不同生长阶段苘麻生物量分配对种群密度和土壤水分的响应
2023-03-23令狐克念
令狐克念 王 姝
(贵州大学林学院,森林生态研究中心,贵阳 550025)
生物量分配是生活史理论的核心概念[1],生物量分配的差异往往比细胞或分子水平的生理机制更重要[2-3]。最优分配理论认为,植物会优先把生物量分配到获取最有限资源的器官中,以使获取受限资源的能力最大化[4-5]。例如,当地下资源如水分或养分欠缺时,植物会将更多生物量分配给根部,降低茎和叶生物量的分配;地上资源如光照受限时,植物往往增大茎和叶生物量分配[6-7]。与非生物环境因素的影响相比,生物环境因素,如竞争[8]、采食作用[9]等,对植物生物量分配的影响复杂得多。
种群密度是自然界中对植物存活、生长和繁殖有重要影响的生物环境因素之一。密度增大不一定导致种内竞争,因为种内相互作用还包括促进作用[10]。大多数类似研究仅从种内或种间竞争对植物影响的角度探讨植物的可塑响应,忽略了密度影响的复合性和动态性。首先,密度增大可能导致多种地上和地下资源同时受限[11],也会使植物间相互作用产生竞争或促进效应,诱导植物同一个特征上的多种可塑响应。植物生物量分配对密度增大的响应是否仍符合最优分配理论,尚没有清晰一致的结论。这可能因为,植物生物量分配对密度的可塑响应不仅随土壤条件而变化,还受个体生长阶段的影响[12-13]。土壤水分[14]、养分[13,15]等非生物环境条件影响竞争或促进作用的相对强度,进而改变植物生物量分配对密度的响应。例如,当土壤养分条件较好时,植株个体更大,地上竞争更强烈,茎部额外伸长或茎生物量分配的增加也会更显著[16],减少根生物量的分配[11-12];土壤养分条件欠缺则加剧高密度种群的地下竞争,更容易导致根、叶、繁殖器官生物量分配的降低[12,17]。与湿润条件相比,干旱条件下种内相互作用更容易促进植物地上生物量的积累,降低地下生物量和根冠比[18]。另外,不仅高密度种群植物间相互作用强度会随时间而变化,植物个体生物量分配方式也随生长阶段而变化(异速生长)[11-12,19],如在生长的早期阶段,相对于低密度种群,高密度种群植物更容易因种内相互关系中的促进作用而使根冠比增大[14];而在生长后期,植物个体的增长导致竞争强度增大,抵消了促进作用,进而导致根冠比降低或不变[20]。因此,生物量分配对密度的可塑响应也会因生长阶段而异。
水分是影响植物存活、生长与繁殖的关键性非生物环境因子。研究表明,干旱会加剧根冠比对种内相互作用的消极响应(降低)和地上生物量对种内相互作用的积极响应[21],意味着干旱加剧地下竞争,减弱地上竞争。但相关研究往往仅观测了根冠比的响应,关于地上不同器官生物量分配对密度的可塑性研究较少,更缺少对生物量分配响应动态的关注(但见[12-13])。另一方面,密度也显著影响植物对水分条件的可塑性[3]。与单独生长的植物个体相比,存在同种植物相互作用时,根冠比更容易在干旱条件下降低[13]。相关研究更多地关注非生物环境条件如何影响植物对密度的可塑响应[22-23],而关于密度是否及如何影响植物对水分的可塑响应很少有研究(但见[21]),更缺少对生物量响应动态的关注。
苘麻(Abutilon theophrasti)是研究区域内常见的草本植物,也经常作为研究可塑性的模式植物。本研究以苘麻为研究对象,在土壤水分增加和对照2 种条件下,对植物进行3 种密度处理,测量并计算根、茎、叶等器官和繁殖部分生物量及其分配,主要探讨以下问题:(1)各器官生物量分配是否会对密度产生可塑响应?(2)土壤水分增加是否影响各器官生物量分配对密度的响应?这种影响是否随生长阶段而变化?(3)种群密度增大是否影响各器官生物量分配对水分的响应?这种影响是否随生长阶段而变化?
1 材料与方法
1.1 研究对象
苘麻系锦葵科(Malvaceae),苘麻属(Abutilon),1 年生草本植物,是中国和印度常见的本地种,目前广泛分布于世界各地。苘麻为直立生长,茎粗壮,高度可达1.0~1.5 m,且生长较快,3个月内达到生殖成熟期,约5个月内完成生命周期[19]。苘麻广泛分布于易受干扰的自然生境,在生物量分配、形态特征和结构方面有显著的可塑性[10]。
1.2 试验设计
研究地点位于吉林省长岭县东北师范大学草地生态研究野外实验站,地理位置为44°40′N,123°44′E。研究区气候类型为中温带大陆性季风气候,年平均气温4.9 ℃,10 ℃以上有效积温2 919 ℃,年降水量470 mm,无霜期约为140 d,植物生长季主要在6—8 月,其平均气温均超20 ℃;6—9 月降水量占70%~80%,全年最高。试验区内的土壤表现出一定程度的盐碱性,一般情况下不影响植物的生长。
苘麻种子在2006年秋季收集于实验站附近的野生种群,保存于-4 ℃干燥环境中。试验开始于2007 年6 月7 日,结束于7 月29 日,在野外实验站附近的田间实验区进行。实验区土壤为沙壤土,生长季土壤pH 8.3 养分含量:有效磷1.1 mg·kg-1,有效氮21.0 mg·kg-1,有机碳3.1 mg·kg-1。为避免土壤营养分布不均,播种前以20 g·m-2的比率施加可溶性颗粒肥料[(NH4)2HPO4,m(N)∶m(P)=18∶46]。采用裂区试验设计,整个试验地分成2 个部分,分别为加水(AW)和对照处理(CK),2 块水分处理的试验地之间间隔1 块试验地,以保证2 块试验地之间水分不会相互渗透。每种土壤水分处理下划分为9个小区(2 m×2 m),3个密度处理和3个区组随机分布,为低(13.4 株·m-2)、中(36 株·m-2)和高(121株·m-2)密度处理,分别对应为30 cm,20 cm和10 cm的植株间距。
密度处理设置主要依据苘麻的生长特点:在试验末期,苘麻的侧枝最长长度可达20 cm,主根(直径>1~3 mm)长度最长达20~30 cm,侧根长度一般不超过20 cm[13],因此低密度设置为植株间距30 cm 可以最大可能地避免植株之间的相互作用,而中密度设置为植株间距20 cm,使植物在生长中后期发生一定程度的相互干扰,高密度设置为植株间距10 cm,可以使植物之间发生强度较大的相互干扰。植物生长过程中,加水(AW)和对照(CK)处理分别用人工灌溉和不灌溉进行控制。加水处理中,在土壤表面水分低于70%时进行人工灌溉,使土壤水分达到饱和状态,保持在70%~90%田间持水量;对照处理不灌溉,土壤水分保持依靠自然降水,土壤水分为30%~50%田间持水量。苘麻种子播种于2007年6月7日,大部分种子在播种后4 d 发芽、出土。幼苗生长到4~5 叶期建苗,调整到目标密度。
1.3 数据收集和分析
在植物生长20 d(6 月27 日)和50 d(7 月29日)时进行取样,2 个时期分别处于苘麻的营养生长期和繁殖生长早期,以比较不同时期苘麻对密度和水分交互作用响应的差异。每次取样时,每种水分处理下,每个密度和区组各选取5~6 株,最大取样总数为:6 株×3 个密度×3 个区组×2 种水分条件×2 个阶段=216 株。取样后,每株植物分为根、茎(连接植物根到顶端的茎秆部分和侧枝茎)、叶片、叶柄等器官和繁殖部分,分别装入牛皮纸信封,置于烘箱中75 ℃烘干72 h 至恒质量,称量质量。总生物量为各器官干质量的总和,繁殖器官生物量为花和果实干质量的总和,并计算各器官生物量比率。
所有数据分析采用SPSS 21.0 软件,作图采用ORIGIN 2018 软件。分别在2 个生长阶段,对总生物量用双因素方差分析(two-way ANOVA),其余特征用双因素协方差分析(two-way ANCOVA),影响因素为种群密度和土壤水分,协方差分析中总生物量为协变量(covariate)。每个生长阶段和土壤水分条件下,密度对总生物量的影响用单因素方差分析(one-way ANOVA),对其他特征的影响用单因素协方差分析(one-way ANCOVA),总生物量为协变量。当总生物量(个体大小)对特征响应与环境因素的变异有显著影响时,此特征被认为表现为表观可塑[10],特征不依赖于总生物量的变异被认为是真正的可塑[3,13]。用一般线性模型(General Linear Model)程序中的最小显著差法(LSD)进行多重比较。
2 结果与分析
2.1 密度与水分交互对生物量分配的可塑影响
由表1 可知,在植物的2 个生长阶段,总生物量(个体大小)对大多数特征均有显著影响;水分处理只影响茎生物量比率(20 d 时,P<0.05),密度对总生物量、根和叶柄生物量比率及根冠比有显著影响(50 d 时,P<0.05),水分与密度交互作用在20 d 时对根生物量比率有显著影响,在50 d 时对总生物量和茎生物量比率有显著影响。
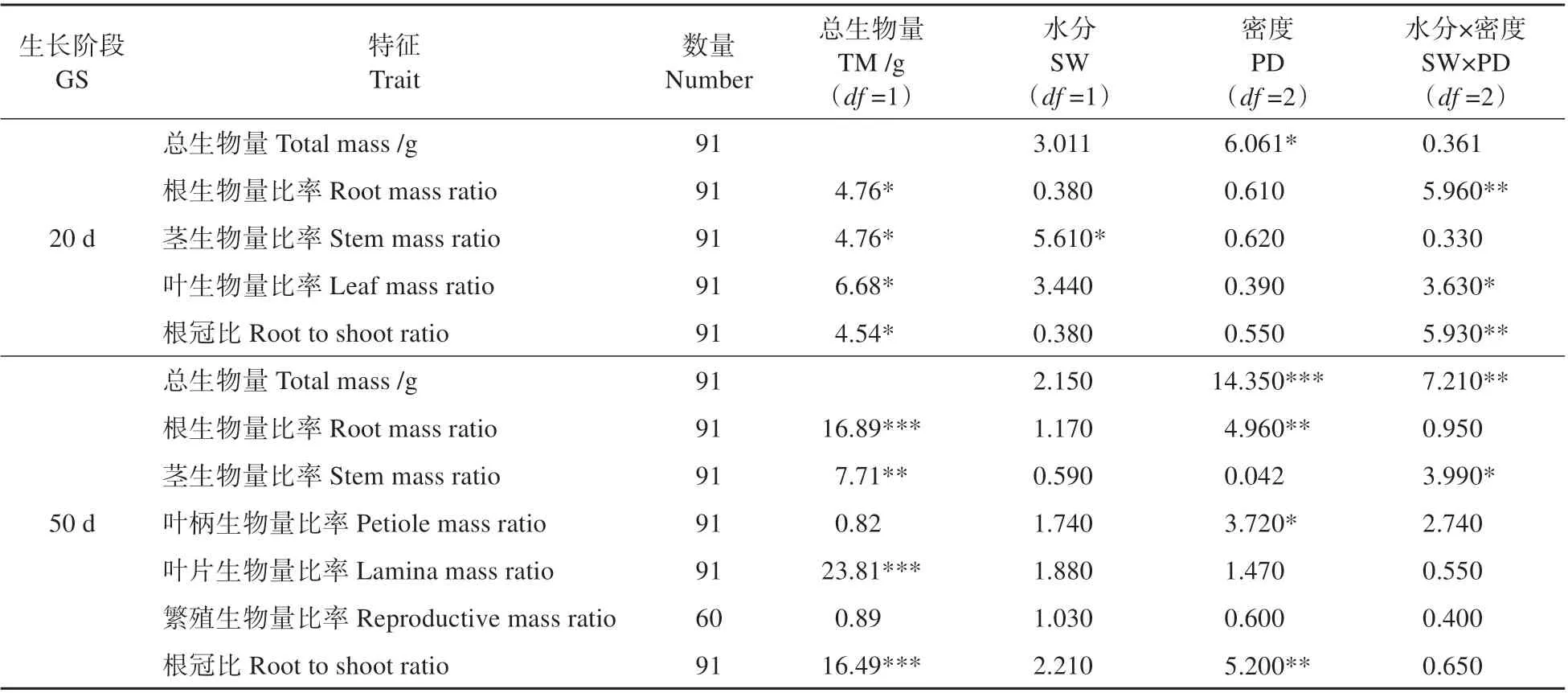
表1 水分、密度及其交互作用对苘麻生物量特征影响的双因素协方差分析Table 1 Two-factor covariance analysis of the effects of water,density and their interaction on mass traits of A.theophrasti
2.2 不同水分处理下生物量分配对密度的可塑响应
由图1 可知,植物生长20 d 时,密度对总生物量影响不显著;50 d时,对照处理(CK)下,低、中密度下总生物量分别为3.85、3.60 g,高密度下总生物量为1.38 g,平均降低了63.5%(P<0.001);加水处理(AW)下,密度对总生物量没有显著影响。
生长20 d 时,对照处理(CK)中,各器官生物量比率均不受密度影响;加水处理(AW)中,与低、高密度相比,中密度平均增大了根冠比21.5%(P<0.010)和根生物量比率19.4%(P<0.010),降低了叶生物量比率34.4%(P<0.05;见图2~3)。生长50 d 时,对照处理(CK)下,与低密度个体相比,高密度分别降低根冠比和根生物量比率14.1%(P=0.010)和7.5%(P=0.008),高密度提高了茎生物量比率20.3%(P=0.031),中密度降低了叶片生物量比率6.1%(P=0.024);加水处理(AW)下,与低密度下个体相比,高密度下茎生物量比率提高了45.1%(P=0.002),根冠比和根生物量比率降低了19.7%(P=0.014)和23.5%(P=0.026);在2 种水分条件下,叶柄生物量比率和繁殖器官生物量比率均不受密度影响(见图2和4)。
2.3 不同密度下生物量分配对水分的可塑响应
生长20 d 时,低密度下,与对照(CK)相比,加水处理(AW)降低了总生物量31.3%(P=0.032;见图1)。生长50 d 时,低密度和高密度下植物个体对水分的响应形成反差:与对照组(CK)相比,加水处理(AW)降低低密度下个体总生物量28.1%(P=0.014),提高了高密度下个体总生物量57.6%(P=0.034),对中密度下个体没有影响。
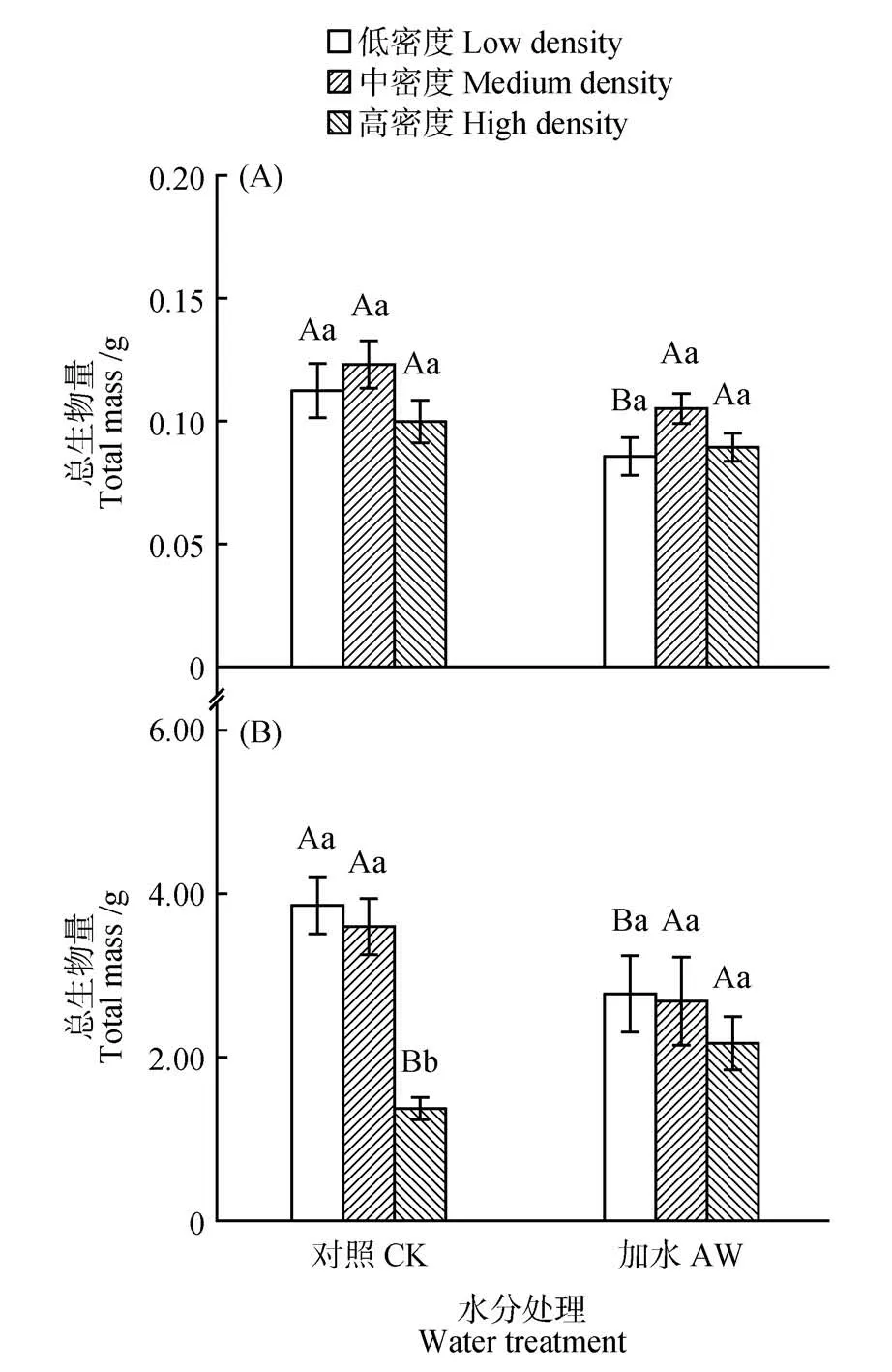
图1 植物生长20 d(A)和50 d(B)时,总生物量平均值(±标准误)对种群密度和土壤水分处理的可塑响应不同小写字母表示在相同水分条件下不同密度处理间的显著差异,不同大写字母表示相同密度下不同水分处理间的显著差异(ANCOVA,P<0.05);下同Fig.1 Mean values(±SE)of total mass in response to population density and soil water treatments at 20 d(A)and 50 d(B)of plant growth CK and AW represent control treatment(30%-50% field water capacity)and water addition treatment(70%-90% field water capacity),respectively:Different lowercase letters indicate significant differences between different density treatments within each water treatment,and different uppercase letters indicate significant differences between different water treatments within each density treatment(ANCOVA,P<0.05);The same as below
对于生物量分配,生长20 d 时,中密度下,加水处理(AW)使根冠比从0.108 提高到0.136 3(P=0.013),根生物量比率增大了23%(P=0.013),在低密度或高密度下,生物量分配不受水分处理影响(见图2~3)。生长50 d 时,在高密度下,加水处理(AW)降低了茎生物量比率(P=0.005),不影响其他密度下个体的生物量分配(见图2和4)。
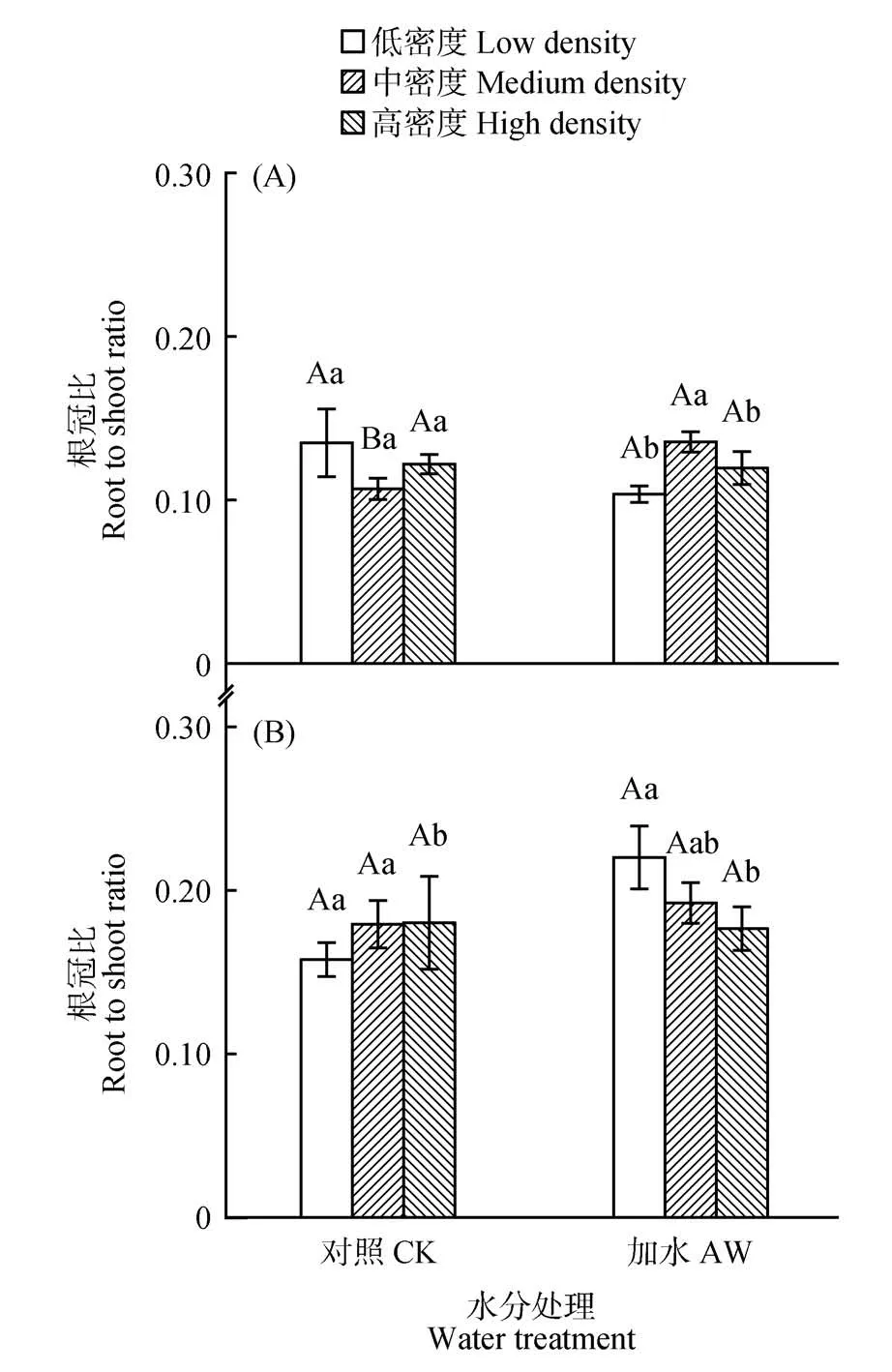
图2 植物生长20 d(A)和50 d(B)时,根冠比平均值(±标准误)对种群密度和土壤水分处理的可塑响应Fig.2 Mean values(±SE)of root/shoot ratio in response to population density and soil water treatments at 20 d(A)and 50 d(B)of plant growth
3 讨论
关于密度增大对植物生物量分配的影响,除导致茎生物量的额外增加以外,其他器官如根、叶和繁殖器官生物量分配如何响应于密度并没有非常清晰一致的结论[24-25],这可能与环境因素和植物生长阶段差异导致的密度影响程度不同有关[13,18]。本研究结果表明,水分处理对苘麻总生物量和生物量分配响应于密度的可塑性有显著影响,并且影响的方式随生长阶段而变化。植物生长20 d 时,总生物量不受密度影响,但加水处理促进了根生物量分配对中密度的积极响应(升高),加剧了叶片生物量分配对中密度的消极响应(降低),但2 种水分处理下生物量分配在高密度下均缺乏响应。这说明早期植物个体较小,地上相互作用强度很低,可忽略不计,因此中、高密度都没有降低总生物量,而中密度下适度的地下相互作用强度有利于促进根生物量分配的增加[12]。密度增大时,植物间相互作用会导致竞争和促进作用的产生,最后结果可能表现为竞争或促进作用[26-27]。生物量分配对密度的响应取决于密度增大导致的地上和地下相互作用的强度[28]。在资源充足且植物生长早期阶段,往往竞争强度较弱,植物间地下相互作用比较适中,更容易表现为促进作用,导致根生物量分配的增大[13]。与高密度相比,中密度下植物间相互作用强度更适中,更有利于地下促进作用的产生。当地下资源欠缺时,地下竞争强度加剧,抵消促进作用,导致根生物量分配不变或降低[21]。

图3 植物生长20 d 时,根(A)、茎(B)、叶(C)生物量比率平均值(±标准误)对种群密度和土壤水分处理的可塑响应Fig.3 Mean values(±SE)of root(A),stem(B)and leaf(C)mass ratio in response to population density and soil water treatments at 20 d
随着植物的生长,个体增大足以造成植物间更强烈的相互作用,并且导致地上竞争和地下竞争的加剧[29]。因此,生长阶段可以通过影响植物个体大小而影响植物相互作用强度,并由此影响生物量分配对密度增大的响应[11,13]。植物生长50 d时,在对照水分处理中,高密度降低了总生物量,但在2种水分处理中,高密度都降低了根冠比和根生物量比率,提高了茎生物量比率。说明水分欠缺加剧密度增大对植物个体大小(总生物量)的不利影响,但加水处理几乎不影响生物量分配对密度的响应。植物生长后期,竞争强度往往超过促进作用,使茎的额外伸长和根生长的降低更显著[30]。而在这个时期,加水处理中植物茎生物量分配响应于密度的可塑性更剧烈,说明加水处理促进了植物响应高密度的能力,这一点也体现为加水处理减缓了高密度对总生物量的降低。
高密度下或存在邻株植物时茎部的积极响应是一种典型的“避阴响应”[31]。高密度种群中的植物为避免相邻植株之间的遮挡,茎的额外伸长可以把叶托得更高,以获取更多的光源,对植物的存活具有重要意义[13,21]。而叶片、根等器官生物量分配对密度的响应说法并不一致[8]。生长20 d时,根生物量对中密度的响应可能以叶片生物量的降低为代价,这在生长50 d时中密度下叶片生物量的降低也有所体现,与其他研究结果[11]一致。这说明叶片生物量分配对密度增大的响应,与地上、地下竞争强度有关。高密度下,植物可能通过增大叶面积等其他叶片特征而提高叶片生物量分配[30]。
密度增大可能提高[32]、降低[33]或不影响[34]繁殖器官生物量分配。本研究中,在2个生长阶段,繁殖器官生物量均不受密度的影响,但其原始平均值(去除个体大小影响之前)随密度升高而降低,说明繁殖器官生物量主要受个体大小的影响,属于表观可塑性[35]。繁殖器官生物量分配对密度增大的响应可能也与植物间相互作用的强度有关,由于繁殖器官的重要性,植物会尽最大努力维持繁殖器官生物量分配的稳定[11,36],但当密度增大导致的地上竞争过强时,也可能导致繁殖器官生物量分配的降低[36]。
密度对植物响应水分处理的可塑性的影响也随生长阶段而变化:生长20 d 时,只有低密度下个体总生物量在加水处理中低于对照处理,而生长50 d 时,只有高密度下总生物量对加水处理有积极响应。加水处理在早期对低密度下植物的不利影响,可能是由于增加土壤水分对植物造成了一定的盐碱胁迫(实验地有一定盐碱度,在未加水状态下影响较小),而中、高密度下的植物受胁迫的影响可以与水分增加导致的竞争减弱和促进作用增强相抵消。植物生长后期,水分增加对高密度种群中植物间的竞争作用的减弱更显著,可能由于自然状态下降水增加,加水处理的频率有所降低,不足以造成植物生长和响应的差异。
4 结论
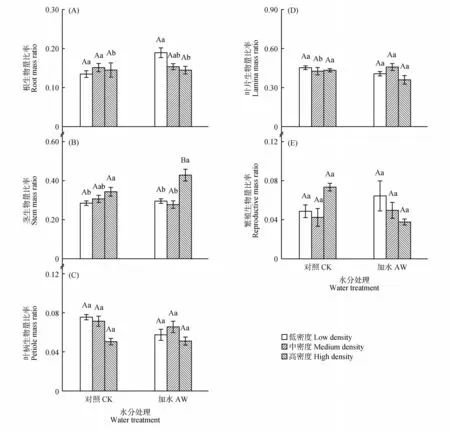
图4 植物生长50 d时,根(A)、茎(B)、叶柄(C)、叶片(D)和繁殖器官(E)生物量比率平均值(±标准误)对种群密度和土壤水分处理的可塑响应Fig.4 Mean values(±SE)of root(A),stem(B),petiole(C),lamina(D)and reproductive organ(E)mass ratio in response to population density and soil water treatments at 50 d of plant growth
本研究结果表明,水分处理对植物响应于密度的可塑性有显著影响。水分增加在植物生长早期促进根生物量分配对密度的积极响应,后期减缓密度增大对总生物量的不利影响,但对生物量分配响应于密度的可塑性的影响不显著。叶片和繁殖器官生物量分配对密度的响应与种内相互作用强度有关,说明生物量分配对密度的响应取决于种内相互作用的强度。早期适中密度下植物间相互关系更容易产生地下促进作用。生长阶段可以通过影响植物个体的大小而影响密度增大导致的植物相互作用强度,并由此影响生物量分配的响应。同时,密度增大导致的种内相互作用可以促进根生物量分配和总生物量对加水处理的积极响应。本研究在植物对生物和非生物环境因子的交互作用的响应方面进行了初步探索,相关结论可以通过未来更多研究得到进一步证实,特别是关注植物多个生长阶段的野外或温室研究。
