花期前后干旱胁迫对复水后夏玉米光合特性与产量的影响
2023-03-23张俊杰陈金平汤钰镂曹红章王丽娟马梦金王泳超郭家萌KRISHNASVJagadish杨青华邵瑞鑫
张俊杰 陈金平 汤钰镂 张 锐 曹红章 王丽娟 马梦金 王 浩 王泳超 郭家萌 KRISHNA SV Jagadish 杨青华 邵瑞鑫,*
花期前后干旱胁迫对复水后夏玉米光合特性与产量的影响
张俊杰1陈金平2汤钰镂1张 锐3曹红章1王丽娟1马梦金1王 浩1王泳超1郭家萌1KRISHNA SV Jagadish4杨青华1邵瑞鑫1,*
1河南农业大学农学院 / 省部共建小麦玉米作物学国家重点实验室 / 作物生长发育调控教育部重点实验室, 中国河南郑州 450046;2河南商丘农田生态系统国家野外科学观测研究站, 中国河南商丘 476000;3河南省农业技术推广总站, 中国河南郑州 450046;4德克萨斯州立大学植物与土壤科学系, 美国德克萨斯州拉巴克 79410
花期前后干旱是影响黄淮海地区夏玉米产量下降的重要因素之一。于2020—2021年开展池栽控制性试验, 以中科玉505为试验材料, 在开花前后28 d内设置4个水分梯度, 包括100%作物蒸散量ETC(CK)、70% ETC(干旱胁迫WD1)、40% ETC(WD2)和0 ETC(WD3), 研究花期前后干旱胁迫对夏玉米复水后的光合特性、干物质积累与分配以及产量的影响。结果表明: 干旱胁迫处理复水后夏玉米光合性能没有恢复, 表现为叶片SPAD值、净光合速率和群体叶面积指数、光合势、净同化速率低于对照, 植株干物质积累因此受阻, 导致籽粒库容能力下降。干旱胁迫处理使开花吐丝间隔期延长1~3 d, 籽粒败育率增加, 行粒数和百粒重降低, 尤其是WD3的败育率在2020—2021年分别显著增加220.71%和100.73%, WD1、WD2和WD3的产量在2020—2021年分别减产14.52%、36.69%、39.83%和19.62%、45.18%、54.42%。综上所述, 花期前后干旱胁迫在复水后, 玉米的光合性能仍受到抑制, 进而影响光合同化物的积累与分配, 最终导致库容量和产量显著下降。
花期; 干旱; 复水; 夏玉米; 光合特性; 产量
干旱是全球发生频率最高、持续时间最长、影响面积最广的自然灾害之一[1]。据统计, 极端干旱导致约9%的全球谷物产量损失[2]。玉米作为干旱敏感作物, 受旱后减产幅度可达20%~80%, 甚至绝收[3-4]。黄淮海地区作为我国最大的夏玉米主产区, 由于地下水超采严重, 加上夏玉米生长季气温高、蒸发量大、降水时空分布不均, 使玉米生长期间季节性干旱和间歇式干旱频发, 经常处于干旱和复水交替的环境中, 严重影响了夏玉米产量[5-7]。
作物产量的形成实质是源库流协调作用的结果,由库的大小、源光合产物的生产与运输能力决定[8]。研究表明: 干旱胁迫通过气孔(如气孔关闭和气孔导度下降)或非气孔(如代谢障碍、光系统被破坏和光合作用关键酶活性降低)抑制光合CO2同化, 降低光合作用, 从而导致源强度降低[9-11]。干旱胁迫通过影响个体光合性能进一步影响群体冠层光合速率, 减少同化物向籽粒中的转运分配, 降低产量[12]。前人研究表明复水后的补偿生长是植株抵御干旱胁迫的重要机制, 气孔阻力及光合活性的恢复是补偿生长的物质基础[5]。但是, 不同程度的干旱胁迫在复水后植株的补偿生长表现不同: 轻度干旱复水后作物的光合气体交换能力可逐步恢复, 然而重度干旱复水后叶片光合作用不能完全恢复[13]。
大量研究表明不同生育阶段干旱胁迫对玉米产量的影响不同[10-11]。苗期干旱虽然植株生长缓慢, 但复水后玉米产量受影响较小, 且适度的干旱胁迫会增强其抗旱性[5,14-15]。而拔节至开花期是玉米需水的旺盛期, 也是决定有效穗数、授精花数及穗大小的关键时期[3,16-17]。此时期发生干旱可抑制穗原基的伸长, 导致成对小穗的异常分化、小花退化和花粉粒发育不健全[17-18], 最终库容和籽粒产量降低[8,18]。于吐丝至灌浆期发生的干旱胁迫, 雌穗花丝抽出延迟, 源器官的碳水化合物输出能力被抑制, 导致玉米籽粒灌浆不足、籽粒充实指数下降, 产量下降[8,18,19-21]。综上, 前人研究大多从苗期、穗期、灌浆期等生育时期开展干旱胁迫对处理期间光合特性及产量的影响。但是, 花期前后长期干旱胁迫对复水后夏玉米光合性能、物质积累分配及产量的影响鲜有报道。
因此, 本试验在水分控制条件下, 根据多年夏玉米生长需水量估算花期前后的灌溉量, 通过测定开花吐丝间隔期、复水后穗位叶的光合特性和群体的光合性能及籽粒灌浆期的库容能力, 从而系统研究干旱胁迫后复水光合特性的动态变化对产量形成的影响, 以期为夏玉米抗旱栽培提供理论依据。
1 材料与方法
1.1 试验区概况
于2020—2021年, 在河南商丘农田生态系统国家野外科学观测研究站(35°31′N, 115°35′E, 海拔高度为50.70 m)的移动式防雨棚下进行。试验区属暖温带亚湿润季风性气候, 多年平均降水量为708.00 mm。2020年试验期间夏玉米生长季降水量592.40 mm, 平均最高气温和最低气温分别为31.37℃和20.88℃ (图1-A)。2021年试验期间夏玉米生长季降水量721.80 mm, 平均最高气温和最低气温分别为32.45℃和22.65℃ (图1-B)。试验期间的有效降水量和气温如图1所示。试验采用池栽, 小区面积为6.60 m2(2.00 m×3.30 m), 深度为1.80 m, 下部设20.00 cm的沙石滤层, 土层深度为1.50 m, 前茬作物为冬小麦。测坑内土壤质地为黏质壤土。0~40 cm平均土壤体积质量为1.36 g cm–3, 田间持水率为24.50%, 耕层土壤有机质和全氮量分别为13.60 g kg–1和1.27 g kg–1, 碱解氮、速效磷和速效钾量分别为56.40、10.50和52.60 mg kg–1。
1.2 试验设计
选用中科玉505为试验材料。依据河南商丘生态站夏玉米生长期间近10年平均需水量, 根据α值法计算作物蒸发蒸腾量(ETC)即作物需水量[22]。于14片叶完全展开时(V14)开始控水, 每隔7 d进行灌溉, 分3次灌溉, 设置4个水分梯度: 100% ETC(CK) (正常水分处理)、70% ETC(WD1) (轻度干旱)、40% ETC(WD2) (中度干旱)和0 ETC(WD3, 即不灌) (重度干旱), 播种密度67,500株 hm–2, 每个处理重复3次。2020年6月10日播种, 7月16日开始控水, 3次灌溉时间分别是7月23日、7月30日和8月6日, 于8月13日复水(授粉后7 d, DAP7), 9月25日收获。2021年6月12日播种, 7月19日开始控水, 分别于7月26日、8月3日和8月10日进行3次灌溉, 8月17日复水(DAP9), 9月24日收获。2020—2021年间, 4个灌溉处理的0~60 cm土层相对含水量分别达到90%~80%、75%~65%、60%~50%和45%~35%, 控水期间仅2日气温大于35℃ (图1); 复水后按照正常需水量灌溉。另外, 根据董志强等的方法[15], 土壤含水量(%) = (原土重–烘干土重)/烘干土重×100。

图1 2020–2021年夏玉米生育时期温度及降水分布
A表示2020年夏玉米生育时期温度及降水分布; B表示2021年夏玉米生育时期温度及降水分布。
A: the temperature and precipitation distribution of maize in the summer of 2020; B: the temperature and precipitation distribution of maize in the summer of 2021.
1.3 测定方法
1.3.1 开花吐丝间隔期(ASI)的记录 抽雄前, 标记每小区生长均匀一致的植株5株, 每个处理3个小区。每天记录抽雄、吐丝和散粉的时间, 单株吐丝的时间与散粉的时间间隔即开花吐丝间隔期, 15株的平均值即该处理的开花吐丝间隔期。
1.3.2 叶绿素相对含量、光合气体交换参数和叶绿素荧光参数的测定 于玉米授粉后10、15、20、30和40 d采用便携式叶绿素仪SPAD-502 (Minolta, 日本), 每小区各取5株代表性植株, 测定穗位叶叶绿素相对含量。利用Handy-PEA (Hansatech, 美国)连续激发式荧光仪, 测定玉米穗位叶的光合性能综合指数(PIABS)。测定前先进行20 min黑暗预处理, 各处理测定重复5次。通过JIP-test理论进行相关参数计算[23-24]:Po= TRo/ABS=(1–o/m);O= ETo/TRo= (1–J);Eo= ETo/ABS = (1–o/m)·O; PIABS= RC/ABS·[Po/(1–Po)]·[O/(1–O)]。在2021年的授粉后20 d, 选取5株长势一致的植株, 采用Li-6400XT便携式光合仪(Hansatech, 英国), 于无风晴朗天气的上午10:00—12:00测定叶片净光合速率(n)和胞间CO2浓度(i)。
1.3.3 叶面积指数和光合势的测定 每小区各取5株代表性植株, 于玉米授粉后10 d, 测量叶片的叶长、叶宽, 于授粉后20 d和30 d, 记录干枯叶面积, 计算叶面积和叶面积指数[25]: 叶面积 = 长×宽× 0.75; 叶面积指数(LAI) = (单株叶面积×单位面积株数)/占地面积; 光合势(LAD) = (L1+L2)×(T2–T1)/2, L1和L2为前后2次所测的叶面积, T2–T1为2个生育时期的间隔天数[26]。
1.3.4 干物质积累的转运与分配 分别于复水期和收获期, 每个小区选择长势一致且具有代表性植株3株进行取样, 分别测定玉米茎、叶、鞘、雄穗、苞叶、穗轴、籽粒等的生物量, 置于105℃烘箱杀青30 min后, 于75℃烘干至恒重并称重。计算净同化速率与物质再利用对籽粒的贡献率: 净同化速率(g m–2d–1) = [(ln LAI2 – ln LAI1)/(LAI2–LAI1)]× [(W2–W1)/(t2–t1)][26], 式中LAI1和LAI2为前后2次测定的叶面积指数, W1和W2为前后2次测定的干物质重, t1和t2为前后2次测定的时间; 物质再利用对籽粒的贡献率 = (复水时植株干重–成熟期植株干重)/成熟期籽粒干重×100[27]。
1.3.5 籽粒库容的测定 在授粉后10 d和20 d, 每小区各取3个植株的果穗, 用容量注水法测定籽粒体积, 记录籽粒数, 计算单个籽粒的体积为籽粒库容、潜在库容(LSC) = 单位面积穗数×穗粒数×籽粒库容、籽粒充实指数(KFI) = 千粒重/(籽粒库容×1000)[28]。
1.3.6 考种计产 在玉米成熟时, 每小区各取3株代表性植株, 测量果穗穗长、穗粗、秃尖长、穗行数、行粒数、败育率、百粒重等指标。收获指数(%) = 玉米籽粒产量/玉米地上部生物量×100[3]。水分利用效率(WUE) = Y/ET, Y为籽粒产量(kg hm–2), ET为作物全生育期的耗水量(mm)[29], 耗水量根据生育期内灌水量(mm)计算[29]。
1.4 数据统计与分析
采用Microsoft Excel 2010整理数据, SPSS 23.0软件统计和分析数据, 采用单因素(one-way ANOVA)和Duncan’s法进行方差分析和多重比较(α=0.05), 用Pearson’s法对果穗性状等其他指标和产量进行相关性分析, 利用SigmaPlot 13.0软件作图。
2 结果与分析
2.1 干旱胁迫对夏玉米开花吐丝间隔期的影响
干旱胁迫处理下, 玉米的开花及吐丝均有延迟, 开花吐丝间隔期延长。2020—2021年, WD1、WD3的开花吐丝间隔期分别延长了1 d和3 d。但是, WD2的开花吐丝间隔时间分别于2020年、2021年延长了1 d和2 d (图2)。总之, 花前干旱胁迫程度越严重, 对夏玉米开花吐丝间隔期的影响越大。
2.2 干旱胁迫对夏玉米穗位叶SPAD及光合特性的影响
花期前后干旱胁迫降低了复水后穗位叶的SPAD值和PIABS(图3)。2020年, 复水后DAP10, WD2、WD3的SPAD值显著降低了10.87%、15.08%; DAP15, 仅WD3的SPAD值显著下降了12.02%; 在DAP30及DAP40, SPAD在各干旱胁迫处理下均大幅下降。2021年的SPAD值与2020年趋势大致相同, 但是WD1、WD2、WD3于DAP30和DAP40的降幅较小, 所有处理在DAP40时无显著差异(图3-A), 可能是因为2021年DAP30~DAP40降雨量比2020年多, 空气湿度高缓解了土壤干旱对玉米叶片光合作用的影响(图1)。2020年DAP10, 仅WD3的PIABS显著降低15.85%; DAP15至DAP30, WD1、WD2、WD3的PIABS值分别降低了16.23%、29.71%、36.72%, 33.41%、49.87%、63.09%和33.25%、47.03%、62.43%。2021年DAP10, WD3的PIABS显著降低了21.12%; DAP15至DAP30, WD1、WD2、WD3的PIABS值分别降低了20.48%、29.02%、30.16%, 14.79%、18.63%、32.28%和22.71%、61.82%、83.16% (图3-B)。2021年干旱胁迫处理下玉米穗位叶n显著降低,i显著增加。与对照相比, WD1、WD2和WD3处理下i分别增加1.28%、13.73%和15.48% (图3-C),n分别降低16.36%、25.92%和31.39% (图3-D), 说明长期干旱胁迫对复水后的玉米叶片光合速率有显著影响。
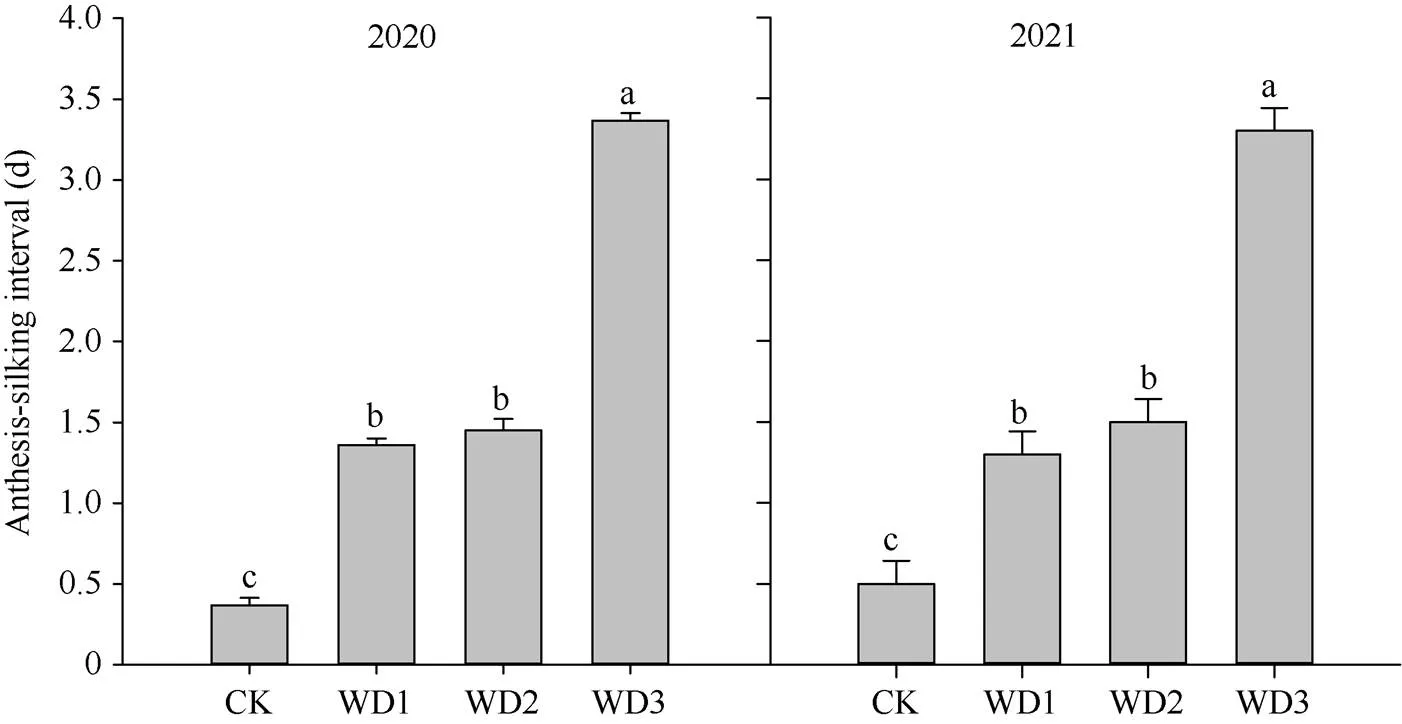
图2 花期前后干旱胁迫对夏玉米开花吐丝间隔期的影响
所有数据为平均值 ± 标准差(= 3)。图中不同小写字母表示不同处理间差异在0.05概率水平差异显著。CK: 100%作物蒸散量ETC(正常水分处理); WD1: 70% ETC(轻度干旱); WD2: 40% ETC(中度干旱); WD3: 0 ETC(重度干旱)。
Data are means ± standard deviations (SDs) (= 3). The lowercase letters in the figure indicate a significant difference of 0.05 between different treatments. CK: 100% ETC(normal water treatment); WD1: 70% ETC(mild drought); WD2: 40% ETC(moderate drought); WD3: 0 ETC(severe drought).

图3 花期前后干旱胁迫对复水后夏玉米叶片光合特性的影响
DAP表示授粉后天数; A、B分别表示2020年和2021年叶片SPAD、PIABS; C、D分别表示2021年授粉后20 d叶片的胞间CO2浓度(i)、净光合速率(n)。*表示该时期有处理和CK有显著差异。所有数据为平均值 ± 标准差 (= 3)。图中不同小写字母表示同一时期不同处理间差异在0.05概率水平差异显著。处理同图2。
DAP indicates days after pollination; A and B indicates leaf SPAD and PIABSin 2020 and 2021 respectively. C and D indicates the intercellular CO2concentration (i) and net photosynthetic rate (n) of leaves at 20 days after pollination in 2021.*indicates that there is a significant difference between treatment and CK in this period. Data are means ± SD (= 3). Different lowercase letters in the figure indicate that the difference between different treatments in the same period is significant at the 0.05 probability level. Treatments are the same as those given in Fig. 2.
2.3 干旱胁迫对夏玉米叶面积指数、光合势、净同化率的影响
合理的叶面积动态变化和物质同化效率是实现作物高产的重要保证[30]。2020年结果表明: 在DAP10, WD3的LAI显著降低了22.30%, DAP30, WD2、WD3的LAI分别显著降低46.20%和49.61%, 但是所有处理在DAP20的LAI无显著差异; 2021年与2020年的LAI不同的是, 所有处理在DAP30无显著差异, 但是在DAP10, WD2和WD3的LAI显著降低了18.30%和21.47%, 且在DAP20, 3个干旱胁迫处理的LAI都显著降低了13.67%、19.73%、27.22% (图4-A)。LAD的数据表明: 2020年仅WD3处理复水后对授粉后20~30 d的LAD有显著影响, 相对于CK下降了26.72%, 2021年WD1、WD2和WD3处理在复水后的LAD均显著降低, 授粉后10~20 d和20~30 d分别降低10.03%、17.70%、22.53%和8.29%、15.19%、26.71% (图4-B)。干旱胁迫显著降低了净同化速率, WD1、WD2、WD3处理复水后2年的平均净同化率比CK显著降低了23.41%、31.17%和42.59% (图4-C), 说明长期的干旱胁迫显著抑制了群体的光合性能。
2.4 干旱胁迫对夏玉米籽粒潜在库容与籽粒充实的影响
2020年, WD1、WD2和WD3于DAP10的籽粒库容和LSC分别显著降低了35.72%、42.20%、45.19%和30.38%、38.82%、41.98%; WD3的KFI降低了35.30%; DAP20, WD2、WD3的库容和LSC分别显著降低了24.87%、28.09%和24.87%、28.06%, 4个处理的KFI无显著差异。2021年, WD1、WD2和WD3在DAP10的籽粒库容显著降低了25.81%、29.44%、31.34%, LSC显著降低了25.81%、29.44、31.34%; WD2、WD3的KFI显著降低了21.02%、40.32%; DAP20, 4个处理的库容、LSC、KFI无显著差异。

图4 花期前后干旱胁迫对复水后夏玉米群体叶面积指数、光合势和净同化速率的影响
DAP表示授粉后天数; A表示2020年和2021年授粉后10 d、20 d和30 d的叶面积指数; B表示2020年和2021年的光合势; C表示2020年和2021年的净同化速率。*表示同一时期不同处理间具有显著差异水平(< 0.05)。所有数据为平均值 ± 标准差(= 3)。图中不同小写字母表示不同处理间在0.05概率水平差异显著。处理同图2。
DAP indicates days after pollination; A indicates leaf area index at 20 and 30 days after pollination in 2020 and 2021; B indicates photosynthetic potential in 2020 and 2021; C indicates net assimilation rate in 2020 and 2021 respectively.*represents the significant difference among different treatments in the same period (< 0.05). Data are means ± SD (= 3). The lowercase letters in the figure indicate significant differences between different treatments at the 0.05 probability level. Treatments are the same as those given in Fig. 2.

图5 花期前后干旱胁迫对复水后夏玉米籽粒库容与籽粒库容指数的影响
DAP表示授粉后天数; A表示2020年和2021年授粉后10 d、20 d的库容; B表示2020年和2021年授粉后10 d、20 d的潜在库容指数 (LSC); C表示2020年和2021年授粉后10 d、20 d的籽粒充实指数(KFI)。所有数据为平均值 ± 标准差(= 3)。图中不同小写字母表示同一时间内不同处理间差异在0.05概率水平差异显著。处理同图2。
DAP: days after pollination; A: grain storage at 10 days and 20 days after pollination in 2020 and 2021, respectively; B: latent storage capacity (LSC) at 10 and 20 days after pollination in 2020 and 2021, respectively; C: kernels filling index (KFI) at 10 days and 20 days after pollination in 2020 and 2021, respectively; Data are means ± SDs (= 3). Different lowercase letters in the figure indicate significant difference at the 0.05 probability level among the different treatments at the same time. Treatments are the same as those given in Fig. 2.
2.5 干旱胁迫对夏玉米干物质积累、分配和干物质对籽粒贡献率的影响
干旱胁迫抑制了茎、叶、鞘、苞叶、雄穗和穗轴的生物量积累(图6)。2020年的数据显示: DAP10, WD1、WD2和WD3苞叶+雄穗+穗轴的总干重和籽粒干重分别显著降低23.51%、27.66%、39.04%和25.76%、33.66%、57.00%, WD3的茎显著降低了36.02%, WD2和WD3的鞘分别显著降低14.56%和24.18%, 4个处理间的叶干重无显著差异; 收获时不同处理的生物量进行对比, WD3的茎、叶、鞘分别显著降低了22.73%、15.37%、14.61%。2021年, 与CK相比, WD1、WD2和WD3在DAP10的茎和籽粒干重显著降低12.83%、26.96、32.95%, 28.30%、45.27%、49.83%, WD3的叶显著降低了28.44%, WD2和WD3的鞘分别降低14.56%和24.18%, WD1、WD2和WD3的苞叶+雄穗+穗轴的总干重显著降低16.58%、18.12%和55.19%; 收获时, WD2、WD3的茎干重显著降低了13.35%、31.81%, WD3的苞叶+雄穗+穗轴的总干重和叶干重显著降低17.20%和21.57%, 4个处理间的鞘干重无显著差异(图6-A)。进一步通过测定复水后和收获时植株生物量的差值与籽粒干重计算叶片同化物对籽粒的贡献率, 结果表明WD1、WD2、WD3对籽粒的贡献率与CK相比分别平均降低28.59%、44.49%和69.09% (图6-B)。
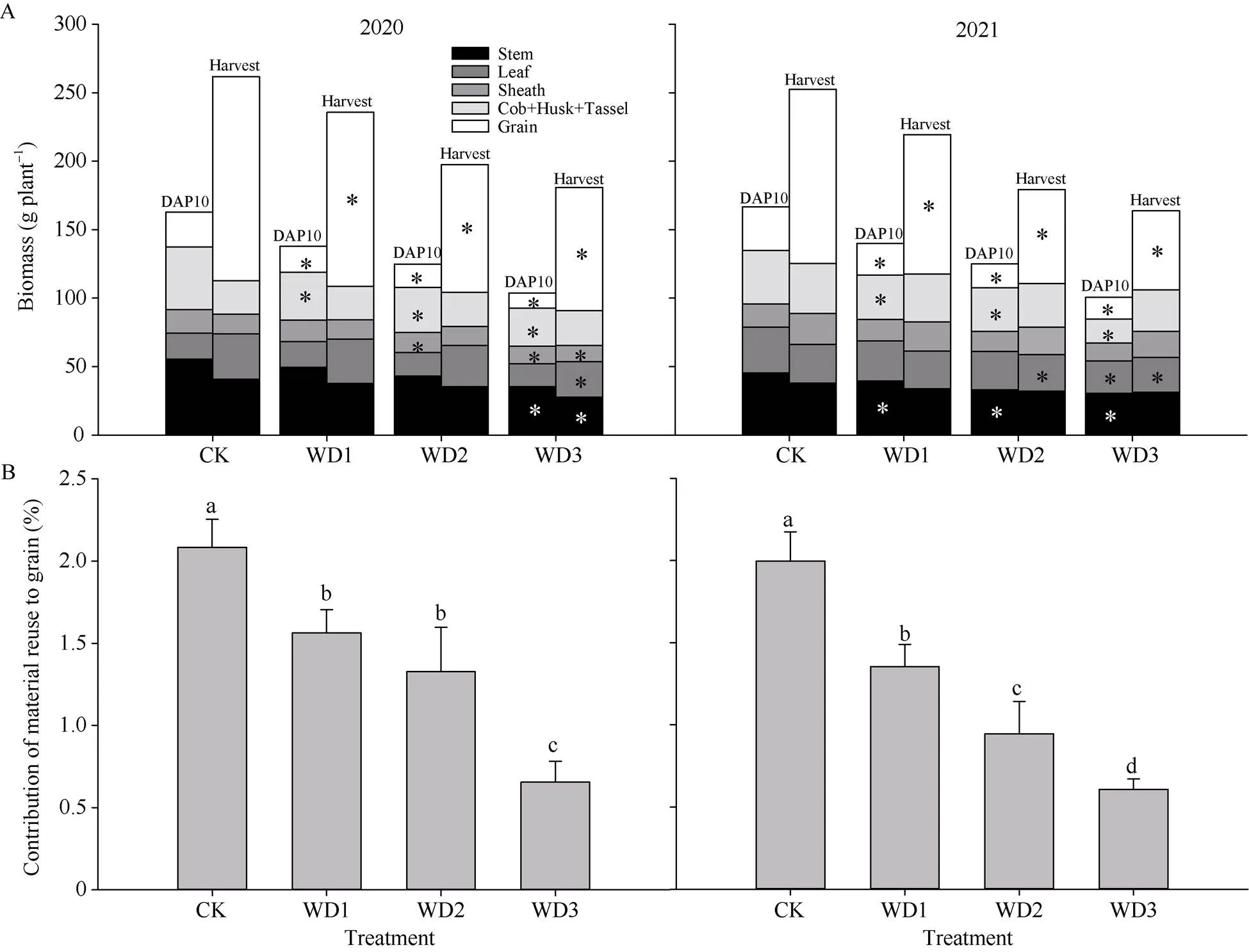
图6 花期前后干旱胁迫对干物质积累及籽粒贡献率的影响
DAP表示授粉后天数; A分别表示2020年和2021年授粉后10 d (DAP10)和收获时茎、叶、鞘、雄穗+苞叶+穗轴、籽粒的积累量; B分别表示2020年和2021年物质再利用对籽粒的贡献率。*表示同一时期不同处理间差异显著(< 0.05)。所有数据为平均值 ± 标准差(= 3)。图中不同小写字母表示不同处理间差异在0.05概率水平差异显著。处理同图2。
DAP indicates days after pollination; A indicates the accumulation of stems, leaves, sheaths, tassel + husk + cob, grain at 10 days after pollination (DAP10) and Harvest in 2020 and 2021 respectively. B indicates the contribution of material reuse to grain in 2020 and 2021 respectively; * represents the significant difference among different treatments in the same period (< 0.05). Data are means ± SDs (= 3). Different lowercase letters in the figure indicate that there is significant difference at the 0.05 probability level between different treatments. Treatments are the same as those given in Fig. 2.
2.6 干旱胁迫对夏玉米产量、产量构成因素及其相关性的影响
2020年, WD2、WD3的穗长、行粒数和穗粒数比CK分别显著降低7.27%、9.15%, 20.79%、20.11%和21.06%、25.28%。WD1、WD2、WD3的穗粗、百粒重、产量、收获指数、WUE分别显著降低2.65%、4.63%、6.74%, 8.99%、9.66%、16.15%, 14.52%、36.69%、39.83%, 14.81%、24.07%、33.33%和13.78%、37.01%、39.85%, 败育率显著增加82.17%、103.57%、220.71%, 但是穗行数和秃尖长在4个处理间无显著差异。2021年, WD2、WD3的穗长、穗粗分别显著降低9.71%、17.93%和8.31%、6.28%, 秃尖长显著增加193.88%、259.18%。WD1、WD2、WD3的行粒数、穗粒数、百粒重、产量、收获指数、WUE分别显著降低8.32%、23.40%、24.89%, 12.11%、27.35%、40.75%, 18.86%、23.42%、30.65%, 19.62%、45.18%、54.42%, 6.82%、15.91%、43.18%和19.32%、45.79%、54.61%, 败育率显著增加33.19%、44.95%、100.73%, 4个处理的穗行数无显著差异(表1)。
为了分析长期干旱胁迫条件下夏玉米产量与光合性能等其他指标之间的相关性, 进一步对2020—2021年夏玉米产量构成因素与光合特性、生物量积累及籽粒库容等指标进行了相关性分析。2020年的数据显示(表2), 穗粒数与行粒数和穗长呈极显著正相关, 与败育率呈极显著的负相关; 百粒重与籽粒库容、LAI、光合势、总生物量和穗粒数呈极显著的正相关, 与ASI、败育率呈极显著的负相关;产量与DAP10的籽粒库容、穗粒数和百粒重呈极显著的正相关, 与ASI和败育率呈极显著的负相关。2021年数据(表3)表明, 穗粒数与籽粒库容、LAI、光合势、总生物量、穗长和行粒数呈极显著的正相关, 与ASI、i和败育率呈极显著的负相关; 百粒重与籽粒库容、n、LAI、光合势、穗粒数和总生物量呈极显著的正相关, 与败育率呈极显著的负相关; 产量与n、LAI、光合势、总生物量、行粒数、穗粒数、穗长和百粒重呈极显著的正相关, 与i和败育率呈极显著的负相关。
3 讨论
叶绿素是光合作用的重要色素, 在光能吸收和运输中起着关键作用[31-32]。本研究中, 干旱处理后, 叶片的SPAD值显著降低, 暗示长时期的干旱胁迫加速了叶绿素的降解或者减少了叶绿素的合成, 叶片对光能的吸收和利用受到影响, 进而导致n显著下降。叶绿素荧光参数PIABS也是以光能吸收为基础, 综合反映光合机构活性的参数[11]。干旱复水后叶片PIABS的下降程度与干旱程度有关, 说明中度以上干旱胁迫对PSII反应中心破坏严重, 用于光能捕获和电子传递的活性降低[11,17]。同时,i值并没有因此下降, 说明长期的干旱胁迫对光合速率的影响是由非气孔因素中叶绿素含量和叶肉细胞的光化学活性降低所致[33]。另外, 本文中这种非气孔限制所致的低光合能力在复水后没有恢复至正常水平, 下降程度与干旱胁迫发生的时期和程度有关。
叶面积指数、光合势、净同化率反映了群体光合生产能力的重要表征[34-36]。本文中, 干旱复水后LAI、LAD和净同化率显著下降, 源强度因此显著降低, 表明长期的干旱胁迫导致玉米群体的光合性能受到严重影响[13]。作物干物质积累、运输分配以及转化的过程实际上就是其产量的形成过程[11,21]。作物高产不仅需要较强的光合能力, 还要求各营养器官的光合产物能够有效运输与分配[12,35], 尤其花前所贮藏的干物质向籽粒的转运与分配对谷类作物产量形成有重要作用[37]。但是, 干旱对群体光合性能的抑制影响了营养器官光合同化物的积累及向籽粒中的运输[13,18,27]。且复水后发生的新组织对干旱的补偿效应决定了作物的最终产量[33]。本研究中, 干旱复水发生在叶片开始衰老的时期, 因此即使复水也难以补偿干旱对产量形成的制约, 表现为复水后籽粒库容、LSC和KFI均低于对照, 说明花期前后长期的干旱胁迫制约了源光合同化产物的积累及其向籽粒中的供应[38-39]。
玉米产量构成因素中, 穗粒数、粒重对土壤供水亏缺程度比较敏感[40]。本试验中, 花期前后长期的干旱胁迫对穗行数无显著影响, 与干旱处理时小穗分化已完成有关, 而穗粒数和百粒重显著下降, 中度及重度干旱条件下穗粒数的降幅高于百粒重。前人研究表明, 吐丝前后干旱胁迫使ASI延长主要是由于玉米雌穗吐丝时间延迟, 且雌穗上部花丝最为明显, 但是对中下部花丝抽出和雄穗散粉时间影响较小[41]。本文表明, 花期前后干旱引起秃尖长增加、穗粒数减少不仅与ASI延长有关, 还因雌穗上部受精的小花数降低和受精后的子房滞育导致籽粒败育; 另外, 由于干旱后复水补偿效应不足, 又因籽粒库容降低不能贮存较多的干物质导致粒重降低。
4 结论
花期前后长期干旱胁迫加速了功能叶片的早衰,显著影响了复水后玉米群体的源光合能力和库的构建, 表现为叶片叶绿素含量、光合气体交换参数、光合性能综合指数的下降及群体叶面积指数、光合势的下降, 严重影响源器官光合同化物的积累及其对籽粒的贡献率; 同时, 籽粒形成期籽粒库容的下降也限制了同化物对籽粒的贡献率, 最终使成熟期玉米粒重显著降低。干旱后开花吐丝间隔期的延长, 导致了败育率的增加和穗粒数的显著下降。花期前后长期干旱胁迫影响源与库之间的关系, 由于干旱影响的滞后性, 即使复水也不能恢复, 且对产量的影响与干旱程度有关。

[1] Zhao T B, Dai A G. The magnitude and causes of global drought changes in the twenty-first century under a low-moderate emissions scenario., 2015, 28: 4490–4512.
[2] Brás T A, Seixas J, Carvalhais N, Jägermeyr J. Severity of drought and heatwave crop losses tripled over the last five decades in Europe., 2021, 16: 65012–65025.
[3] Sah R P, Chakraborty M, Prasad K, Pandit M, Tudu V K, Chakravarty M K, Narayan S C, Rana M, Moharana D. Impact of water deficit stress in maize: phenology and yield components., 2020, 11: 2944–2958.
[4] 于振文. 作物栽培学各论北方本. 北京: 中国农业出版社, 2013. pp 69–111.
Yu Z W. Monographs on Northern Origin of Crop Cultivation. Beijing: China Agriculture Press, 2013. pp 69–111 (in Chinese).
[5] 赵成凤, 王晨光, 李红杰, 郑学慧, 杨梅, 张仁和. 干旱及复水条件下外源褪黑素对玉米叶片光合作用的影响. 生态学报, 2021, 41: 1431–1439.
Zhao C F, Wang C G, Li H J, Zheng X H, Yang M, Zhang R H. Effects of exogenous melatonin on photosynthesis of maize leaves under drought stress and re-watering., 2021, 41: 1431–1439 (in Chinese with English abstract).
[6] Liu S L, Wu W B, Yang X G, Yang P, Sun J. Exploring drought dynamics and its impacts on maize yield in the Huang-Huai-Hai farming region of China., 2020, 163: 415–430.
[7] Hu Z H, Wu Z R, Zhang Y X, Li Q, Islam A R M T, Pan C C. Risk assessment of drought disaster in summer maize cultivated areas of the Huang-Huai-Hai plain, eastern China., 2021, 193: 441–455.
[8] Fernie A R, Bachem C W B, Helariutta Y, Neuhaus H E, Prat S, Ruan Y L, Stitt M, Sweetlove L J, Tegeder M, Wahl V, Sonnewald S, Sonnewald U. Synchronization of developmental, molecular and metabolic aspects of source-sink interactions., 2020, 6: 55–66.
[9] Song X Y, Zhou G S, He Q J. Critical leaf water content for maize photosynthesis under drought stress and its response to re-watering.. 2021, 13: 7218–7232.
[10] Wang Y F, Guo Y Y, Zhao C F, Li H J, Zhang R H. Exogenous melatonin achieves drought tolerance by improving photosynthesis in maize seedlings leaves., 2021, 68: 718–727.
[11] 贾双杰. 穗期干旱胁迫对玉米光合性能和雌花序发育的影响. 河南农业大学硕士学位论文, 河南郑州, 2020.
Jia S J. Effect of Drought Stress on Photosynthetic Performance and Female Inflorescence Development of Maize in Panicle Stage. MS Thesis of Henan Agricultural University, Zhengzhou, Henan, China, 2020 (in Chinese with English abstract).
[12] 王佳旭, 王宏伟, 姜文野, 赵彬, 满艳苹, 张旷野, 刁玉霖, 朱康宁. 不同种植方式对玉米干物质积累、分配和产量的影响. 玉米科学, 2021, 29(5): 128–136.
Wang J X, Wang H W, Jiang W Y, Zhao B, Man Y P, Zhang K Y, Diao Y L, Zhu K N. Effect of different planting patterns on dry matter accumulation, distribution and yield of maize., 2021, 29(5): 128–136 (in Chinese with English abstract).
[13] Jia Y Y, Xiao W X, Ye Y S, Wang X L, Liu X L, Wang G H, Li G, Wang Y B. Response of photosynthetic performance to drought duration and re-watering in maize., 2020, 10: 533–549.
[14] 董智强, 李曼华, 李楠, 薛晓萍, 陈辰, 张继波, 赵红, 侯英雨,潘志华. 山东夏玉米土壤干旱阈值研究与影响评价. 中国农业科学, 2020, 53: 4376–4387.
Dong Z Q, Li M H, Lin N, Xue X P, Chen C, Zhang J B, Zhao H, Hou Y Y, Pan Z H. The thresholds of soil drought and its impacts on summer maize in Shandong province., 2020, 53: 4376–4387 (in Chinese with English abstract).
[15] Zhang X C, Myrold D D, Shi L L, Kuzyakov Y, Dai H C, Hoang D T T, Dippold M A, Meng X T, Song X N, Li Z Y, Zhou J, Razavi B S. Resistance of microbial community and its functional sensitivity in the rhizosphere hotspots to drought., 2021, 161: 108360–108371.
[16] Shao R X, Jia S J, Tang Y L, Zhang J J, Li H W, L L P, Chen J H, Guo J M, Wang H, Yang Q H, Wang Y C, Liu T X, Zhao X. Soil water deficit suppresses development of maize ear by altering metabolism and photosynthesis., 2021, 192: 104651–104660.
[17] 贾双杰, 李红伟, 江艳平, 赵国强, 王和洲, 杨慎骄, 杨青华, 郭家萌, 邵瑞鑫. 干旱胁迫对玉米叶片光合特性和穗发育特征的影响. 生态学报, 2020, 40: 854–863.
Jia S J, Li H W, Jiang Y P, Zhao G Q, Wang H Z, Yang S J, Yang Q H, Guo J M, Shao R X. Effects of drought on photosynthesis and ear development characteristics of maize., 2020, 40: 854–863 (in Chinese with English abstract).
[18] Li Y B, Tao H B, Zhang B C, Huang S B, Wang P. Timing of water deficit limits maize kernel setting in association with changes in the source-flow-sink relationship., 2018, 9: 1326–1336.
[19] Li F W, Zhang M J, Liu Y Z. Quantitative research on drought loss sensitivity of summer maize based on AquaCrop model., 2022, 112: 1065–1084.
[20] Liu X W, Yu Y H, Huang S B, Xu C C, Wang X Y, Gao J, Meng Q F, Wang P. The impact of drought and heat stress at flowering on maize kernel filling: Insights from the field and laboratory., 2022, 312: 108733–108744.
[21] Shemi R, Wang R, Gheith, El-Sayed M S, Hussain H A, Hussain A, Muhammad I, Cholidah L, Zhang K, Zhang S, Wang L C. Effects of salicylic acid, zinc and glycine betaine on morpho- physiological growth and yield of maize under drought stress., 2021, 11: 3195–3207.
[22] Tigkas D, Vangelis H, Tsakiris G. Implementing crop evapotranspiration in RDI for farm-level drought evaluation and adaptation under climate change conditions., 2020, 34: 4329–4343.
[23] 李鹏民, 高辉远, Strasser R J. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用. 植物生理与分子生物学学报, 2005, 31: 559–566.
Li P M, Gao H Y, Strasser R J. Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study., 2005, 31: 559–566 (in Chinese with English abstract).
[24] Xue H Y, Wang S F, Zhang X, Zhang Z Y. The rapid chlorophyll a fluorescence characteristics of different cotton genotypes reflect differences in leaf senescence., 2021, 29: 870–879.
[25] 柏延文, 杨永红, 朱亚利, 李红杰, 薛吉全, 张仁和. 种植密度对不同株型玉米冠层光能截获和产量的影响. 作物学报, 2019, 45: 1868–1879.
Bai Y W, Yang Y H, Zhu Y L, Li H J, Xue J Q, Zhang R H. Effect of planting density on light interception within canopy and grain yield of different plant types of maize., 2019, 45: 1868–1879 (in Chinese with English abstract).
[26] 马正波, 董学瑞, 唐会会, 闫鹏, 卢霖, 王庆燕, 房孟颖, 王琦, 董志强. 四甲基戊二酸对夏玉米光合生产特征的调控效应. 作物学报, 2020, 46: 1617–1627.
Ma Z B, Dong X R, Tang H H, Yan P, Lu L, Wang Q Y, Fang M Y, Wang Q, Dong Z Q. Effect of tetramethyl glutaric acid on summer maize photosynthesis characteristics., 2020, 46: 1617–1627 (in Chinese with English abstract).
[27] Molla S H, Nakasathien S, Ali A, Kha A, Alam R, Hossain A, Farooq M, Sabagh M F. Influence of nitrogen application on dry biomass allocation and translocation in two maize varieties under short pre-anthesis and prolonged bracketing flowering periods of drought., 2019, 65: 928–944.
[28] 李宁, 段留生, 李建民, 翟志席, 李召虎. 播期与密度组合对不同穗型小麦品种花后旗叶光合特性、籽粒库容能力及产量的影响. 麦类作物学报, 2010, 30: 296–302.
Li N, Duan L S, Li J M, Zhai Z X, Li Z H. Effect of sowing date and planting density on flag leaf photosynthesis, storage capacity and yield in different spike type varieties., 2010, 30: 296–302 (in Chinese with English abstract).
[29] 蔡晓, 王东, 吴祥运, 吴雨晴, 林祥, 张俊鹏. 氮肥减施对夏玉米生长及水氮利用效率的影响. 玉米科学, 2022, 30(1): 158–165.
Cai X, Wang D, Wu X Y, Wu Y Q, Lin X, Zhang J P. Effects of nitrogen reduction on growth and water-nitrogen use efficiency of summer maize., 2022, 30(1): 158–165 (in Chinese with English abstract).
[30] 杨慧. 不同耕法和密度对春玉米产量及根系的调控研究. 东北农业大学硕士学位论文, 黑龙江哈尔滨, 2018.
Yang H. The Regulation of Yield and Roots Formations through Different Tillage Method and Density Treatments. MS Thesis of Northeastern Agricultural University, Harbin, Heilongjiang, China, 2018 (in Chinese with English abstract).
[31] Qi M, Liu X D, Li Y B, Song H, Yin Z T, Zhang F, He Q J, Xu Z Z, Zhou G S. Photosynthetic resistance and resilience under drought, flooding and re-watering in maize plants., 2021, 148: 1–15.
[32] Simkin A J, Kapoor L, Doss C G P, Hofmann T A, Lawson T, Ramamoorthy S. The role of photosynthesis related pigments in light harvesting, photoprotection and enhancement of photosynthetic yield in planta., 2022, 152: 23–42.
[33] 夏璐, 赵蕊, 王怡针, 金海燕, 吴锡冬, 葛均筑, 臧凤艳, 李子芳, 王金龙. 干旱胁迫对夏玉米光合作用和叶绿素荧光特性的影响. 华北农学报, 2019, 34(3): 102–110.
Xia L, Zhao R, Wang Y Z, Jin H Y, Wu X D, Ge J Z, Zang F Y, Li Z F, Wang J L. Effect of drought stress on photosynthesis and chlorophyll fluorescence characteristics of summer maize., 2019, 34(3): 102–110 (in Chinese with English abstract).
[34] 李静, 王洪章, 刘鹏, 张吉旺, 赵斌, 任佰朝. 夏玉米不同栽培模式花后叶片光合性能的差异. 作物学报, 2021, 47: 1351–1359.
Li J, Wang H Z, Liu P, Zhang J W, Zhao B, Ren B C. Differences in photosynthetic performance of leaves at post-flowering stage in different cultivation modes of summer maize., 2021, 47: 1351–1359 (in Chinese with English abstract).
[35] 穆心愿, 夏来坤, 谷利敏, 张凤启, 张君, 丁勇, 齐建双, 唐保军, 赵发欣, 邢建伟. 花期干旱胁迫对不同夏玉米品种花后干物质积累运转及产量的影响. 南方农业学报, 2021, 52: 931–941.
Mu X Y, Xia L K, Gu L M, Zhang F Q, Zhang J, Ding Y, Qi J S, Tang B J, Zhao F X, Xing J W. Effects of drought stress during flowering on post-flowering dry matter accumulation and transfer and yield of different maize cultivars., 2021, 52: 931–941 (in Chinese with English abstract).
[36] Jahangirlou M R, Akbari G A, Alahdadi I, Soufizadeh S, Parsons D. Phenotypic traits, grain yield and yield components of maize cultivars under combinations of management practices in semi-arid conditions of Iran., 2021, 15: 459–471.
[37] Yu Y, Qian C R, Gu W R, Li C F. Responses of root characteristic parameters and plant dry matter accumulation, distribution and transportation to nitrogen levels for spring maize in Northeast China., 2021, 11: 308–331
[38] 胡旦旦, 李荣发, 刘鹏, 董树亭, 赵斌, 张吉旺, 任佰朝. 密植条件下玉米品种混播提高籽粒灌浆性能和产量. 中国农业科学, 2021, 54: 1856–1868.
Hu D D, Li R F, Liu P, Dong S T, Zhao B, Zhang J W, Ren B C. Mixed-cropping improved on grain filling characteristics and yield of maize under high planting densities., 2021, 54: 1856–1868 (in Chinese with English abstract).
[39] Lucas B, Lucas N V M. Maize reproductive development and kernel set under limited plant growth environments., 2018, 69: 3235–3243.
[40] 高震, 梁效贵, 张莉, 赵雪, 杜雄, 崔彦宏, 周顺利. 不同时期灌溉对华北平原春玉米穗粒数的影响. 作物学报, 2021, 47: 1324–1331.
Gao Z, Liang X G, Zhang L, Zhao X, Du X, Cui Y H, Zhou S L. Effects of irrigating at different growth stages on kernel number of spring maize in the North China Plain., 2021, 47: 1324–1331 (in Chinese with English abstract).
[41] 李叶蓓, 陶洪斌, 王若男, 张萍, 吴春江, 雷鸣, 张巽, 王璞. 干旱对玉米穗发育及产量的影响. 中国生态农业学报, 2015, 23: 383–391.
Li Y B, Tao H B, Wang R N, Zhang P, Wu C J, Lei M, Zhang X, Wang P. Effects of drought on ear development and yield of maize., 2015, 23: 383–391 (in Chinese with English abstract).
Effects of drought stress before and after anthesis on photosynthetic characteristics and yield of summer maize after re-watering
ZHANG Jun-Jie1, CHEN Jin-Ping2, TANG Yu-Lou1, ZHANG Rui3, CAO Hong-Zhang1, WANG Li-Juan1, MA Meng-Jin1, WANG Hao1, WANG Yong-Chao1, GUO Jia-Meng1, KRISHNA SV Jagadish4, YANG Qing-Hua1, and SHAO Rui-Xin1,*
1Agronomy College of Henan Agricultural University / National Key Laboratory of Wheat and Maize Crop Science / Key Laboratory of Regulating and Controlling Crop Growth and Development, Ministry of Education, Zhengzhou 450046, Henan, China;2National Field Science Observation and Research Station of Shangqiu Agricultural Ecology, Shangqiu 476000, Henan, China;3Henan Province Agricultural Technology Extension General Station, Zhengzhou 450046, Henan, China;4Department of Plant and Soil Science, Texas Tech University, Lubbock 79410, Texas, USA
Drought before and after anthesis is one of the important factors affecting the decrease of summer maize yield in Huang-Huai-Hai Rivers region. From 2020 to 2021, a pool planting control experiment was carried out. To study the effect of regulated deficit irrigation before and after anthesis on photosynthetic characteristics, dry matter accumulation and distribution, and yield of summer maize after re-watering, four water gradients were set within 28 days before and after anthesis, including 100% Eapotranspiration (ETc) (CK), 70% ETC(Water deficit, WD1), 40% ETC(WD2), and 0 ETC(WD3) using Zhongkeyu 505 as the test materials. The result showed that the photosynthetic performance of summer maize leaves did not recover after re-watering under drought stress. The SPAD value, net photosynthetic rate, population leaf area index, photosynthetic potential and net assimilation rate were lower than the control, and the dry matter accumulation of the plants was blocked, photosynthetic potential and net assimilation rate were lower than the control, and the dry matter accumulation of the plants was blocked, resulting in the decrease of grain storage capacity. Drought stress prolonged the anthesis-silking interval for 1–3 day (s), increased the seed abortion rate, and decreased the grain number per row and 100-seed weight after re-watering, especially the seed abortion rate in WD3 was significantly increased by 220.71% and 100.73% in 2020 and 2021, respectively. The yield of WD1, WD2, WD3 was decreased by 14.52%, 36.69%, 39.83% and 19.62%, 45.18%, 54.42% in 2020 and 2021, respectively. In conclusion, after re-watering under water deficit before and after anthesis, the photosynthetic performance of maize was still inhibited, which further affected the accumulation and distribution of photosynthetic assimilates, and ultimately leading to a significant decrease in storage capacity and yield.
anthesis; drought; summer maize; re-watering; photosynthetic characteristics; yield
10.3724/SP.J.1006.2023.23003
本研究由河南省高校科技创新人才支持计划(20HASTIT036), 国家重点研发计划项目子课题(2021YFD1901002-8)和中原英才计划——中原青年拔尖人才项目资助。
This study was supported by the Henan University Science and Technology Innovation Talents Support Plan (20HASTIT036), the Sub-project of the National Key Research and Development Program of China (2021YFD1901002-8), and the Central Plains Talents Program—Central Plains Youth Top Talents.
邵瑞鑫, E-mail: shao_rui_xin@126.com
E-mail: zjj3021435607@163.com
2022-01-07;
2022-10-10;
2022-10-26.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20221025.1144.002.html
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
