结直肠癌组织中miR-876-5p、PAX6 mRNA表达变化及意义
2023-03-17徐杰张树朋李照晋邵建平
徐杰,张树朋,李照晋,邵建平
天津市第五中心医院普外科,天津 300450
据全球癌症统计数据显示,2020 年我国结直肠癌新发病例为55.5 万例,死亡病例28.6 万例[1];预计2022年新发病例和死亡病例分别为59.2、30.9万例[2]。尽管近年来结直肠癌的诊治取得了一定进展,但患者预后仍然较差,因此还需进一步探索结直肠癌发病的分子机制。研究表明,非编码RNA 参与结直肠癌进展[3]。微小RNA(miRNA)是一种小分子非编码RNA,miRNA-876-5p 是miRNA 家族的一个新成员。miR-876-5p 参与髓母细胞瘤、肝癌等恶性肿瘤的发生、发展[4-5]。配对框基因6(PAX6)是一种多功能转录因子,能通过调节下游靶基因转录参与多种恶性肿瘤细胞增殖、分化、侵袭、凋亡[6-7]。本研究通过TargetScan 数据库预测发现,miR-876-5p 与PAX6 存在结合位点,但关于结直肠癌组织中miR-876-5p、PAX6 表达变化及意义尚缺乏报道。基于此,本研究做了相关探讨,现报告如下。
1 资料与方法
1.1 临床资料 选取2018年1月—2019年10月本院收治的86例结直肠癌患者,均经术后病理学检查确诊。其中男51 例,女35 例;≥60 岁41 例、<60 岁45例;肿瘤部位:直肠55 例、结肠31 例;肿瘤最大径≥5 cm 24 例、<5 cm 62 例;分化程度:低分化35 例、中高分化51 例;TNM 分期[8]:Ⅰ期17 例Ⅱ期、21 例、Ⅲ期48 例;有淋巴结转移35 例、无淋巴结转移31 例。纳入标准:初诊且入院前未接受任何抗肿瘤治疗;TNM 分期Ⅰ~Ⅲ期;接受姑息性或根治性手术;患者或家属签署知情同意书。排除标准:合并克罗恩病、溃疡结肠炎等其他肠道疾病;合并全身性感染性疾病、自身免疫性疾病、其他部位恶性肿瘤;年龄<18 岁;临床资料不完整或不能接受随访;院内死亡。本研究经医院伦理委员会批准。
1.2 miR-876-5p 与PAX6 的关系预测 用TargetScan 数据库(https://www. targetscan. org/)预测miR-876-5p与PAX6的关系。
1.3 结直肠癌组织和癌旁组织中miR-876-5p、PAX6 mRNA 表达检测 采用RT-qPCR 法。术中留取部分癌组织和癌旁组织(距肿瘤边缘>2 cm)用液氮冷冻后研磨成粉末,TRIzol法提取组织总RNA,微量分光光度计验证纯度、浓度合格后使用Takara 反转录试剂盒(上海翌圣生物科技股份有限公司)合成cDNA。参考SYBR®Premix Ex TaqTM试剂盒(广州威佳科技有限公司)说明书进行扩增,引物设计和合成由上海生工生物工程股份有限公司完成。miR-876-5p 引物:上游 5′-ACACTCCAGCTGGGTTTGTGACCTGGTCCA-3′,下游5′-TGGTGTCGTGGAGTCGTGTCGTGGAGTC-3′;内参U6引物:上游5′-CGCTTCGGCAGCACATATACTA-3′,下游5′-GCGAGCACAGAATTAATACGAC-3′;PAX6 引物:上游5′-TCTTTGCTTGGGAAATCCG-3′,下游5′-CTGCCCGTTCAACATCCTTAG-3′;内参GAPDH 引物:上游5′-GGCACTGAGAAGCGGGGCCG-3′,下游5′-GAAGATGGTGATGGGATTTC-3′。 反应条件:95 ℃ 90 s;95 ℃ 30 s,63 ℃ 30 s,72 ℃ 15 s,循环40次。用2-ΔΔCT法计算miR-876-5p、PAX6 mRNA 相对表达量。
1.4 预后分析 患者出院后通过门诊或电话随访3年,随访至2022年10月或患者死亡,计算3年累积生存率。根据结直肠癌组织中miR-876-5p、PAX6 mRNA表达水平将患者分为miR-876-5p高表达组(≥0.59,n=42)、miR-876-5p 低表达组(<0.59,n=44),PAX6 mRNA 高表达组(≥0.98,n=47)、PAX6 mRNA低表达组(<0.98,n=39)。
1.5 统计学方法 采用SPSS28.0 统计软件。符合正态分布的计量资料以±s表示,两组比较采用t检验;计数资料比较采用χ2检验;结直肠癌组织中miR-876-5p 与PAX6 mRNA 表达的相关分析用Pearson 相关法;绘制结直肠癌患者生存曲线,组间生存率比较用Log-rank 检验。P<0.05 为差异有统计学意义。
2 结果
2.1 结直肠癌组织与癌旁组织中miR-876-5p、PAX6 mRNA表达比较 结直肠癌组织中miR-876-5p相对表达量低于癌旁组织,PAX6 mRNA相对表达量高于癌旁组织(P均<0.05)。见表1。
表1 结直肠癌组织与癌旁组织中miR-876-5p、PAX6 mRNA表达比较(±s)

表1 结直肠癌组织与癌旁组织中miR-876-5p、PAX6 mRNA表达比较(±s)
组织癌组织癌旁组织P n 86 86 miR-876-5p 0.59 ± 0.11 0.98 ± 0.17<0.05 PAX6 mRNA 0.98 ± 0.15 0.52 ± 0.08<0.05
2.2 结直肠癌组织中miR-876-5p 与PAX6 mRNA表达的相关性 经在线网站https://www. targetscan. org/预测,miR-876-5p 与PAX6 的3'-非翻译区691-697 存在结合位点。Pearson 相关分析显示,结直肠癌组织中miR-876-5p 表达与PAX6 mRNA 表达呈负相关(r=-0.754,P<0.05)。
2.3 结直肠癌组织中miR-876-5p、PAX6 mRNA 表达与临床病理参数的关系 结直肠癌组织中miR-876-5p、PAX6 mRNA 表达与肿瘤分化程度、TNM 分期、淋巴结转移有关(P均<0.05),与性别、年龄、肿瘤部位、肿瘤最大径无关(P均>0.05)。见表2。
表2 结直肠癌组织中miR-876-5p、PAX6 mRNA表达与患者临床病理参数的关系(±s)
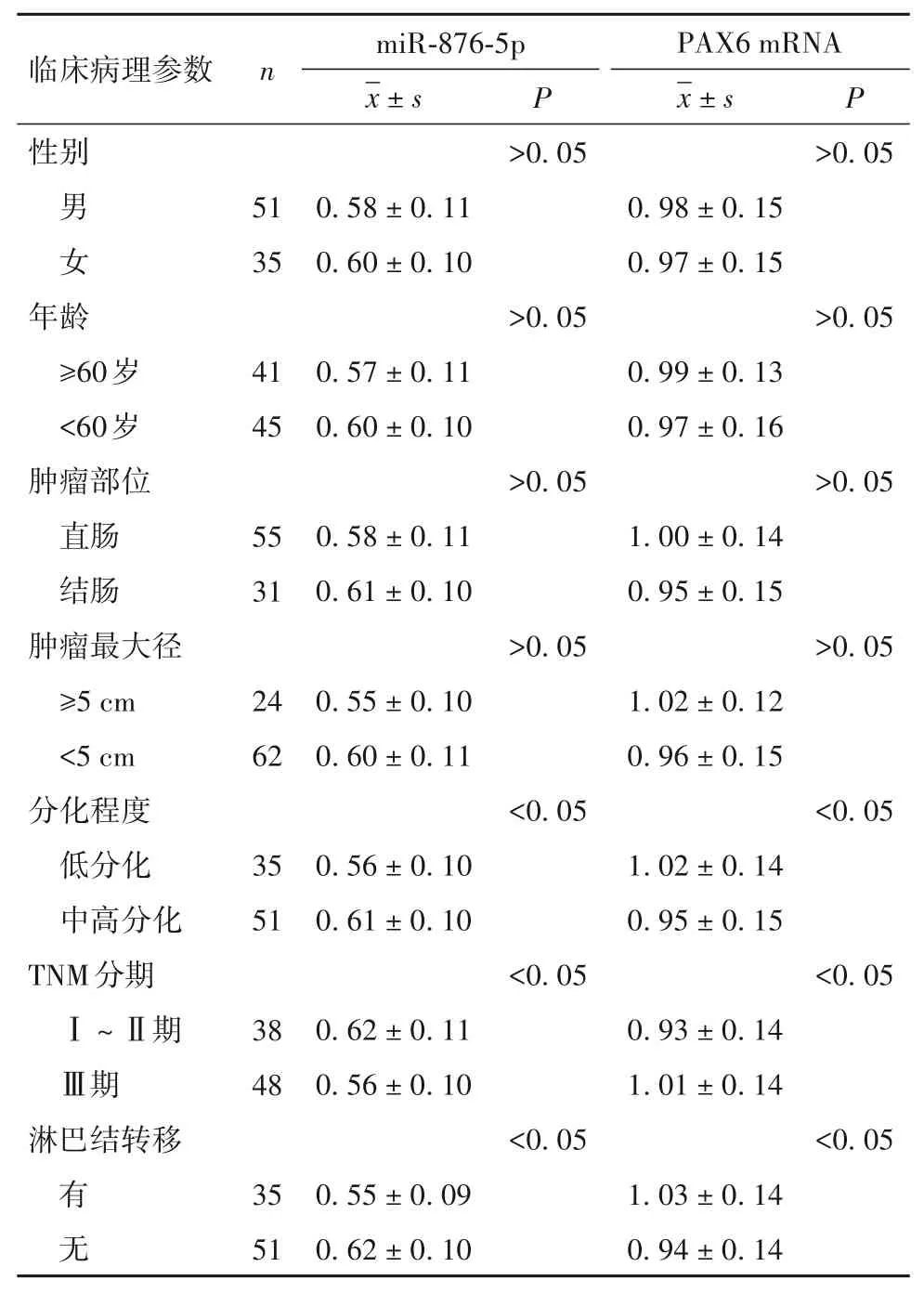
表2 结直肠癌组织中miR-876-5p、PAX6 mRNA表达与患者临床病理参数的关系(±s)
临床病理参数性别 男 女年龄 ≥60岁 <60岁肿瘤部位 直肠 结肠肿瘤最大径 ≥5 cm <5 cm分化程度 低分化 中高分化TNM分期 Ⅰ~Ⅱ期 Ⅲ期淋巴结转移 有 无n 51 35 41 45 55 31 24 62 35 51 38 48 35 51 miR-876-5pimages/BZ_13_674_338_698_384.png±s 0.58 ± 0.11 0.60 ± 0.10 0.57 ± 0.11 0.60 ± 0.10 0.58 ± 0.11 0.61 ± 0.10 0.55 ± 0.10 0.60 ± 0.11 0.56 ± 0.10 0.61 ± 0.10 0.62 ± 0.11 0.56 ± 0.10 0.55 ± 0.09 0.62 ± 0.10 P>0.05>0.05>0.05>0.05<0.05<0.05<0.05 PAX6 mRNAimages/BZ_13_674_338_698_384.png±s 0.98 ± 0.15 0.97 ± 0.15 0.99 ± 0.13 0.97 ± 0.16 1.00 ± 0.14 0.95 ± 0.15 1.02 ± 0.12 0.96 ± 0.15 1.02 ± 0.14 0.95 ± 0.15 0.93 ± 0.14 1.01 ± 0.14 1.03 ± 0.14 0.94 ± 0.14 P>0.05>0.05>0.05>0.05<0.05<0.05<0.05
2.4 结直肠癌组织中miR-876-5p、PAX6 mRNA 表达与预后的关系 86例结直肠癌患者随访6~36个月,失访6 例,死亡18 例,3 年累积生存率为79.07%(68/86)。miR-876-5p 高表达组3 年累积生存率90.48%(38/42)高 于miR-876-5p 低 表 达 组 的68.18%(30/44);PAX6 mRNA 高表达组3 年累积生存率70.21%(33/47)低于PAX6 mRNA 低表达组的89.74%(36/39),比较差异均有统计学意义(P均<0.05)。
3 讨论
研究表明,表观遗传学在结直肠癌中扮演重要角色[9]。miRNA 是表观遗传学的一个新的研究热点,能通过与mRNA 的3'-非翻译区互补或配对引发mRNA 降解或转录后沉默,进而调控结直肠癌发生发展[10-11]。miR-876-5p 定位于人9 号染色体短臂21.1 处,最初研究发现miR-876-5p 能通过Toll 样受体4/核因子-κB 信号通路参与炎症级联反应;近年研究发现miR-876-5p 定还参与肿瘤过程,如miR-876-5p 能靶向G 蛋白亚基γ12 激活磷脂酰肌醇 3-激酶/蛋白激酶B 信号通路抑制胶质瘤细胞增殖、迁移[12];同时能靶向G 蛋白偶联受体激酶5 抑制三阴性乳腺癌细胞增殖、迁移[13]。本研究结果显示,结直肠癌组织中miR-876-5p 表达降低,其机制可能是miR-876-5p 启动子被甲基化导致表达沉默有关[12-13]。本研究结果还显示,结直肠癌组织中miR-876-5p 表达与分化程度、TNM 分期、淋巴结转移有关,进一步说明miR-876-5p 低表达参与结直肠癌发生发展,其机制可能与miR-876-5p 能靶向RAS 蛋白激活样因子2(RASAL2)有关。RASAL2 是一种RAS-GTP 酶激活蛋白,能通过诱导结直肠癌上皮-间充质转化促进其发生发展[14]。REN 等[15]实验显示,miR-876-5p 能靶向RASAL2 抑制结直肠癌细胞增殖和侵袭。
结直肠癌是多因素参与的复杂过程,其过程中伴随多个基因的异常表达[16-17]。PAX6 是一种核转录因子,具备同源结构域和配对结构域两个DNA 结合域,能通过识别和结合特点的DNA 序列盒,激活或抑制钙黏蛋白、B 细胞淋巴瘤/白血病-2、细胞周期蛋白D1、p21、p27等下游靶基因调控细胞增殖、分化、侵袭、迁移和凋亡[18]。基于PAX6 对多种基因的调控作用,有大量研究报道了其与肿瘤的关系,如PAX6 能靶向抑制同源域转录因子1 增强胃癌细胞增殖和侵袭能力[19];能靶向上调N-钙黏蛋白、波形蛋白促进视网膜母细胞瘤增殖和侵袭[20]。本研究结果显示,结直肠癌组织中PAX6 mRNA 表达降低,其机制可能与PAX6 的启动子被结直肠癌中原癌基因激活转录而大量表达有关[19-20]。本研究结果还显示,结直肠癌组织中PAX6 mRNA 表达与分化程度、TNM 分期、淋巴结转移有关,进一步说明PAX6 mRNA 高表达参与结直肠癌发生发展,其机制可能与激活或抑制下游靶基因有关。如PAX6能上调N-钙黏蛋白表达和抑制E-钙黏蛋白表达,增强结直肠癌细胞活力,促进其增殖和转移[21]。通过TargetScan数据库预测发现,miR-876-5p 与PAX6 存在结合位点,相关性分析也发现结直肠癌组织中miR-876-5p与PAX6 mRNA 表达呈负相关,提示二者可能共同参与结直肠癌的发生、发展。李程等[22]研究也报道,上调miR-876-5p 表达能靶向下调PAX6 表达,抑制结肠癌细胞增殖、迁移、侵袭和促进其凋亡。进一步验证了本研究结果。为探究miR-876-5p、PAX6 与结直肠癌患者预后的关系,本研究通过随访发现,miR-876-5p 高表达和PAX6 mRNA 低表达的结直肠癌患者3 年累积生存率更高,提示miR-876-5p、PAX6 mRNA表达还与结直肠癌患者预后有关,有望成为结肠癌患者预后潜在指标。
综上所述,结直肠癌组织中miR-876-5p 高表达和PAX6 mRNA 低表达与病理参数和预后相关,二者可能共同参与结直肠癌进展。但本研究样本量较少且为临床研究,还需进一步大样本研究证实,并深入研究miR-876-5p、PAX6参与结直肠癌发生发展的机制。
