γ-氨基丁酸对大豆种子萌发及根系形态建成的调控效应
2023-03-29强斌斌金喜军周伟鑫褚霈宇张玉先
强斌斌,金喜军,周伟鑫,刘 佳,褚霈宇,张玉先
(黑龙江八一农垦大学农学院,黑龙江 大庆 163319)
大豆富含优质植物蛋白和油分,是我国主要经济作物之一,在国民经济和人民生活中占有举足轻重的地位;其产量表现主要取决于品种特性和环境条件。优良品种和适宜环境条件很大程度上保证了作物产量水平,而采取更加有效的手段进一步挖掘产量潜力是栽培科研工作者追求的目标。壮苗是保证作物高产和稳产的基础,根粗、根表面积、根体积等形态指标是豆科作物壮苗的关键指标[1-2],因此根系的生长状况作为壮苗的重要因素之一,对作物产量提高具有重要的意义[3-4]。众所周知,作物已经在形态、生理、生化和分子水平上进化出各种调控机制来适应多变的环境,以便于物种延续、发展和繁荣。在错综复杂的调控机制中,激素调节是非常重要的方面。植物激素参与植物生长发育的每一个阶段,包括萌发、营养生长、生殖生长、成熟,控制着各个器官的形态、大小和数量,并对外界环境如温度、水分、重金属、虫害、病害等作出反应,起到协调植株生长发育与环境条件之间关系的作用[5]。除了我们熟知的生长素、赤霉素、细胞分裂素、脱落酸和乙烯以外,近年来还有一些调节物质被认定为新型植物激素,其中就包括γ-氨基丁酸(GABA)。GABA是一种非蛋白质氨基酸,存在于包括一些植物和动物在内的许多生物中[6]。科研人员首先在马铃薯块茎中发现了天然存在的GABA,而后证实其存在于其他多种植物中[7]。研究表明,植物内源GABA可以作为植物调控生长和发育的内源性信号分子发挥代谢作用[8];此外,GABA也可参与环境适应行为,如pH的变化、冷和热休克反应、抗虫机制和氮代谢[7],说明植物内源GABA含量的变化与其生长发育状态存在密切关系。Li等[9]研究表明,外源GABA的施用可以显著提高玉米根系和地上部幼苗的鲜重;李敬蕊等[10]在研究中发现GABA浸种可显著促进小白菜生长,提高其生物量,并且影响了作物的硝酸盐代谢。以上研究说明外源GABA可以影响不同类型的植物和植物的不同生长阶段,具有与内源GABA相同的效应。然而,关于外源GABA调控大豆种子萌发及其根系形态建成的研究鲜有报道。
本研究以龙垦310、合丰50为试验材料,设置不同浓度GABA进行浸种处理,通过萌发和根系形态关键指标的比较,明确GABA对大豆种子萌发和根系形态建成的影响;在最佳浓度下比较分析萌发和根系发育关键生理指标,明确GABA对大豆萌发和根系发育生理调控效应;最后,结合形态、生理指标结果明确氨基丁酸对大豆种子萌发、根系形态建成的调控效应,以期为实际生产中通过施用外源GABA培育壮苗、提高大豆产量提供理论依据和指导。
1 材料与方法
1.1 供试材料
选用黑龙江主栽大豆品种龙垦310和合丰50为试验材料,龙垦310为高蛋白品种,无限结荚习性;合丰50为高油大豆品种,亚有限结荚习性。γ-氨基丁酸(GABA)购自sigma公司,分析纯。
1.2 试验设计
试验于2021年在黑龙江八一农垦大学生物技术中心试验室内进行。首先挑选大小均匀、无破损和病斑的种子,10%次氯酸钠消毒15 min后用蒸馏水冲洗5次,在黑暗条件下分别浸泡在浓度为0、5、10、25、50、100 mmol·L-1氨基丁酸溶液中8 h,使其达到吸胀状态。之后在直径为9 cm培养皿中,垫两层无菌滤纸并加入5 ml蒸馏水,每个培养皿放10粒种子,3次重复,置于25℃恒温培养箱中发芽,培养7 d。每2 d更换一次滤纸,采用称重法每天补充相应的溶液使滤纸达到湿润状态,观察记录种子萌发相关的各项指标,于发芽开始后的1、3、4、5、7 d取样测定萌发相关生理指标,在第7天观察并测定根系形态指标,筛选出最佳浓度。以最佳浓度为处理浓度,蒸馏水为对照,再次进行发芽和根系观察试验,经对比分析进一步明确处理效果。
1.3 测定指标与方法
1.3.1 萌发指标 处理1 d后开始每天观察并记录种子发芽数(以胚根突破种皮达到种子1/2定义为发芽),并测算萌发指标。
发芽势=培养n天后发芽数/供试种子数×100%
发芽指数GI=∑(Gt/Dt)
式中,Gt指在t日内的发芽数,Dt为相应的发芽天数(d)。
活力指数VI=GI×S
式中,S为幼苗鲜重(g)。
1.3.2 形态指标 种子萌发第7天使用FGX-A型根系分析系统扫描根系,并用形态学分析软件winRHIZO分析根长、根面积、根体积、侧根数;使用游标卡尺测定根粗,测量点选择下胚轴与主根的根节处;之后用滤纸吸干根系水分,用万分之一天平称量其鲜重。
1.3.3 生理指标 种子萌发第7天取根系及子叶用液氮冷冻后于-80℃保存。超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、抗坏血酸过氧化物酶(APX)和过氧化氢酶(CAT)活性的测定,根系活力、游离氨基酸含量的测定参照李小方等[11]方法进行。蒽酮法测定可溶性糖含量[12],考马斯亮蓝G-250染色法测定可溶性蛋白质含量[11]。电导率采用电导率仪进行测定。GABA含量基于韩延丽等[13]改良后的显色反应测定。
1.4 数据处理与分析
数据采用SPSS 20.0进行显著行分析,使用Prism 8.0进行作图。
2 结果与分析
2.1 GABA浸种对大豆种子萌发的影响
由表1可知,不同浓度GABA浸种对合丰50和龙垦310大豆种子发芽率、发芽指数、活力指数均有不同程度的影响。与对照相比,合丰50在5、10、25 mmol·L-1GABA浸种处理下,第1天和第2天的发芽率分别提高了58.8%和25.6%、105.8%和55.7%、88.4%和55.1%;第7天后,各浓度处理间发芽率无显著差异;发芽指数和活力指数分别提高了16.5%、27.1%、22.5%和28.2%、63.3%、30.7%。100 mmol·L-1GABA浸种处理的发芽指数、活力指数均较对照有所下降。说明适宜浓度的GABA浸种可以显著提高发芽指数和活力指数,有利于种子提前发芽,而高浓度100 mmol·L-1则会抑制种子萌发。对于龙垦310而言,10 mmol·L-1和25 mmol·L-1外源GABA处理有利于促进其种子的萌发,其中第1天、第2天、第3天发芽率及发芽指数、活力指数较对照组分别提高了39.3%和51.0%、22.6%和33.0%、19.5%和14.9%、12.2%和14.7%、98.9%和77.9%。综上可知,随GABA浓度的升高,两品种的发芽率、发芽指数、活力指数呈先上升后下降的趋势,10~25 mmol·L-1GABA浸种处理对种子萌发的促进效果最显著,而浓度过高则起抑制作用,不利于种子的萌发。
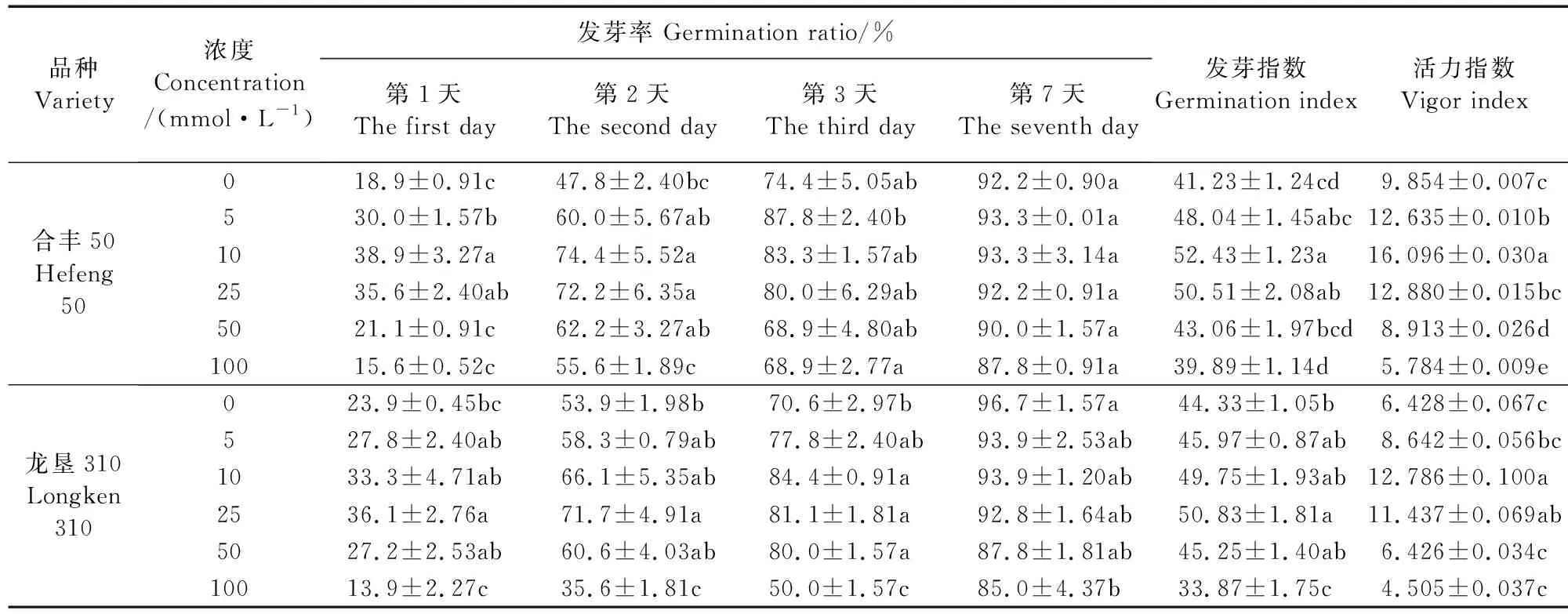
表1 GABA浸种对大豆种子萌发指标的影响Table 1 Effects of soaking seeds with GABA on soybean seed germination index
2.2 GABA浸种对大豆根系形态指标的影响
如图1和图2所示,不同浓度GABA浸种对合丰50和龙垦310大豆根系各项形态指标的影响有所差异。10 mmol·L-1GABA浸种处理对两品种的根长、侧根数、根鲜重促进显著,较对照组分别提高了27.3%和27.1%、93.4%和90.5%、28.5%和77.2%,其中侧根数增加最明显。GABA对两品种的根表面积和根体积调控存在差异,合丰50在25 mmol·L-1GABA浸种处理下根表面积和根体积分别较对照处理提高了32.7%和22.7%;与对照相比,龙垦310在5 mmol·L-1GABA浸种处理下根表面积增加效果最显著,增幅为35.8%,浓度为25 mmol·L-1对根体积提升效果最显著,增幅为20.9%。高浓度100 mmol·L-1时合丰50根长、根表面积、根体积较对照分别降低了58.5%、42.9%、34.6%,龙垦310根长、根表面积较对照分别降低了59.4%、37.0%。不同的是,合丰50和龙垦310根粗随GABA浸种浓度的增加呈逐渐增加后平稳的趋势,100 mmol·L-1浓度处理分别较对照处理增加了37.9%和23.6%。以上结果表明,适宜浓度的GABA浸种处理对大豆根长和侧根数可起到显著促进作用,高浓度GABA则起抑制作用。
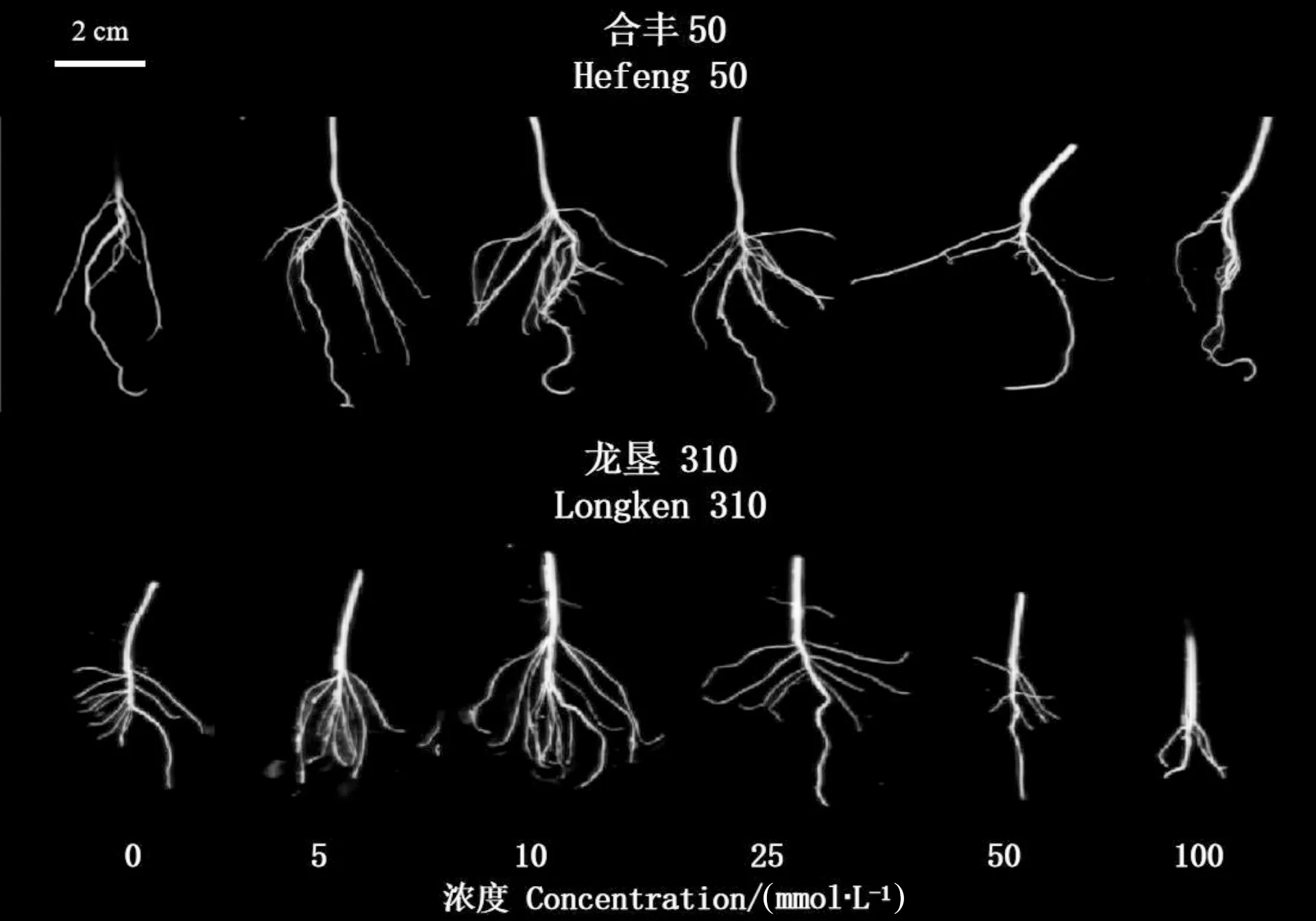
注: 图中的2 cm代表根系标尺。Note: The 2 cm in the figure represents the root scale.图1 GABA浸种对大豆根系表型的影响Fig.1 Effects of soaking seeds with GABA on root phenotype of soybean
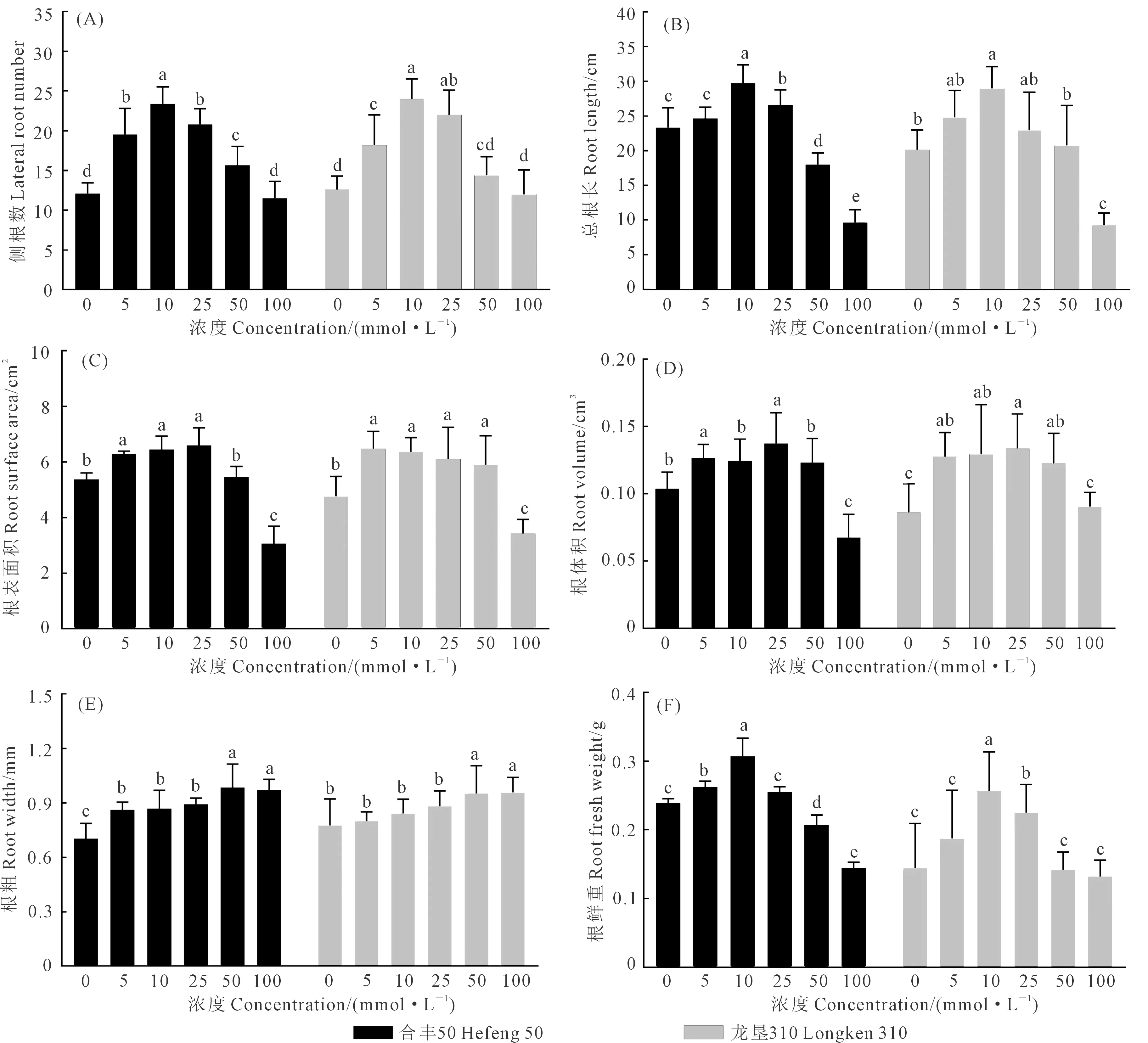
注: 不同小写字母代表处理间差异显著(P<0.05) ,下同。Note: Different lowercase letters represent significant differences among treatments (P<0.05),the same below.图2 GABA浸种对大豆根系形态指标的影响Fig.2 Effects of soaking seeds with GABA on root morphological indexes of soybean
2.3 GABA浸种对大豆根系活力、电导率的影响
如图3所示为不同浓度GABA浸种对大豆根系活力和电导率的影响。合丰50和龙垦310在10 mmol·L-1浓度时的根系活力显著高于对照组,增幅分别为26.8%和21.3%,而浓度为50 mmol·L-1和100 mmol·L-1时对根系活力产生抑制现象。2个品种电导率在5、10、25 mmol·L-1浓度时显著低于对照处理,而在100 mmol·L-1浓度下显著高于其他处理。说明10~25 mmol·L-1外源GABA浸种可以显著增加大豆根系活力,降低根系电导率,浓度过高,可能导致膜透性增大,表现出抑制效果。
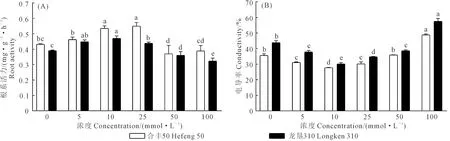
图3 GABA浸种对大豆根系活力与电导率的影响Fig.3 Effects of soaking seeds with GABA on root activity and electrical conductivity of soybean
2.4 GABA浸种对大豆根系抗氧化酶活性的影响
如图4所示为GABA浸种对大豆根系抗氧化酶活性的影响,可以看出,外源GABA浸种可改变大豆根系抗氧化酶活性。随GABA浓度增加,合丰50和龙垦310抗氧化酶SOD、POD、CAT、APX活性均呈先上升后下降趋势,浓度为10 mmol·L-1时增加效果最显著,分别较对照提高了91.3%和88.3%、38.5%和33.3%、48.7%和35.8%、64.3%和46.2%。GABA浓度大于25 mmol·L-1时,抗氧化酶活性出现缓慢下降趋势。以上结果表明,10 mmol·L-1浓度GABA浸种处理能够显著提升大豆萌发期间根系抗氧化酶活性,促进根系生长发育,而高浓度100 mmol·L-1GABA浸种处理导致抗氧化酶活性变化不显著或者降低,不利于根系生长发育。
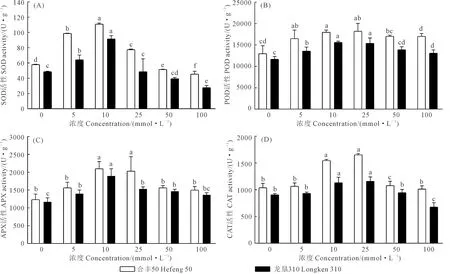
图4 GABA浸种对大豆根系抗氧化酶活性的影响Fig.4 Effects of soaking seeds with GABA on antioxidant enzyme activities in soybean roots
2.5 GABA浸种对大豆种子萌发期代谢物质的影响
如图5所示为不同浓度GABA浸种对大豆萌发期间子叶与根系代谢物质及渗透调节物质的影响。可溶性糖与可溶性蛋白是植物新陈代谢的重要产物,图5(A、B)为GABA对大豆根系及子叶中可溶性糖与可溶性蛋白含量的影响。不同浓度GABA对两品种可溶性糖与可溶性蛋白影响趋势一致。随GABA浓度的增加,合丰50子叶和根系中的可溶性糖与可溶性蛋白均呈先上升后下降的趋势,且均在10 mmol·L-1浓度时含量最高,与对照相比,子叶增幅为54.23%和22.33%,根系增幅为17.46%和26.14%。
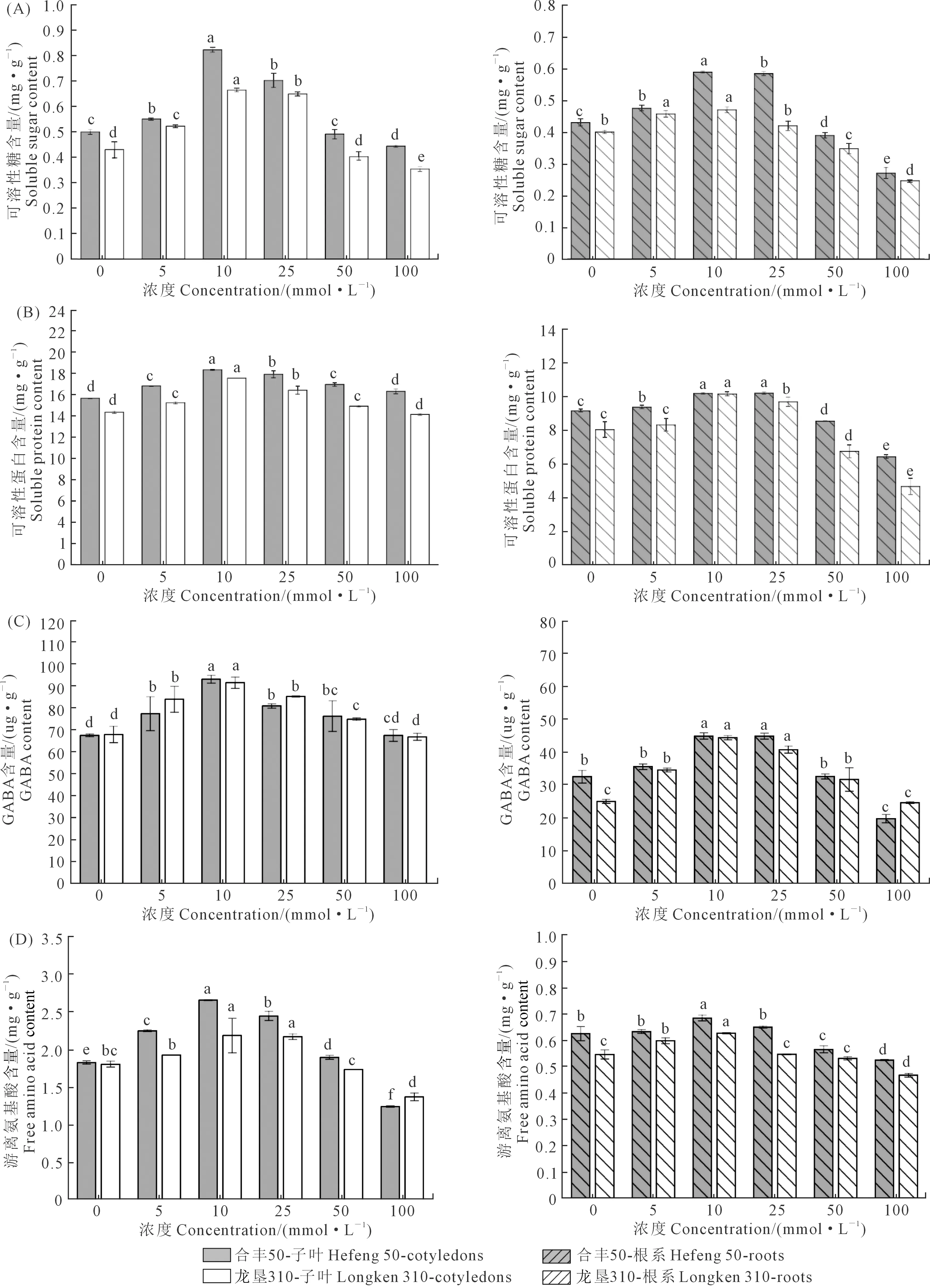
图5 GABA浸种对大豆子叶和根系谢物质含量的影响Fig.5 Effects of soaking seeds with GABA on metabolites content in cotyledons and roots of soybean
100 mmol·L-1处理时,子叶可溶性蛋白含量与对照无显著差异,而根系可溶性糖与可溶性蛋白含量显著低于对照。如图5(C、D)所示,外源GABA浸种显著改变子叶与根系内源GABA的含量,合丰50子叶与根系内源GABA含量和游离氨基酸含量均随GABA浓度的增加呈先上升后降低的趋势。其中,10 mmol·L-1浓度处理的内源GABA含量与游离氨基酸含量达到最大值,与对照组相比,子叶中的GABA含量和氨基酸含量分别增加了37.7%和45.3%,根系分别增加了44.3%和9.52%。龙垦310各指标表现趋势与合丰50基本相同,但相同浓度下合丰50子叶与根系中的可溶性糖、可溶性蛋白、游离氨基酸含量均高于龙垦310。以上结果表明,在大豆种子萌发期间,10 mmol·L-1GABA浸种对子叶与根系代谢、种子萌发和根系生长的促进效果显著,在合丰50上表现更明显。
3 讨 论
γ-氨基丁酸(GABA)作为一种四碳非蛋白质氨基酸,在植物中担任代谢物质和信号物质的双重角色[6]。外源GABA通过调控植物信号转导、转录调控、激素合成、活性氧生成和多胺代谢等相关基因的表达,可提高植物抗逆能力[14]。无论在应激条件还是非应激条件下,外源GABA被植物吸收后,其作用方式与内源GABA相同[15],对不同植物类型和植物不同生长阶段均可起调控作用[16]。外源GABA可调控脱落酸和乙烯生物合成,脱落酸具有促进种子休眠的作用,乙烯具有促进种子萌发的作用。近年来,大量研究表明,GABA浸种可提高种子发芽率、发芽速度,促进胚轴和胚根的生长,且不同物种间的浓度响应有所差异[17-18]。本研究发现随GABA浸种浓度增加,大豆种子在萌发期间内源GABA含量和游离氨基酸含量呈先上升后下降趋势,10 mmol·L-1浓度处理下内源GABA含量较对照处理增加最显著,同时该浓度下大豆的发芽势、发芽指数和活力指数也显著提高。这可能是因为适宜浓度的GABA可以降低ABA的含量,提高乙烯含量,进而调控内源激素促进种子的萌发。这与施征等[19]研究得出的10 mmol·L-1外源GABA能提高种子萌发率,而高浓度GABA具有抑制作用的研究结果一致。
种子萌发过程中各种代谢活动会产生活性氧(ROS),对细胞膜、核酸、蛋白质等大分子物质产生破坏作用[20]。活性氧的产生往往会造成细胞膜的损伤及各种细胞结构和细胞器的解体,最终导致细胞死亡[21]。GABA在植物中具有清除活性氧的作用[8],已有研究证明GABA可以显著提高水稻、黑麦草等作物中多种抗氧化酶活性[22-23]。本研究发现适宜浓度(10 mmol·L-1)的GABA浸种处理可显著提高根系SOD、POD、CAT、APX活性,可能是因为GABA诱导的抗氧化活性的变化,有助于增强抗氧化酶的活性,提高细胞清除ROS的能力,从而维持了细胞完整性[24]。本试验结果与 Li等[9]的结果一致,即适宜浓度的GABA提高了作物中各种抗氧化酶的活性,清除ROS的产生。也有研究显示,在MS培养基上用外源GABA抑制了拟南芥幼苗根系的伸长,而用1/8的培养基培养可以促进根系的伸长,这可能与硝酸盐的水平不同有关[25]。
淀粉的分解代谢过程为种子的萌发和根系的生长提供有效的碳水化合物,可溶性糖对维持细胞的充盈度和能量供应起了重要的作用[26];而可溶性蛋白作为重要的渗透调节物质和营养物质,对参与细胞建成的生命物质及生物膜具有保护作用[27]。有研究表明,GABA可诱导淀粉分解,为白三叶种子萌发和生长提供有效的碳水化合物[26]。本研究结果表明在大豆萌发期间,无论是子叶还是根系,可溶性糖和可溶性蛋白含量均随外源GABA浓度的增加呈先上升后下降的趋势,其含量在10 mmol·L-1浓度处理时达到最高。说明适宜浓度GABA浸种可促进淀粉代谢,提高大豆子叶和根系可溶性糖和可溶性蛋白的含量,为种子萌发提供更多碳水化合物。这可能是因为GABA与激素相互作用,调控了种子内部蛋白和糖类代谢,从而促进种子萌发[28]。
根系活力是客观反映根系生命活动重要的生理指标,体现了根系吸收水分和营养物质的能力[29];根系的形态特征体现了其对水分、矿质养分的吸收效率。侧根的数量和位置决定了根系的大小和结构,根系结构特征对作物产量有很大的影响[4]。有研究发现GABA可以调控脱落酸和乙烯并诱导根系所需生长素的生物合成[30],且证明IAA/ABA诱导铝活化的苹果酸转运蛋白(ALMT)家族基因在根尖表皮细胞的质膜上表达[31]。此外,GABA对ALMT活性的调节导致了根系生长的变化,改变根系pH。本研究结果表明,外源10 mmol·L-1GABA提高了根系活力,促进总根长、侧根数及根表面积增加,而高浓度明显抑制了主根的生长与侧根的形成。可能是因为GABA与乙烯、脱落酸和生长素等激素产生互作效应,共同调控种子的萌发与根系的生长;也可能是因为GABA作为一种可被根系吸收的氨基酸态氮源,参与硝酸盐吸收,适量浓度的GABA处理可促进植物体对硝酸盐的吸收,刺激根系伸长[32]。综合来看,适宜外源GABA浸种可能通过调控内源GABA与其他激素间的互作效应或参与硝酸盐的吸收,促进种子的萌发和根系的生长与发育,而浓度过高时会抑制这种效果,这与Renault等[33]在拟南芥中研究发现的高浓度对下胚轴和初生根有抑制作用的研究结果一致。
4 结 论
适宜浓度的GABA浸种可提高大豆种子发芽指数和活力指数,增加根长、侧根数、根表面积,提高种子萌发过程中抗氧化酶活性、根系活力和代谢活性,有利于种子萌发和根系形态建成。综合分析发现,对于合丰50和龙垦310,10 mmol·L-1的GABA浸种浓度对其种子萌发及根系形态建成的促进效果最佳。
