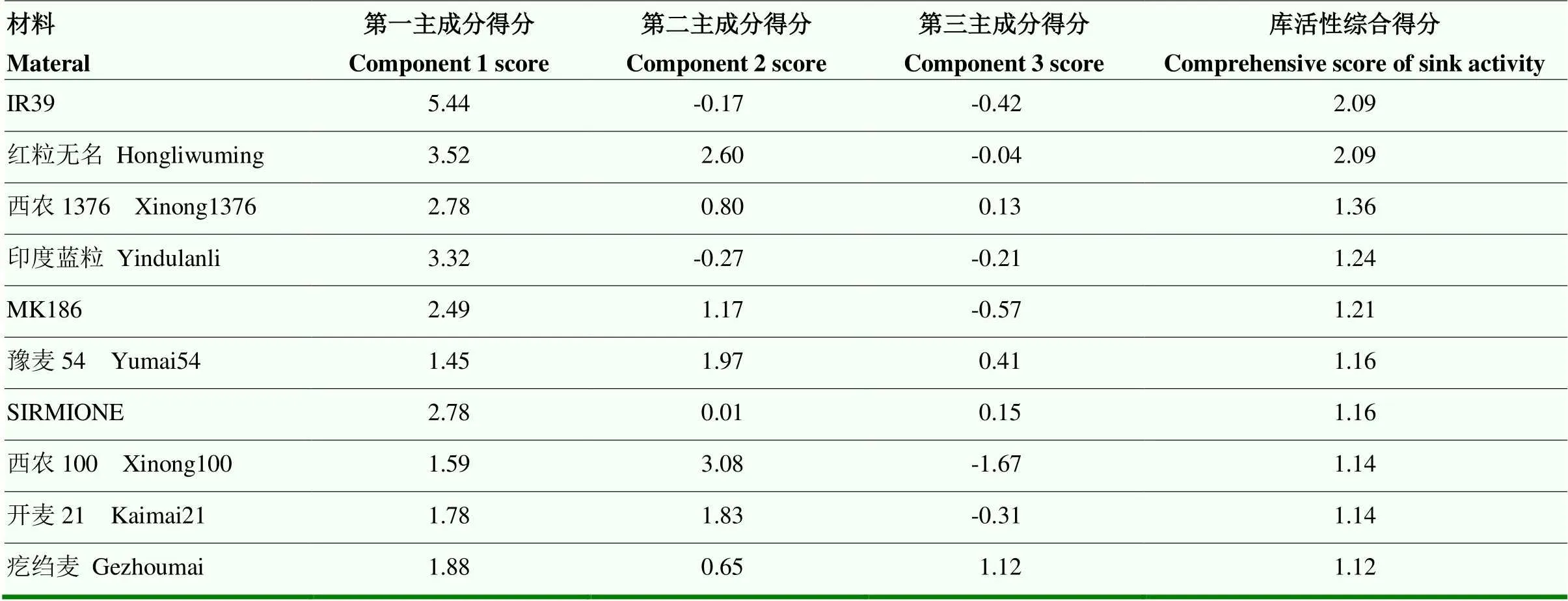
表4 源活性排名前十的小麦品种及其各主成分得分

表5 源大小排名前十的小麦品种
2.2.2 库活性主成分分析 库性状中,灌浆持续期、平均灌浆速率、最大灌浆速率、千粒重和籽粒面积反映小麦库活性的不同特征,穗粒数反映库数目,因此,对灌浆持续期等5个与库活性密切相关的指标进行主成分分析。在特征值大于1的情况下可将原本5个指标转化为2个新的主成分,第一主成分特征值为3.49,贡献率为69.89%;第二主成分特征值为1.15,贡献率为23.01%,累积贡献率达到92.90%,反映了原始数据绝大多数的信息(表2),因此,可以选取这两个主成分对小麦库活性进行综合分析。第一主成分中最大灌浆速率的特征向量最大,其次为平均灌浆速率;第二主成分中灌浆持续期的特征向量值最大(表3),因此,可将第一主成分概括为灌浆速率因子,将第二主成分概括为灌浆时长因子,表明小麦库活性的主成分因子可分为灌浆速率因子和灌浆时长因子。
利用主成分计算综合得分,利用综合得分评价小麦库活性,综合得分越高表明库活性越强。以3个主成分对应的方差贡献率为权重,可得出综合得分计算公式为C-Sink=69.89%C1+23.01%C2,综合得分排名前十的品种为开麦21、豫麦19、西农928、中麦578、宁麦12、FS056、周麦26、内乡182、豫农949、丰抗8号(表6),小麦穗粒数可代表库的数目,穗粒数排名前十的品种为:洲元9369、Claudia、YDSS、红粒(zh)、焦麦266、IR89、IR202、西昌76-9、IR4和IR211(表7)。上述材料可作为小麦育种的种质材料。
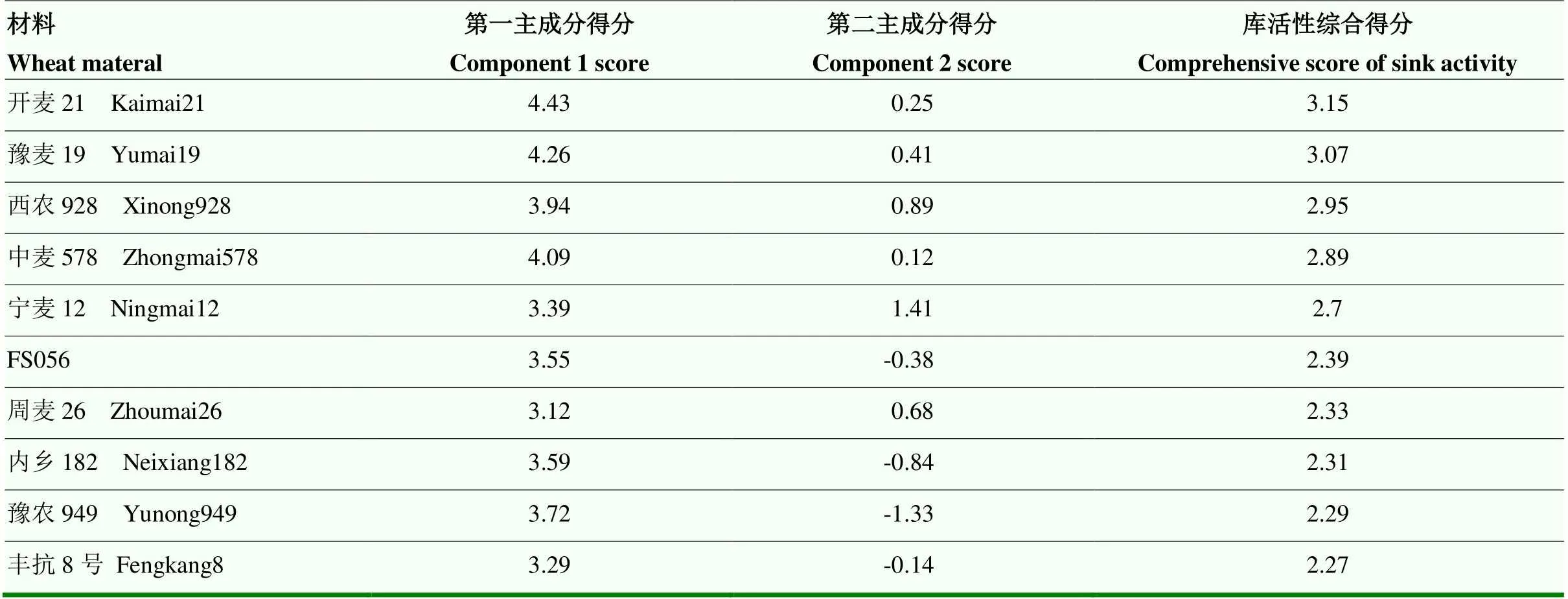
表6 库活性排名前十的小麦品种及其各主成分得分
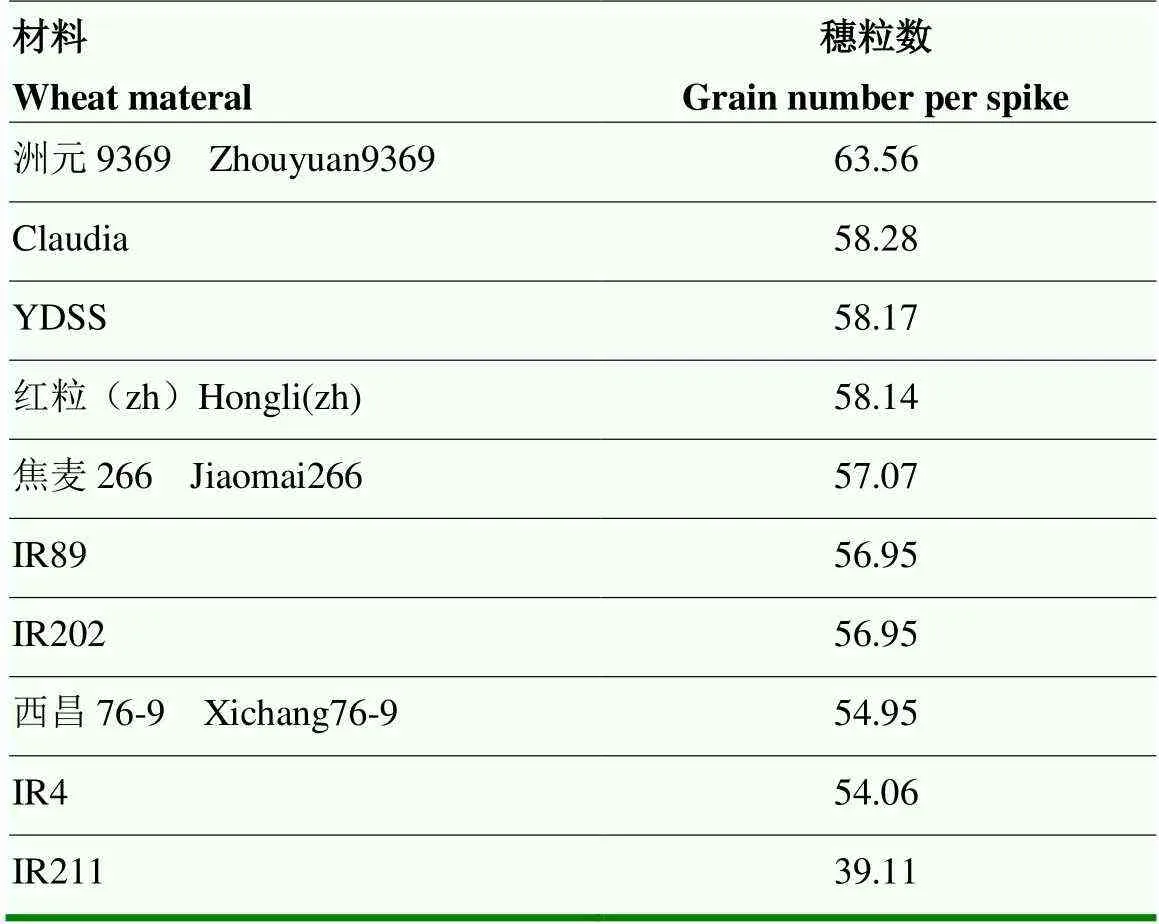
表7 库数目排名前十的小麦品种
2.3 聚类分析
以小麦源活性综合得分、旗叶叶面积、库活性综合得分和穗粒数为指标,采用欧式距离进行层次聚类,构建树状聚类图,聚类热图的不同颜色反映数值的相对大小。
2.3.1 小麦源库类型的划分 从图1中可以看出,190份供试小麦材料被划分为8个类群,利用SPSS对8个类群源活性、叶面积、库活性和穗粒数进行方差分析,类群间差异显著(表8),说明聚类结果合理,总结各类群源库特征。
第Ⅰ类群:包含19个品种,占供试材料的10%。该类群主要特点是源活性中等,叶面积中等,库活性较低,穗粒数较高,总之,源供应同化物的能力和库吸收同化物的能力都较匮乏,库活性是限制植株源库代谢的主要因素,因此,可将其归纳总结为受限于库活性的源弱库弱型。对于该类群,库活性提升空间较大,在育种工作中,可首先通过提高库活性来增强其源库代谢能力。
第Ⅱ类群:包含3个品种,占供试材料的1.58%。该类群主要特征是源活性高,叶面积小,穗粒数较高,库活性较低,总之,源供应同化物的能力和库吸收同化物的能力都处在中等水平,叶面积和库活性共同限制了植株的代谢能力,因此,可将其归纳总结为受限于叶面积与库活性的源中库中型,对于该类群,叶面积和库活性提升空间都较大,在育种工作中可通过提高叶面积和库活性增强其源库代谢能力。
第Ⅲ类群:包含4个品种,占供试材料的2.11%。该类群主要特征是源活性低,叶面积小,库活性高,穗粒数较低,总之,源供应同化物的能力较为匮乏,库吸收同化物的能力相对充足,源活性和叶面积都是限制植株代谢能力的主要因素,源代谢强度整体较低,因此,可将其归纳总结为受限于源强度的源弱库中型。对于该类群,在育种工作中可通过提高叶面积和源活性来有效增强其源库代谢能力。
第Ⅳ类群:包含15个品种,占供试材料的7.89%。该类群主要特点是源活性中等,叶面积中等,库活性低,穗粒数低,总之,库吸收同化物的能力很匮乏,源供应同化物的能力相对充足,库活性和穗粒数都是限制植株代谢能力的主要因素,库代谢强度整体较低,因此,将该类群总结为受限于库强度的源中库弱型。对于该类群,在育种工作中可通过提高穗粒数和库活性来有效增强其源库代谢能力。

图1 190份小麦材料的源库聚类结果
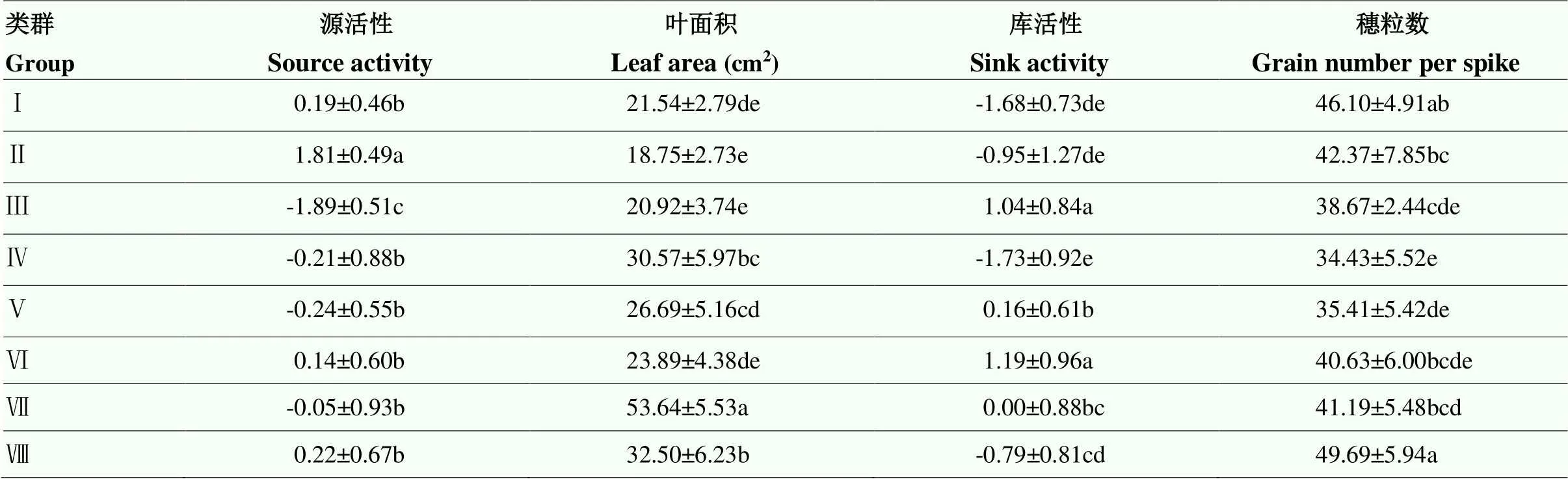
表8 190份小麦材料在8个类群的分布
不同小写字母表示材料间达到显著水平(=0.05)。下同
different lowercase letters indicate a significant level between materials (=0.05). The same as below
第Ⅴ类群:包含56个品种,占供试材料的29.47%。该类群主要特点是源活性较低,叶面积中等,库活性中等,穗粒数较低,总之,源供应同化物的能力和库吸收同化物的能力都处在中等水平,源活性和穗粒数共同限制了植株的代谢能力,因此,可将其归纳总结为受限于源活性和穗粒数的源弱库弱型,对于该类群,在育种工作中可通过提高源活性和穗粒数增强其源库代谢能力。
第Ⅵ类群:包含59个品种,占供试材料的31.05%。该类群主要特点是源活性中等,叶面积中等,库活性高,穗粒数中等,总之,该类群库吸收同化物的能力很强,源供应同化物的能力相对匮乏,源活性和叶面积共同限制了植株的代谢能力,因此,将该类群总结为受限于源强度的源中库强型。对于该类群,在育种工作中可通过提高源活性和叶面积来使其源库代谢能力近进一步增强。
第Ⅶ类群:包含3个品种,占供试材料的1.58%。该类群主要特点是源活性中等,叶面积大,库活性中等,穗粒数中等,总之,源供应同化物的能力很强,库吸收同化物的能力相对匮乏,库活性和穗粒数共同限制了植株的代谢能力,库代谢强度整体相对较低,因此,将类群归纳总结为受限于库强度的源强库中型。对于该类群,在育种工作中可通过提高库活性和穗粒数来使其源库代谢能力近进一步增强。
第Ⅷ类群,包含31个品种,占供试材料的16.32%。该类群主要特点是源活性较高,叶面积较大,库活性较低,穗粒数很高,总之,源供应同化物的能力和库吸收同化物的能力都很强,库活性是限制植株的代谢能力的主要因素,因此,将该类群归纳总结为受限于库活性的源强库强型。对于该类群而言,在育种工作中可尝试增强其库活性来使其源库代谢能力更进一步。
综上所述,小麦源库类型丰富,从源库平衡的角度来看,上述聚类所得的8个类群包含了所有源库关系,第Ⅰ、Ⅱ、Ⅴ、Ⅷ类可归纳为源库平衡型,其余4个类群则都属于源库不平衡型,其中,第Ⅲ类和第Ⅵ类可归纳为源乏库足型,第Ⅳ类和第Ⅶ类可归纳为源足库乏型。从材料占比上来看,第Ⅴ、Ⅵ、Ⅷ类分别占到供试材料的29.47%、31.05%和16.32%,表明多数小麦品种属于受限于源活性与穗粒数的源弱库弱型、源中库强型和源强库强型,其中,第Ⅷ类所包含的31个小麦品种源库性状优异,且源、库供需情况较为均衡,可用于源库平衡方面的研究与利用。
2.3.2 不同麦区小麦特征分析 统计不同麦区及国外品种的开花时间、成熟时间,以及其在不同源库类别的分布情况。总之,参试品种中,不同麦区小麦品种的开花时间与成熟时间依次为长江中下游麦区、西南春麦区、黄淮冬麦区、北部冬麦区,参试的国外品种相较于国内品种开花时间与成熟时间跨度相对较大,平均开花时间与成熟时间相对较晚(表9),长江中下游麦区、西南春麦区多数品种为春性品种,黄淮冬麦区、北部冬麦区多为冬性或半冬性品种(电子附表1),表明不同麦区小麦的开花期与成熟期具有一定的差异,且小麦生育期与品种冬春性有一定的相关性,春性强的品种开花早,成熟早,冬性强的品种开花晚,成熟晚。根据不同麦区品种及国外品种在不同源库类别的分布情况可知,国外品种在不同小麦源库类别中的分布较为均匀,而中国麦区多数品种分布在第Ⅴ类源弱库弱型和第Ⅵ类源中库强型,其中,北部冬麦区品种、多数西南春麦区、多数黄淮冬麦区品种分布在第Ⅵ类,长江中下游麦区品种多数分布在第Ⅴ类(表9),表明中国品种与国外品种在源库类别上具有差异,中国多数品种源库关系相近,其源活性、叶面积、穗粒数中等,库活性较高(表8),说明中国品种源代谢能力稍弱,在育种过程中可通过增强源代谢能力探索高产新途径。长江中下游冬麦区品种与北部冬麦区、西南春麦区、黄淮冬麦区品种在源库关系上有一定的差别,根据第Ⅴ、Ⅵ类的差异可知,长江中下游冬麦区品种相比于其他麦区品种,具有更高的叶面积、更低的源活性、更低的库活性与穗粒数,库活性的差别最为显著,表明对于长江中下游冬麦区品种而言,增强源活性与库活性是提高其整体源库代谢能力的有效途径。
2.3.3 不同源库类群小麦农艺性状及单株产量表现情况 利用SPSS对上述8个类群株高、穗下节长、小穗数、千粒重、穗粒数、单株产量进行方差分析(表10),由表可知,不同源库类群间株高和穗下节长差异较显著,小穗数差异不显著,千粒重和穗粒数差异较高,单株产量差异较低。表明源库关系对株高、穗下节长、千粒重、穗粒数的影响较大,对小穗数和单株产量影响较小。
株高、穗下节长、小穗数表现为源足库乏型>源库平衡型>源乏库足型,表明当源的供应能力强于库的吸收能力时,剩余的同化物用于茎秆和穗下节的生长以及增加小穗数;从千粒重来看,第Ⅲ、Ⅵ类千粒重最高,表明源乏库足型小麦具有更高的千粒重;从穗粒数来看,第Ⅷ类穗粒数最高,表明受限于库活性的源强库强型小麦具有更高的穗粒数;从单株产量来看,第Ⅳ类最低,第Ⅶ类最高,且第Ⅳ类和第Ⅶ类的单株产量具有显著差异,第Ⅳ、Ⅶ类同属于源足库乏型,且都受到库强度限制,但第Ⅶ类的源库强度整体高于第Ⅳ类,这是第Ⅶ类高产的原因,表明源足库乏型小麦具有更高的产量潜力,受限于库强度的源强库中型小麦具有更高的单株产量。第Ⅴ、Ⅵ、Ⅷ类包含了大多数品种,代表性较强,三者的单株产量表现为第Ⅷ类>第Ⅵ类>第Ⅴ类,在这三类中源活性和穗粒数与单株产量皆呈正相关(表8),源活性主成分分析中,第一主成分占比40.62%,主成分中特征向量值最大的是光化学淬灭系数(qp),与源活性综合得分最为密切,表明对于多数小麦而言,提高叶片光化学淬灭系数和穗粒数是提高小麦单株产量的有效措施。
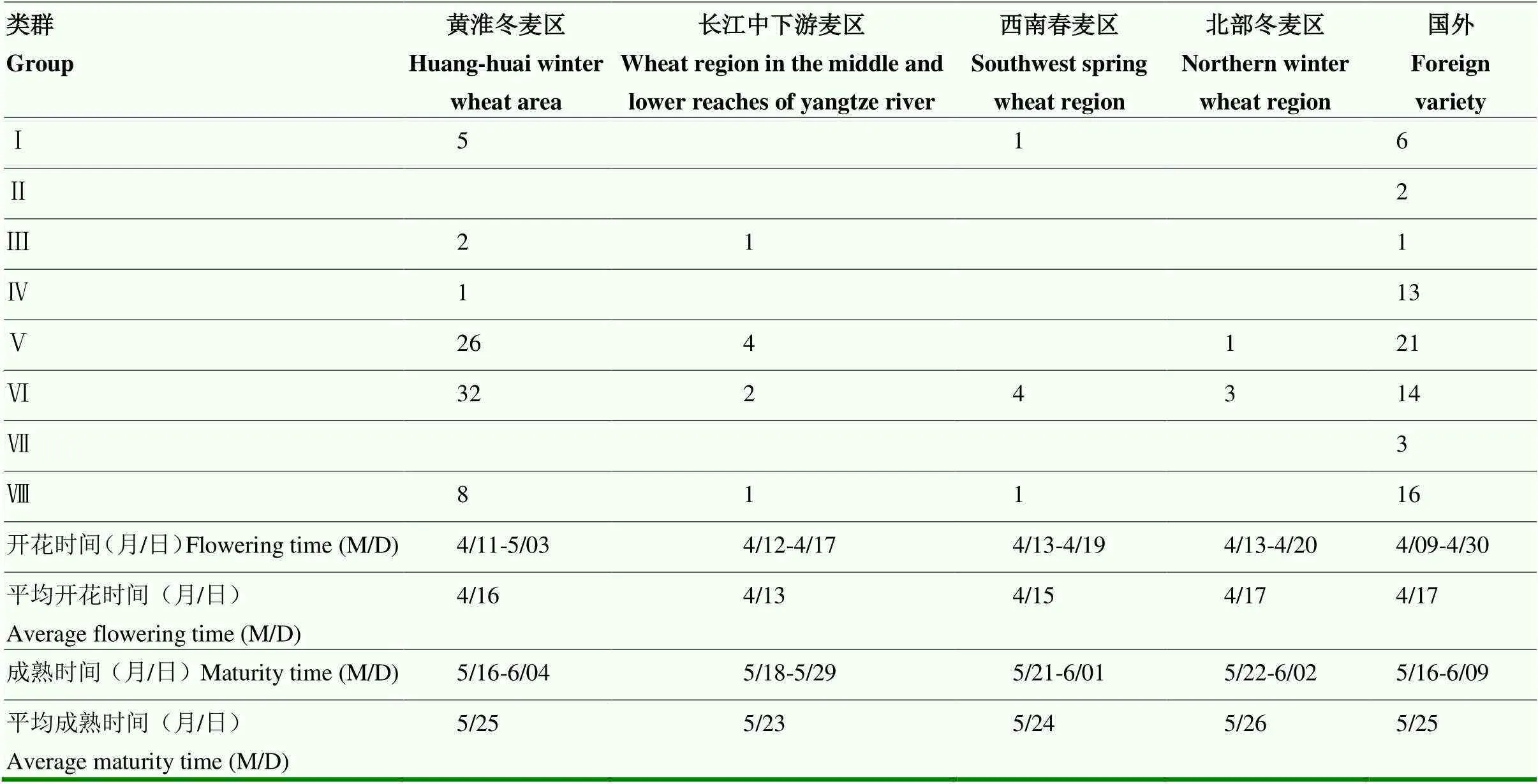
表9 不同区域小麦生育期及源库类型分布
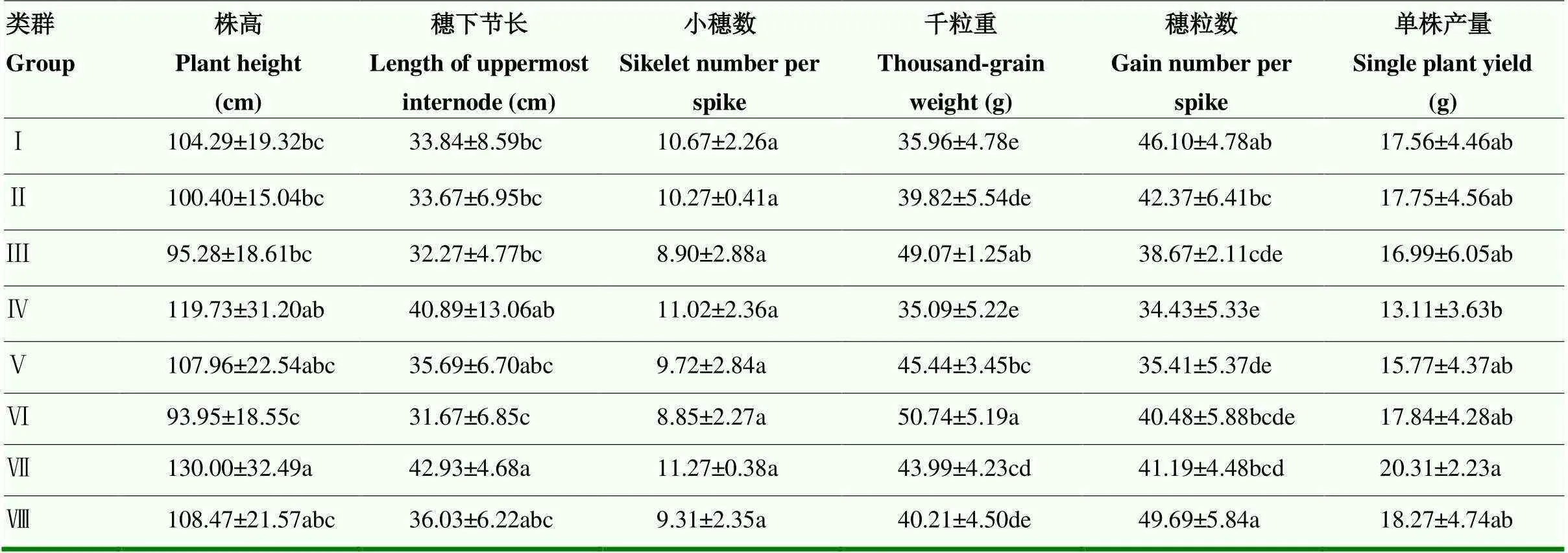
表10 不同源库类群小麦农艺及产量性状
2.3.4 不同源库类群小麦品质表现情况 由表可知,不同源库类群小麦蛋白质干基、水分、湿面筋干基、容重无显著差异,沉降值和吸水率具有一定的差异(表11),表明源库关系对籽粒蛋白质干基、水分、湿面筋干基和容重影响较小,对沉降值和吸水率有一定的影响。
蛋白质干基、湿面筋干基、沉降值大致表现为源足库乏型>源库平衡型>源乏库足型,吸水率大致表现为库活性越强沉降值越高,第Ⅳ类的整体品质表现最优,其蛋白质干基、水分、湿面筋干基在所有类群里面最高,沉降值在所有类群里面排第二位,吸水率最低,容重中等。表明源库关系影响小麦蛋白质含量、沉降值、湿面筋含量和吸水率,对沉降值和吸水率的影响较为显著,对容重几乎没有影响。当源的供应能力强于库的吸收能力时,小麦籽粒的蛋白质干基、沉降值、湿面筋会有一定程度的提高,吸水率会有一定程度的降低,但库活性对吸水率有正面的影响效果,相较于其他品种,受限于库强度的源中库弱型小麦的籽粒品质较为优良,在育种过程中选择库活性较高的源足库乏型小麦有利于小麦品质的提升。

表11 不同类群小麦品质性状
3 讨论
3.1 源库性状衡量指标
源库性状衡量指标是源库理论研究内容的重要组成,衡量源性状的指标主要包括净光合速率、胞间二氧化碳浓度、蒸腾速率、暗呼吸速率、初始羧化效率、叶片蒸腾速率、气孔导度、胞间二氧化碳浓度、叶绿素含量、叶绿素荧光参数等描述源活性的指标以及叶面积、叶面积指数等描述源大小的指标[34-39],衡量库性状的指标较少,过去的研究中提及的指标有开花后整株作物的光合生长率、花后生长持续期、库潜力、库强和库持续期等[40-41]。上述指标多为单一指标,有的研究人员以其为基础,衍生出一些反映源库综合能力的指标,如Wilson认为源强度是源大小与源活性的乘积,库容量是库活性与库大小的乘积[42],刘晓冰等[41]在1992年提出源强等于有效面积、源活性、绿叶持续期三者的乘积,由于这些指标的复杂性,在实际育种工作中并未被育种家们广泛运用。衡量源库性状的各类指标中,源活性与库活性含义及内容较为复杂,本研究针对二者进行了相关分析。以国内外190份小麦种质资源为试验材料,对叶绿素荧光参数、SPAD值等6个与源活性密切相关的指标和千粒重、平均灌浆速率等5个与库活性密切相关的指标分别进行主成分分析,累积贡献率分别达到82.80%和92.90%,反映了绝大多数信息,以各因子中特征向量大的指标描述小麦的源库活性,源活性主要分为有效光合因子(qp的特征向量值最大),光合潜力因子(v/m),叶绿素因子(SPAD),库活性主要分为最大灌浆速率因子(最大灌浆速率)和灌浆时长因子(灌浆持续期)。在诸多源代谢研究中,由于净光合速率等光合参数能直观反映干物质生产情况而被广泛应用,但光合参数的测量往往受环境制约较大而难以大批量测定,本研究采用了与光合作用密切相关但测量受环境因素制约较小的叶绿素荧光参数[43],荧光参数尽管不能像净光合速率一样直观地反映作物干物质生产情况,但可以更全面地反映光系统的量子活动情况,在反映作物光能利用情况方面具有优越性。
3.2 小麦源库类型的划分
在一定的生态及栽培条件下,对作物的源库类型进行划分,不仅可以完善作物源库理论,还有助于明确作物的育种方向。关于作物源库类型划分的研究,前人已取得了一定的进展。多数研究通过测量作物源、库的相关性状或对源库器官进行处理确定作物源库类型,如有学者认为凡穗花数较多、不结实小花数较多的为源限制型,穗花数较少,不结实小花数较少的为库限制型[24];有学者通过减源缩库后观察对应籽粒的变化动态的方式划分作物源库类型[44],也有学者对多个源性状、库性状利用主成分分析与聚类分析总结作物源库类型[45],此外,光合同化物从源到库的运输还需借助流这一途径,因此也有研究将流作为作物源库类型划分的标准之一[23];整体而言,上述划分作物源库类型的方式各有优劣,可根据实验目的和实验条件采取不同的方法。对于流是否是限制产量的因素,至今尚未有统一的定论,有研究证明,在小麦和水稻中,流并不是限制籽粒生长的因素[46-47],也有研究表明,维管束数目与灌浆强度、粒长、粒重、穗粒数、结实小穗数显著相关[48-49],本研究未将流纳入源库类型划分的影响因素中,可能会对结果造成一定影响。本研究选择了190份小麦材料,利用主成分分析评价源活性和库活性,利用源活性、源大小、库活性、库数目进行层次聚类,将小麦品种分为三大类八小类:源足库乏型(源中库弱型、源强库中型),源乏库足型(源中库强型、源弱库强型)和源库平衡型(受限于库活性的源弱库弱型、受限于源活性和穗粒数的源弱库弱型、源中库中型、源强库强型),中国麦区辽阔、品种繁多,但目前对小麦源库类型划分缺乏统一的标准与方法,不同的方法造成的结果可能有所差异。
3.3 源库性状对小麦农艺、产量及品质性状的影响
有机物在植物体内的分配受到源和库共同的调节,不同的分配方式对作物农艺、产量及品质性状都会造成一定的影响。在小麦和水稻中,前人通过人为改变源库性状的方式证明,当源的供应能力变强则光合产物向穗部分配减少,向茎鞘分配增多[50-51],较大的库容能促进小麦光合产物向穗部运输[39, 52],本研究显示,株高、穗下节长、小穗数在不同源库类群的表现遵循源足库乏型>源库平衡型>源乏库足型,表明在大多数小麦品种中,源的供应能力相对强于库的吸收能力时,光合产物向茎秆的运输增多且还可用于小穗数的增加。在过去,小麦产量的提高主要源自收获指数的提高,源是产量进一步提高的限制因素[53],水稻产量的提高主要源自穗粒数的提高[54],本研究显示,中国多数麦区小麦品种源库类型相近,源代谢能力稍差,可通过提高源代谢能力探索小麦高产新途径,尽管不同源库类群间育种改良方向不尽相同,但对于多数品种而言,提高源活性和穗粒数将有利于小麦单株产量的提高。源库性状的改变对作物品质也有一定的影响,对小麦而言,减源会降低小麦籽粒蛋白质含量,缩库则会增加小麦籽粒蛋白质含量[55-57],在玉米中,减源同样会降低玉米籽粒蛋白质含量[58],本研究蛋白质干基、湿面筋干基、沉降值大致表现为源足库乏型>源库平衡型>源乏库足型,吸水率表现为库活性越强沉降值越高,表明源的供应能力相对强于库的吸收能力有助于小麦籽粒蛋白质干基、湿面筋干基、沉降值的增加,较强的库活性有助于提高小麦籽粒吸水率。本研究在未进行人为源库关系调节的情况下,对源库关系与小麦农艺、产量及品质性状的关系进行研究,结论与前人得出的结论较为一致,使得部分结论的准确性与普适性得到提高,一定程度上丰富了小麦的源库理论。
4 结论
光化学淬灭系数(qP)、最大光合潜力(v/m)、叶绿素含量(SPAD)可作为评价小麦源活性的主要指标,最大灌浆速率、灌浆持续期可作为评价小麦库活性的主要指标;供试的小麦材料根据源库关系划分为8种源库类型:源足库乏型(源中库弱型、源强库中型)、源乏库足型(源中库强型、源弱库强型)和源库平衡型(受限于库活性的源弱库弱型、受限于源活性和穗粒数的源弱库弱型、源中库中型、源强库强型);国外小麦品种源库类别丰富,中国小麦品种源库类别相近,具有源活性、叶面积、穗粒数中等和库活性较高的特点;源的供应能力相对强于库的吸收能力将促进小麦株高、穗下节长、小穗数、蛋白质干基、湿面筋干基和沉降值的增加,较强的库活性有助于提高小麦籽粒吸水率;筛选出的源活性、源大小(叶面积)、库活性和库数目(穗粒数)排名前十的小麦品种可用于增强作物源库代谢能力方面的研究与利用,31份源强库强型小麦可用于源库平衡方面的研究与利用。
[1] YANG F, ZHANG J J, LIU Q E, LIU H, ZHOU Y H, YANG W Y, MA W J. Improvement and re-evolution of tetraploid wheat for global environmental challenge and diversity consumption demand. International Journal of Molecular Sciences, 2022, 23(4): 2206.
[2] LI A, LIU D, YANG W, KISHII M, MAO L. Synthetic hexaploid wheat: Yesterday, today, and tomorrow. Engineering, 2018, 4(4): 552-558.
[3] DE SOUSA T, RIBEIRO M, SABENÇA C, IGREJAS G. The 10,000-year success story of wheat!. Foods(Basel, Switzerland), 2021, 10(9): 2124.
[4] WHITE A C, ROGERS A, REES M, OSBORNE C P. How can we make plants grow faster? A source-sink perspective on growth rate. Journal of Experimental Botany, 2015, 67(1): 31-45.
[5] PAUL M J, WATSON A, GRIFFITHS C A. Linking fundamental science to crop improvement through understanding source and sink traits and their integration for yield enhancement. Journal of Experimental Botany, 2019, 71(7): 2270-2280.
[6] 赵全志, 黄丕生. 略论作物生产的源库系统行为. 耕作与栽培, 1998(6): 36-38.
ZHAO Q Z, HUANG P S. On the source-sink system behavior of crop production. Tillage and Cultivation, 1998(6): 36-38. (in Chinese)
[7] 马冬云, 郭天财, 宋晓, 查菲娜, 岳艳军. 小麦源库关系研究进展. 河南农业科学, 2008, 37(1): 12-15.
MA D Y, GUO T C, SONG X, ZHA F N, YUE Y J. Advances in research on source-sink relationship of wheat. Journal of Henan Agricultural Sciences, 2008, 37(1): 12-15. (in Chinese)
[8] BISWAL A K, KOHLI A. Cereal flag leaf adaptations for grain yield under drought: knowledge status and gaps. Molecular Breeding, 2013, 31(4): 749-766.
[9] CHEN X, LIU J Y, JIANG X D.Effect of source-sink change on photosynthesis and yield of wheat. Agricultural Science & Technology, 2010, 11(7): 59-61.
[10] 陈晓杰, 张建伟, 杨保安, 范家霖, 张福彦, 程仲杰, 胡银岗. 中国冬小麦微核心种质遗传多样性分析. 河南农业科学, 2015, 44(4): 14-20.
CHEN X J, ZHANG J W, YANG B A, FAN J L, ZHANG F Y, CHENG Z J, HU Y G. Genetic diversity of micro-core collections of Chinese winter wheat. Journal of Henan Agricultural Sciences, 2015, 44 (4): 14-20. (in Chinese)
[11] 卓武燕, 张正茂, 刘苗苗, 刘玉秀, 刘芳亮, 孙茹. 不同类型小麦光合特性及农艺性状的差异. 西北农业学报, 2016, 25(4): 538-546.
ZHUO W Y, ZHANG Z M, LIU M M, LIU Y X, LIU F L, SUN R. Difference of photosynthetic characteristics and agronomic traits in different types of wheat. Acta Agriculturae Boreali-Occidentalis Sinica, 2016, 25(4): 538-546. (in Chinese)
[12] 张凡, 刘国涛, 杨春玲. 620份小麦种质资源农艺性状调查及其遗传多样性分析. 山东农业科学, 2022, 54(3): 15-21.
ZHANG F, LIU G T, YANG C L. Agronomic characters and genetic diversity of 620 wheat germplasms. Shandong Agricultural Sciences, 2022, 54(3): 15-21. (in Chinese)
[13] 王小波, 关攀锋, 辛明明, 汪永法, 陈希勇, 赵爱菊, 刘曼双, 李红霞, 张明义, 逯腊虎, 魏亦勤, 刘旺清, 张金波, 倪中福, 姚颖垠, 胡兆荣, 彭惠茹, 孙其信. 小麦种质资源耐热性评价. 中国农业科学, 2019, 52(23): 4191-4200.
WANG X B, GUAN P F, XIN M M, WANG Y F, CHEN X Y, ZHAO A J, LIU M S, LI H X, ZHANG M Y, LU L H, WEI Y Q, LIU W Q, ZHANG J B, NI Z F, YAO Y Y, HU Z R, PENG H R, SUN Q X. Evaluation of heat tolerance of wheat germplasm resources. Scentia Agricultura Sinica, 2019, 52(23): 4191-4200. (in Chinese)
[14] 李龙, 毛新国, 王景一, 昌小平, 柳玉平, 景蕊莲. 小麦种质资源抗旱性鉴定评价. 作物学报, 2018, 44(7): 988-999.
LI L, MAO X G, WANG J Y, CHANG X P, LIU Y P, JING R L. drought tolerance evaluation of wheat germplasm resources. Acta Agronomica Sinica, 2018, 44(7): 988-999. (in Chinese)
[15] 王维领, 刘畅, 张雨婷, 段瑞华, 赵灿, 李国辉, 许轲, 霍中洋. 不同基因型小麦品种倒春寒抗性分析及其评价. 麦类作物学报, 2022, 42(10): 1220-1230.
WANG W L, LIU C, ZHANG Y T, DUAN R H, ZHAO C, LI G H, XU K, HUO Z Y. Analysis and evaluation of resistance to late spring coldness in different genotypes of whea. Journal of Triticeae Crops, 2022, 42(10): 1220-1230. (in Chinese)
[16] 黄亮, 刘太国, 肖星芷, 屈春艳, 刘博, 高利, 罗培高, 陈万权. 中国79个小麦品种(系)抗条锈病评价及基因分子检测. 中国农业科学, 2017, 50(16): 3122-3134.
HUANG L, LIU T G, XIAO X Z, QU C Y, LIU B, GAO L, LUO P G, CHEN W Q. Evaluation and molecular detection of stripe rust resistance in 79 wheat varieties (lines) in China. Scentia Agricultura Sinica, 2017, 50(16): 3122-3134. (in Chinese)
[17] 庞云星, 崔宏梅, 蔺瑞明, 杨慧, 冯晶, 马东方, 徐世昌, 王凤涛. 89份河北、山西小麦品种抗条锈性评价及抗条锈病基因检测. 植物保护, 2021, 47(6): 49-57, 92.
PANG Y X, CUI H M, LIN R M, YANG H, FENG J, MA D F, XU S C, WANG F T. Resistance evaluation and detection of the resistance genes in 89 wheat cultivars from hebei and shanxi provinces to stripe rust. Plant Protection, 2021, 47(6): 49-57, 92. (in Chinese)
[18] 赵瑞, 张旭辉, 张程炀, 郭泾磊, 汪妤, 李红霞. 小麦种质资源成株期氮效率评价及筛选. 中国农业科学, 2021, 54(18): 3818-3833.
ZHAO R, ZHANG X H, ZHANG C Y, GUO J L, WANG Y, LI H X. Evaluation and screening of nitrogen efficiency of wheat germplasm resources at adult stage. Scentia Agricultura Sinica, 2021, 54(18): 3818-3833. (in Chinese)
[19] 张子豪, 李想成, 吴昊天, 付鹏浩, 高春保, 张运波, 邹娟. 基于主成分和聚类分析筛选氮高效小麦品种. 湖南农业大学学报(自然科学版), 2022, 48(5): 513-519.
ZHANG Z H, LI X C, WU H T, FU P H, GAO C B, ZHANG Y B, ZOU J. Screening of wheat cultivars with high nitrogen efficiency using principal component and cluster analysis. Journal of Hunan Agricultural University (Natural Science), 2022, 48(5): 513-519. (in Chinese)
[20] 褚宏欣, 牟文燕, 党海燕, 王涛, 孙蕊卿, 侯赛宾, 黄婷苗, 黄倩楠, 石美, 王朝辉. 我国主要麦区小麦籽粒微量元素含量及营养评价. 作物学报, 2022, 48(11): 2853-2865.
CHU H X, MOU W Y, DANG H Y, WANG T, SUN R Q, HOU S B, HUANG T M, HUANG Q N, SHI M, WANG Z H. Evaluation on concentration and nutrition of micro-elements in wheat grains in major wheat production regions of China. Acta Agronomica Sinica, 2022, 48(11): 2853-2865. (in Chinese)
[21] 温艳霞, 张春娥, 廖若宇. 不同品种小麦的品质特性分析. 食品工业, 2022, 43(7): 299-302.
WEN Y X, ZHANG C E, LIAO R Y. Analysis of quality characteristics of different wheat varieties. Food industry, 2022, 43(7): 299-302. (in Chinese)
[22] 徐风. 小麦品种源库生态规律的研究(续): 我国小麦高产育种若干问题的探讨. 安徽农学院学报, 1985, 12(2): 1-15.
XU F. Study on the source-sink ecological law of wheat varieties (continued): Discussion on some problems of wheat breeding for high yield in China. Journal of Anhui Agricultural University, 1985, 12(2): 1-15. (in Chinese)
[23] 韩守良, 单玉珊, 慕美财, 张中兰, 王学芬, 孔凡克, 张道玺, 刘翠红, 孙庆英. 小麦超高产栽培理论探讨. 沈阳农业大学学报, 1999, 30(6): 576-580.
HAN S L, SHAN Y S, MU M C, ZHANG Z L, WANG X F, KONG F K, ZHANG D X, LIU C H, SUN Q Y. Discussion on the super high yield cultivation of winter wheat. Journal of Shenyang Agricultural University, 1999, 30(6): 576-580. (in Chinese)
[24] 王振林, 尹燕枰, 贺明荣, 孙学振. 小麦源库比与产量形成期同化物分配及结实性的关系. 山东农业大学学报, 1995, 26(2): 144-150.
WANG Z L, YIN Y P, HE M R, SUN X Z. Relationship of source-sink ratio and the partitioning of photoassimilates during the grain-filling phase with the grain setting of wheat. Journal of Shandong Agricultural University, 1995, 26(2): 144-150. (in Chinese)
[25] XU H, HASSAN M A, SUN D, WU Z, JIANG G, LIU B, NI Q, YANG W, FANG H, LI J, CHEN X. Effects of low temperature stress on source-sink organs in wheat and phosphorus mitigation strategies. Frontiers in Plant Science, 2022, 13: 807844.
[26] LIANG Y, CHEN Y X, LI D Q, CHENG J P, ZHAO G, FAHIMA T, YAN J. Selenium application effects on quality and distribution of trace elements in sink-source organs of wild emmer wheat. Journal of Animal and Plant Sciences, 2020, 31(1): 188-202.
[27] MADANI A, SHIRANI-RAD A, PAZOKI A, NOURMOHAMMADI G, ZARGHAMI R, MOKHTASSI-BIDGOLI A. The impact of source or sink limitations on yield formation of winter wheat (L.) due to post-anthesis water and nitrogen deficiencies. Plant, Soil and Environment, 2010, 56(5): 218-227.
[28] ABDELRAHMAN M, BURRITT D J, GUPTA A, TSUJIMOTO H, TRAN L S P. Heat stress effects on source-sink relationships and metabolome dynamics in wheat. Journal of Experimental Botany, 2019, 71(2): 543-554.
[29] 薛香, 吴玉娥, 陈荣江, 韩占江, 郜庆炉. 小麦籽粒灌浆过程的不同数学模型模拟比较. 麦类作物学报, 2006, 26(6): 169-171.
XUE X, WU Y E, CHEN R J, HAN Z J, GAO Q L.Comparison of different mathematical equations for simulating the qrain filling process of wheat. Journal of Triticeae Crops, 2006, 26(6): 169-171. (in Chinese)
[30] 冯伟, 李晓, 郭天财, 朱云集. 氮用量对不同穗型小麦品种籽粒灌浆及穗部性状的影响. 干旱地区农业研究, 2005, 23(4): 22-25.
FENG W, LI X, GUO T C, ZHU Y J. Effects of nitrogen application levels on grain filling and spike traits of winter wheat cultivars with different spike-type. Agricultural Research in Arid Areas, 2005, 23(4): 22-25. (in Chinese)
[31] DOU X X, SHEN D, ZHANG X H, CHENG J Q, LI S G, SONG J P, WANG H P, ZHANG W, LI X X. Diversity of sex types and seasonal sexual plasticity in a cucumber germplasm collection. Horticultural Plant Journal, 2015, 1(2): 61-69.
[32] CHEN C, CHEN H, ZHANG Y, THOMAS H R, FRANK M H, HE Y, XIA R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Molecular Plant, 2020, 13(8): 1194-1202.
[33] 姚国才, 周朝飞, 钱存鸣, 盛培英, 陈志德. 小麦品种源库性状演变分析及其在育种中的利用. 江苏农业科学, 1994, 22(2): 1-4.
YAO G C, ZHOU C F, QIAN C M, SHENG P Y, CHEN Z D. Evolution analysis of source-sink traits of wheat varieties and their utilization in breeding. Jiangsu Agricultural Sciences, 1994, 22(2): 1-4. (in Chinese)
[34] 张贵合, 郭华春. 马铃薯不同品种(系)的光合特性比较与聚类分析. 作物学报, 2017, 43(7): 1067-1076.
ZHANG G H, GUO H C. Comparison of photosynthetic characteristics and cluster analysis in potato varieties (lines). Acta Agronomica Sinica, 2017, 43(7): 1067-1076. (in Chinese)
[35] 李纯佳, 覃伟, 徐超华, 刘洪博, 毛钧, 陆鑫. 国外引进甘蔗栽培品种的光合气体交换参数遗传差异与聚类分析. 中国农业科学, 2018, 51(12): 2288-2299.
LI C J, QIN W, XU C H, LIU H B, MAO J, LU X. Genetic variations and cluster analysis of photosynthetic gas exchange parameters in exotic sugarcane cultivars. Scientia Agricultura Sinica, 2018, 51(12): 2288-2299. (in Chinese)
[36] 朱峰, 王长发, 苗芳, 张嵩午. 冬小麦品种(系)叶绿素荧光参数的因子分析. 麦类作物学报, 2010, 30(2): 290-295.
ZHU F, WANG C F, MIAO F, ZHANG S W. Factor analysis of chlorophyll fluorescence parameters of winter wheat varieties (lines). Journal of Triticeae Crops, 2010, 30(2): 290-295. (in Chinese)
[37] 池宏康. 冬小麦单位面积产量的光谱数据估产模型研究. 遥感信息, 1995, 10(3): 15-18.
CHI H K. Research on yield estimation model of winter wheat based on spectral data. Remote sensing information, 1995, 10(3): 15-18. (in Chinese)
[38] 夏天, 吴文斌, 周清波, 周勇, 于雷. 基于高光谱的冬小麦叶面积指数估算方法. 中国农业科学, 2012, 45(10): 2085-2092.
XIA T, WU W B, ZHOU Q B, ZHOU Y, YU L. An estimation method of winter wheat leaf area index based on hyperspectral data. Scientia Agricultura Sinica, 2012, 45(10): 2085-2092. (in Chinese)
[39] 郭文善, 封超年, 严六零, 彭永欣, 朱新开, 宗爱国. 小麦开花后源库关系分析. 作物学报, 1995, 21(3): 334-340.
GUO W S, FENG C N, YAN L L, PENG Y X, ZHU X K, ZONG A G. Analysis on source-sink relationship after anthesis in wheat. Acta Agronomica Sinica, 1995, 21(3): 334-340. (in Chinese)
[40] R.A.FISCHER, 黄沃译. 小麦籽粒灌浆时的源: 库互作. 麦类作物学报, 1985, 5(6): 36-39.
FISCHER R A, HUANG W Y. Source-sink interaction during grain filling in wheat. Journal of Triticeae Crops, 1985, 5(6): 36-39. (in Chinese)
[41] 刘晓冰, 张秋英. 试论作物的源库系统. 农业系统科学与综合研究, 1992, 8(2): 131-134.
LIU X B, ZHANG Q Y. On the source-sink system of crops. Systems Science and Comprehensive Studies in Agricultural, 1992, 8(2): 131-134. (in Chinese)
[42] 何闻静, 韩霜, 曹亚娟, 沙莎, 易镇邪, 陈平平. 作物产量形成理论与玉米高产栽培研究进展. 湖南生态科学学报, 2018, 5(2): 36-45.
HE W J, HAN S, CAO Y J, SHA S, YI Z X, CHEN P P. Research progress of crop yield formation theory and high yield cultivation of maize. Hunan Journal of Ecological Sciences, 2018, 5(2): 36-45. (in Chinese)
[43] MURCHIE E H, LAWSON T. Chlorophyll fluorescence analysis: a guide to good practice and understanding some new applications. Journal of Experimental Botany, 2013, 64(13): 3983-3998.
[44] 曹显祖, 朱庆森. 水稻品种的库源特征及其类型划分的研究. 作物学报, 1987, 13(4): 265-272.
CAO X Z, ZHU Q S. Study on characteristics of the relationship between source and sink in rice varieties and their classification. Acta Agronomica Sinica, 1987, 13(4): 265-272. (in Chinese)
[45] 高芳, 刘兆新, 赵继浩, 汪颖, 潘小怡, 赖华江, 李向东, 杨东清. 北方主栽花生品种的源库特征及其分类. 作物学报, 2021, 47(9): 1712-1723.
GAO F, LIU Z X, ZHAO J H, WANG Y, PAN X Y, LAI H J, LI X D, YANG D Q. Source-sink characteristics and classification of main peanut varieties in northern China. Acta Agronomica Sinica, 2021, 47(9): 1712-1723. (in Chinese)
[46] 严威凯. 小麦茎秆运输能力对籽粒产量限制作用的研究. 陕西农业科学, 1988, 34(3): 3-6.
YAN W K. Study on the effect of wheat stem transport capacity on grain yield limitation. Shaanxi Journal of Agricultural Sciences, 1988, 34(3): 3-6. (in Chinese)
[47] 张文虎. 新株型稻与粳/灿杂交稻源库特征与物质运转的研究[D]. 扬州: 扬州大学, 2003.
ZHANG W H. Study on the source-sink characteristics and material transport of new plant type rice and japonica/indica hybrid rice[D]. Yangzhou: Yangzhou University, 2003. (in Chinese)
[48] 远彤, 郭天财, 罗毅, 孙金花. 冬小麦不同粒型品种茎叶组织结构与籽粒形成关系的研究. 作物学报, 1998, 24(6): 876-883.
YUAN T, GUO T C, LUO Y, SUN J H. Study on the relationship between stem-leaf tissue structure and grain formation of winter wheat varieties with different grain types. Acta Agronomica Sinica, 1998, 24(6): 876-883. (in Chinese)
[49] 范平, 张娟, 李新平, 张岱, 赵东旺. 不同小麦品种(系)茎秆组织结构与产量潜力关系研究. 河南农业大学学报, 2000, 34(3): 216-219.
FAN P, ZHANG J, LI X P, ZHANG D, ZHAO D W. Relationship between the anatomical structure of stem and the yield potential of different wheat varieties. Journal of Henan Agricultural University, 2000, 34(3): 216-219. (in Chinese)
[50] 王振林, 贺明荣, 傅金民, 田奇卓, 尹燕枰, 曹鸿鸣. 源库调节对灌溉与旱地小麦开花后光合产物生产和分配的影响. 作物学报, 1999, 25(2): 162-168.
WANG Z L, HE M R, FU J M, TIAN Q Z, YIN Y P, CAO H M. Effects of source-sink regulation on photosynthate production and distribution in irrigated and dryland wheat after anthesis. Acta Agronomica Sinica, 1999, 25(2): 162-168. (in Chinese)
[51] 凌启鸿, 杨建昌. 水稻群体“粒叶比”与高产栽培途径的研究. 中国农业科学, 1986, 19(3): 1-8.
LING Q H, YANG J C. Study on ' grain-leaf ratio ' of rice population and high-yielding cultivation approaches. Scientia Agricultura Sinica, 1986, 19(3): 1-8. (in Chinese)
[52] 贺明荣, 王振林, 曹鸿鸣. 源库关系改变对小麦灌浆期植株光合速率及14C同化物运转分配的影响. 西北植物学报, 1998, 18(4): 82-87.
HE M R, WANG Z L, CAO H M. Effects of source-sink relationship on photosynthetic rate and translocation and distribution of ~ (14) C-assimilates during grain filling stage of wheat. Northwest Botanical Journal, 1998,18 (4): 82-87. (in Chinese)
[53] 孙道杰, 王辉, 闵东红, 李学军, 冯毅. 小麦品种产量改良的限制因素分析. 西南农业学报, 2002, 15(3): 13-16.
SUN D J, WANG H, MIN D H, LI X J, FENG Y. An analysis of the restrictive factors of yield improvement of wheat cultivars. Southwest Agricultural Journal, 2002, 15(3): 13-16. (in Chinese)
[54] 杨建昌, 王朋, 刘立军, 王志琴, 朱庆森. 中籼水稻品种产量与株型演进特征研究. 作物学报, 2006, 32(7): 949-955.
YANG J C, WANG P, LIU L J, WANG Z Q, ZHU Q S. Evolution characteristics of grain yield and plant type for mid-season indica rice cultivars. Acta Agronomica Sinica, 2006, 32(7): 949-955. (in Chinese)
[55] 马冬云, 郭天财, 宋晓, 岳艳军, 查菲娜, 韩巧霞. 源库改变对小麦籽粒蛋白质含量及其组分的影响. 西北农业学报, 2009, 18(1): 112-116.
MA D Y, GUO T C, SONG X, YUE Y J, ZHA F N, HAN Q X. Effects of source-sink manipulation on accumulation of grain protein and its component in wheat (L.). Acta Agriculturae Boreali-Occidentalis Sinica, 2009, 18(1): 112-116. (in Chinese)
[56] 刘晓冰, 李文雄. 源库改变对小麦籽粒蛋白质、淀粉含量和产量的影响. 东北农业大学学报, 1996, 27(4): 321-325.
LIU X B, Li W X. Effects of source-sink alteration on grain protein, starch content and yield of wheat. Journal of Northeast Agricultural University, 1996, 27(4): 321-325. (in Chinese)
[57] 张英华, 周顺利, 张凯, 王志敏. 源库调节对小麦不同品种籽粒微量元素及蛋白质含量的影响. 作物学报, 2008, 34(9): 1629-1636.
ZHANG Y H, ZHOU S L, ZHANG K, WANG Z M. Effects of source and sink reductions on micronutrient and protein contents of grain in wheat. Acta Agronomica Sinica, 2008, 34(9): 1629-1636. (in Chinese)
[58] 马兴林, 王庆祥. 通过剪叶改变源库关系对玉米子粒营养组分含量的影响. 玉米科学, 2006, 14(6): 7-12, 22.
MA X L, WANG Q X. Effects of source-sink alteration by leaf-clipped treatment on grain nutrition components accumulation. Journal of Maize Sciences, 2006, 14(6): 7-12, 22. (in Chinese)
Identification of Excellent Wheat Germplasms and Classification of Source-sink Types
NAN Rui1, YANG YuCun1, SHI FangHui1, ZHANG LiNing1, MI TongXi1, ZHANG LiQiang2, LI ChunYan2, SUN FengLi1, XI YaJun1, ZHANG Chao1
1College of Agronomy, Northwest A&F University/Key Laboratory of Wheat Biology and Genetic Breeding, Ministry of Agricultural and Rural Affairs, Yangling 712100, Shaanxi;2Baoji Longxian Seed Workstation, Baoji 712100, Shaanxi
【Objective】The aim of this study is to screen the evaluation indexes of wheat source-sink and classify the source-sink types. In addition, the relationships between different source-sink types and the agronomic traits, yield and grain quality of wheat were also clarified, which provides a better understanding of wheat source-sink metabolism and wheat breeding. 【Method】In this study, the related agronomic traits of source metabolism and sink metabolism of 190 wheat varieties which from different regions were measured. Then, the source-sink metabolic capacity of wheat was evaluated by principal component analysis, and the superior wheat materials were selected according to the composite score. Furthermore, the hierarchical clustering was conducted based on the source size (leaf area), source activity, sink number and sink activity. Then, based on the clustering results, the source-sink types of wheat were classified and the source-sink characteristics of different region of wheats were analyzed. Meanwhile, the agronomic, yield and quality traits of different wheat source-sink types were compared and analyzed. 【Result】For better understanding the results, six indicators related to source activity were converted into three independent indicators (photochemical quenching coefficient, maximum photosynthetic potential, chlorophyll content), and five indicators related to sink activity were converted into two independent indicators (maximum filling rate, filling duration) based on the principal component analysis. The cumulative contributions of three source activity indicators and two sink activity indicators were 82.80% and 92.90%, respectively. Then the top 10 wheat varieties were screened based on the source activity, source size (leaf area), sink activity, and sink number (number of spike grains). According to the source-library relationship, all the wheat varieties were divided into three major categories and eight subcategories, including sufficient source-weak sink type (medium source-weak sink type, strong source-medium sink type), weak source-sufficient sink type (medium source-strong sink type, weak source-medium sink type) and source-sink balance type (weak source-weak sink type limited by sink activity, weak source-weak sink type limited by source activity and grain number per spike, medium source-medium sink type, and strong source-strong sink type). 76.84% of wheat lines were contained in three subcategories (weak source-weak sink type, the medium source-strong sink type, and strong source-strong sink type), other wheat lines were distributed in other subcategories evenly. Most wheat varieties of China showed similar source-sink relationship, which presented the medium level of source activity, leaf area and grain number per spike, while the sink activity was high. But the sink activity of wheat varieties which were cultivated at the middle and lower reaches of the Yangtze River is low. The plant height, length of uppermost internode and spikelet number in different categories indicated that the sufficient source-weak sink type>source-sink balance type>weak source-sufficient sink type, and the dry protein content, dry wet gluten content and sedimentation value in different categories showed that source-weak sink type > source-sink balance type > weak source-sufficient sink type. Grain water absorption showed that the stronger the sink activity had the higher the water absorption rate. The yield of per plant was different among different source-bank groups, but it was positively correlated with the source activity and the number of grains per spike in the three subcategories which contained most wheat varieties. 【Conclusion】 In this study, photochemical quenching coefficient, maximum photosynthetic potential and chlorophyll content could be used as the main indexes to evaluate the activity of wheat source. The maximum filling rate and filling duration could be used as the main indexes to evaluate the activity of wheat sink. In practical production, the wheat yield could be improved by increasing the number of grains per spike and the activity of source. When the supply capacity of source is stronger than the absorption capacity of sink, the plant height, peduncle length, spikelet number, dry protein content, dry wet gluten content and sedimentation value would be increased. Strong sink activity would help us to improve the water absorption of wheat grains.
wheat; evaluation indicators; source-sink type; agronomic traits; yield traits; quality traits

10.3864/j.issn.0578-1752.2023.06.001
2022-11-17;
2023-01-03
陕西省重点研发计划(2022NY-193)、西安市科技计划(22NYYF001)
南瑞,Tel:15029299873;E-mail:2416517720@qq.com。通信作者张超,Tel:18292883757;E-mail:ahzc2009@163.com
(责任编辑 李莉)
