乌鳢globin基因家族的鉴定及在鳃和鳃上器官中的表达模式分析❋
2023-03-14陶泽鑫李建龙温海深孙冬磊陈基伟
陶泽鑫,李建龙,温海深,李 超,孙冬磊,陈基伟,李 昀❋❋
(1.海水养殖教育部重点实验室(中国海洋大学),山东 青岛 266003;2.威海海洋职业学院海洋生物系,山东 威海 264300;3.青岛农业大学海洋科学与工程学院, 山东 青岛 266200)
珠蛋白(Globin)是一类含有血红素的蛋白质,能够在卟啉环中心的铁原子和多肽链上的组氨酸残基之间可逆地结合氧气与其他气体[1]。在脊椎动物中,Globin进化成血红蛋白(Hemoglobin,Hb)、肌红蛋白(Myoglobin,Mb)、细胞珠蛋白(Cytoglobin,Cygb)、神经珠蛋白(Neuroglobin,Ngb)、雄激素结合珠蛋白(Androglobin,Adgb)、珠蛋白E(GbE)、珠蛋白Y(GbY)和珠蛋白X(GbX)8种不同的类型[2]。其中Adgb起源于动物进化早期[3],与其他globin基因家族成员关系遥远,不在本文研究范围内。Hb和Mb是目前研究较多的两类Globin。Hb存在于红细胞中,是由两个α亚基和两个β亚基组成的四聚体结构,每条链均能与一个亚铁血红素结合。Mb存在于脊椎动物的心肌和骨骼肌中,是一种只有三级结构的单链蛋白质,与Hb的α和β亚基三级结构极为相似。Hb和Mb不仅能够储存和运输氧气,还具有催化亚硝酸盐降解、调节一氧化氮(Nitric oxide,NO)的产生和消除以及活性氧(Reactive oxygen species,ROS)防御等作用[4-5]。此外,Mb还具有增加线粒体中可用氧气的功能[6]。近十几年,随着基因组研究的迅速发展,Ngb、Cygb和GbE等新的globin基因家族成员被鉴定出来,然而其具体的功能机制尚未明确。Ngb主要在神经细胞和内分泌细胞中表达,推测其具有氧化代谢、消除NO和ROS以及抑制细胞凋亡等多种功能[7]。Cygb主要在成纤维细胞以及神经细胞中表达,可能参与胶原蛋白合成[8-9]和ROS防御[10]过程。此外,在一些脊椎动物中还存在特殊的变异Globin亚型,如仅在鸟类、海龟和肺鱼等存在的GbE[11-12],仅在热带爪蟾(Xenopustropicalis)、非洲爪蟾(Xenopuslaevis)和鸭嘴兽(Ornithorhynchusanatinus)等部分有颌类脊椎动物中存在的GbY[13-14],只在硬骨鱼类和爪蟾中存在的GbX[15]。目前这几种Globin的具体生理功能仍处于研究阶段[16]。
乌鳢(Channaargus)隶属鲈形目(Perciformes)鳢科(Channidae)鳢属(Channa),是一种营养价值较高的淡水经济鱼类,富含蛋白质、必需氨基酸、二十二碳六烯酸(DHA)等多不饱和脂肪酸以及铁、锌和铜等多种微量元素[17-18]。乌鳢生活在含氧率较低的下层水环境,具有很强的低氧耐受能力。研究发现,乌鳢的红细胞数量非常高[19],且血红蛋白含量也显著高于其他鱼类[20]。同时乌鳢还具有一种特殊的辅助呼吸器官——鳃上器官(Suprabranchial organ),可用于在低氧环境中进行空气呼吸,甚至在离开水的情况下短暂生存[21]。然而,关于乌鳢耐低氧及空气呼吸的分子机制尚未被阐明。
目前,许多学者对于硬骨鱼类中globin基因的结构和功能进行了研究。有学者对黄河裸裂尻鱼(Schizopygopsispylzovi)中globin基因家族进行了鉴定,发现9个基因成员,并对其在低氧条件下的表达模式进行研究,结果表明在重度缺氧条件下hb、cygb和ngb等基因表达水平显著降低,但mb在心脏中表达水平显著升高[22]。蟾胡子鲶(Clariasbatrachus)是一种能在陆地上呼吸和移动的淡水鱼,具有能够进行空气呼吸的鳃上器官。比较基因组学和转录组学研究表明,蟾胡子鲶基因组中mb基因相较于其他硬骨鱼类出现扩增,且hb基因在辅助呼吸器官中表达水平显著高于鳃[23],推测其与空气呼吸机制有关。为探究Hb、Mb及其他Globin在同样具有耐低氧和空气呼吸能力的乌鳢中发挥的潜在作用,本文对乌鳢globin基因家族进行了全基因组鉴定、系统进化分析及基因结构分析,并对其在鳃上器官发育空气暴露胁迫中鳃和鳃上器官的表达模式进行了分析。研究结果为阐明Globin在乌鳢耐低氧和空气呼吸过程中的功能机制提供了理论基础。
1 材料与方法
1.1 乌鳢globin基因的鉴定和序列分析
从NCBI(http://www.ncbi.nlm.nih.gov/)数据库中获取人(Homosapiens)、斑马鱼(Daniorerio)和攀鲈(Anabastestudineus)的所有globin基因家族成员的氨基酸序列,通过TBLASTN(1×10-5)对本课题组前期获得的乌鳢基因组数据库(JAJQTP000000000)和转录组数据库(PRJNA832507)进行检索,初步确定乌鳢的globin基因候选核苷酸序列,并通过在线网站SoftBerry(http://linux1.softberry.com/berry.phtml)的基因结构预测功能进一步注释。然后,通过BLASTN(1×10-10)进行比对,明确基因序列和拷贝数,并对几种不同脊椎动物的globin基因拷贝数进行比较分析。通过NCBI在线工具Open Reading Frame(ORF)Finder(https://www.ncbi.nlm.nih.gov/orffinder/)预测乌鳢globin基因的氨基酸序列。利用ExPASy Prot-Param工具(https://web.expasy.org/protparam/)预测乌鳢Globin的分子量(molecular weight,MW)和等电点(isoelectric point,pI)。通过CDD数据库(https://www.ncbi.nlm.nih.gov/cdd/)对乌鳢Globin的特征结构域进行预测。利用在线网站MEME(https://meme-suite.org/meme/tools/meme)对乌鳢Globin的保守基序(motif)进行分析,并通过TBtools软件[24]对结构域和motif进行可视化。
1.2 globin基因系统进化分析和共线性分析
将预测获得的乌鳢globin基因的氨基酸序列与人、小鼠(Musmusculus)、鸡(Gallusgallus)、斑马鱼和攀鲈等脊椎动物的氨基酸序列进行系统进化分析。将所有氨基酸序列输入MEGA 7.0软件[25],用ClustalW进行多重序列比对(默认参数),通过Neighbor-Joining(NJ)法构建系统进化树,bootstrap值选择重复1 000次。利用在线网站Interactive Tree Of Life(http://itol.embl.de/)制作出进化树图。
通过共线性分析进一步确定基因的同源性。利用多重共线性扫描工具包(MCScanX)的默认参数[26]分析乌鳢基因组的共线性关系以及乌鳢基因与其他物种基因的共线性关系。通过Circos[27]展示乌鳢基因组中的共线性基因,并将globin基因定位到乌鳢染色体上。在NCBI数据库获取斑马鱼(GRCz11)和攀鲈(fAnaTes1.2)的最新版参考基因组数据,通过Multiple Synteny Plot工具(https://github.com/CJ-Chen/TBtools)进行物种间的共线性分析。
1.3 乌鳢鳃上器官发育时期的样品采集
乌鳢鳃上器官发育时期的实验用鱼取自山东省临沂市沂水县个体养殖户。在繁殖季节挑选体格健壮、性腺发育良好的乌鳢亲鱼,通过人工授精获得受精卵,在繁殖池进行孵化和培育。分别在乌鳢仔稚鱼受精后3、5和8 d时,将仔稚鱼用MS-222麻醉后在体视镜下用镊子采集鳃上器官样品,每个时间点设置3个生物学重复(每个样品采集24尾鱼进行混样),样品在RNA保护液中保存备用。
1.4 乌鳢空气暴露胁迫实验的样品采集
空气暴露胁迫的实验用鱼取自于江苏省连云港市东海县辰发黑鱼养殖专业合作社,随机挑选体质健康、规格均一的4月龄乌鳢幼鱼(体长为(23.69±1.36)cm,体质量为(113.27±14.05)g)暂养2周后进行实验。将乌鳢幼鱼放在底部放置了湿润海绵的100 L实验空桶中进行空气暴露胁迫实验,每个桶中放入20尾幼鱼。分别在暴露于空气中0、3、6和24 h时,将幼鱼用MS-222麻醉后采集鳃和鳃上器官样品,每个时间点设置3个生物学重复(每个样品采集3尾鱼进行混样),样品在液氮中速冻后,放置于-80 ℃冰箱中保存备用。
1.5 乌鳢globin基因的表达分析
采用TRIZOL法(Invitrogen,美国)对以上样品总RNA进行提取,并用NanoDrop 2000核酸定量仪(Thermo Scientific,美国)和琼脂糖凝胶电泳对总RNA进行质量检测。质检合格后,用TruSeq Stranded mRNA LTSample Prep Kit试剂盒(Illumina,美国)构建cDNA文库,并通过Agilent 2100生物分析仪(Agilent Technologies,美国)进行质量检测。质检合格后,用Illumina HiSeq X Ten平台进行高通量测序(High-Throughput Sequencing)。
测序得到原始数据(Raw data),然后通过Trimmomatic软件[28]进行质控过滤,得到干净数据(Clean data)。通过Hisat2软件[29]将Clean data比对到乌鳢参考基因组,利用Samtools软件[30]对比对结果文件排序去重,使用stringtie软件[31]计算基因表达水平,得到globin基因表达量的FPKM值,并通过DESeq2 R包(http://www.bioconductor.org/packages/release/bioc/html/DESeq2.html)进行差异表达统计分析,以P<0.05表示差异显著。取log10(FPKM+1),利用pheatmap R包(https://CRAN.R-project.org/package=pheatmap)展示globin基因的表达量热图。
2 实验结果
2.1 乌鳢globin基因的鉴定及拷贝数分析
在乌鳢中共鉴定获得18条globin基因序列,分别是hbaa、hba1、hbad、hbb1、hbb2.1、hbb2.2、hbb2.3、hbb2.4、hbba、hbbc、hbae1.1、hbae1.2、hbae1.3、hbae1.4、mb、cygb、cygb1和ngb(见表1)。各globin序列的ORF长度在372~687个核苷酸之间,编码含有123~228个氨基酸的蛋白,它们的预测分子量在13.96~26.05 kDa之间。此外,其等电点和登录号也在表1中列出。

表1 乌鳢globin基因家族成员的序列特征
对globin基因在人、小鼠、鸡、斑马鱼、斑点叉尾鮰(IetalurusPunetaus)及具有空气呼吸能力的攀鲈、黄鳝(Monopterusalbus)和乌鳢共8种脊椎动物中的拷贝数进行了总结(见表2)。结果显示,相对于人、鼠、鸡等高等脊椎动物,globin基因家族在硬骨鱼类中发生显著扩增。其中,乌鳢中鉴定出18个拷贝,与黄鳝、斑点叉尾鮰比较接近,而攀鲈中具有最多的拷贝数,共23个globin基因。在globin基因家族的各亚家族中,Hb亚家族的基因种类和拷贝数是最多的,在乌鳢和其他硬骨鱼类中有14~20个,可达人、鼠、鸡等高等脊椎动物的1.5~2倍。在硬骨鱼类的Hb亚家族中主要包括hba、hbb和hbae基因成员,其拷贝数分别有3~4个、5~8个和3~8个,而不存在hbd、hbe、hbg、hbm、hbq和hbz。Cybg、Ngb和Mb亚家族基因在各物种中都有分布,且仅有1~2个拷贝。除部分硬骨鱼类中存在GbX以及鸡中存在GbE外,大多数物种中GbX、GbE和GbY亚家族基因都是缺失的(见表2)。

表2 几种典型脊椎动物中globin基因的拷贝数
2.2 globin基因系统进化分析
选取人、鼠、鸡、斑马鱼、攀鲈以及乌鳢的全部globin基因的氨基酸序列构建系统进化树。由于gbx、gbe和gby在大部分物种中是缺失的,添加斑点雀鳝(Lepisosteusoculatus)、弓鳍鱼(Amiacalva)、斑胸草雀(Taeniopygiaguttata)、纳氏鲟(Acipensernaccarii)、热带爪蟾等物种的gbx、gbe和gby的氨基酸序列以保证进化树的完整性。如图1所示,根据系统进化关系globin基因共分为8组,分别为Hbα、Hbβ、Cygb、Mb、Ngb、GbY、GbE和GbX。各组中硬骨鱼类的同类基因聚为一枝,其他高等脊椎动物聚为另一枝。乌鳢的各globin基因与硬骨鱼类聚为一枝,且最先与亲缘关系最近的攀鲈聚在一起(见图1)。系统进化分析结果表明globin基因在进化上较为保守,该结果为乌鳢globin基因注释提供了依据。
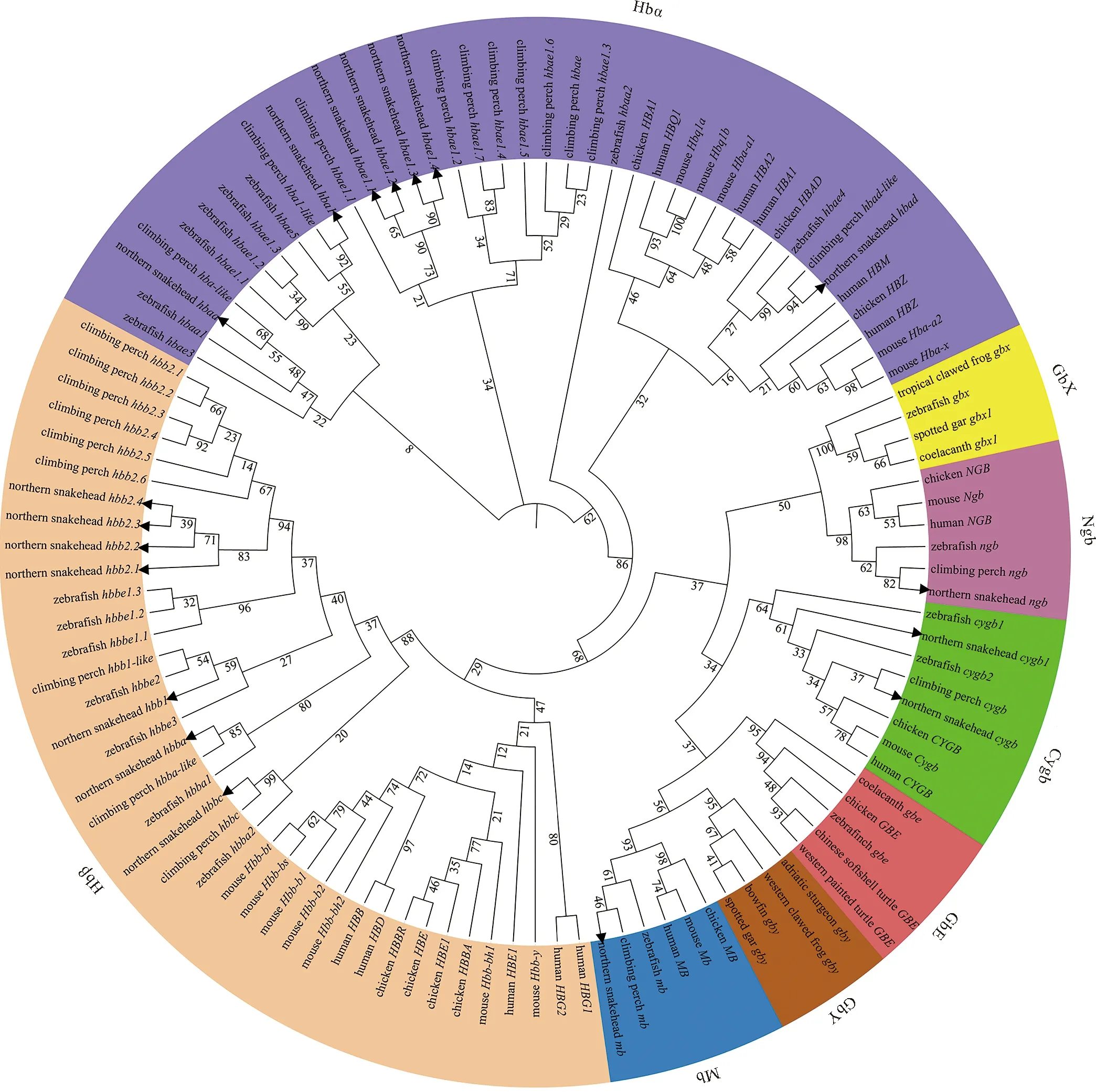
(分支节点上的数字表示bootstrapping值,黑色箭头表示乌鳢的globin基因,不同颜色圆环表示globin基因不同的亚家族。Numbers on node denote the bootstrapping values, black arrows denote globin genes of northern snakehead, and different colored rings denote different subfamilies of globin genes.)
2.3 乌鳢globin基因结构分析
图2展示了乌鳢globin基因的氨基酸序列的结构域和motif。利用CDD数据库预测得到4种类型的保守结构域,包括血红蛋白α链的Hb-α-like结构域,血红蛋白β链的Hb-β-like结构域,细胞珠蛋白的Cygb结构域和其他珠蛋白具有的Globin-like超家族结构域。此外,通过MEME数据库预测得到5个motif。Hbα链具有motif 1、2、3和4;Hbβ链具有motif 1、2、3、4和5,其中hbbc只有motif 1和3。其他亚家族差异较大,cygb和cygb1缺少motif 5,mb只含有motif 1和2,而ngb只含有motif 3和4(见图2)。

图2 乌鳢globin基因结构可视化分析
2.4 乌鳢globin基因共线性分析
通过Circos对乌鳢基因组的共线性区块、基因密度和globin基因在染色体上的位置进行展示。如图3所示,乌鳢的globin基因分布于Chr3、Chr17、Chr18和Chr20四条染色体上。Chr17上包含12个globin基因,是所有染色体中最多的,其中包括11个Hb亚家族基因和1个Cygb亚家族基因。Chr3上有1个Mb亚家族基因和1个Cygb亚家族基因,Chr18上有1个Ngb亚家族基因,Chr20上有3个Hb亚家族基因。此外还发现Chr17上的hbb2和hbae1均存在4个拷贝,在染色体上交替排列,形成串联重复事件。

(灰线表示乌鳢基因组的共线性区块,蓝色方块表示乌鳢染色体,外环表示基因密度。The gray line denotes the genome collinearity block of northern snakehead, the blue square denotes the chromosome of northern snakehead, and the outer ring denotes the gene density.)
对攀鲈、斑马鱼和乌鳢进行物种间共线性分析,共得到12个攀鲈和乌鳢globin基因有直系同源关系的共线性区块,以及10个斑马鱼和乌鳢globin基因的共线性区块,分别位于攀鲈的Chr1、Chr8、Chr12和Chr19,斑马鱼的Chr3、Chr12和Chr17,以及乌鳢的Chr3、Chr17、Chr18和Chr20染色体上(见图4)。

(灰线表示物种间的共线性区块,黑线表示乌鳢globin基因的共线性区块,不同颜色方块表示不同物种的染色体。The gray line denotes the collinearity block between species, the black line denotes the collinearity block of northern snakehead globin genes, and different colored ring squares denote the chromosomes of different species.)
2.5 乌鳢globin基因在鳃上器官发育和空气暴露胁迫中的表达分析
对乌鳢鳃上器官发育阶段的转录组数据、乌鳢空气暴露胁迫中鳃和鳃上器官的转录组数据进行整理,将globin基因的表达量用聚类热图进行展示(见图5)。
在仔稚鱼鳃上器官发育过程中,hbba和hbbc几乎检测不到表达(FPKM<1),其他基因表达水平差异较大,其中hba1、hbb1在鳃上器官不同发育时期均具有相对较高的表达水平(FPKM>1000)。hba1、hbb1、hbae1(包括hbae1.1、hbae1.2和hbae1.3)、hbb2(包括hbb2.1、hbb2.2和hbb2.4)和hbaa的表达量与其在鳃上器官发育早期阶段(受精后3 d)的表达量相比发生显著变化(P<0.05)。其中hbae1.1、hbb2.1和hbb2.2的表达量在发育中期阶段(受精后5 d)和发育后期阶段(受精后8 d)持续显著升高(受精后5 d的表达量分别为受精后3 d的5.14、4.36和3.87倍,受精后8 d的表达量分别为受精后3 d的26.03、23.83和23.63倍),而hbae1.2、hbae1.3、hbb2.4和hbaa的表达量在受精后8 d显著升高(表达量分别为受精后3 d的22.27、13.75、27.19和15.42倍)。此外,hba1和hbb1的表达量在受精后5 d显著降低(表达量分别为受精后3 d的0.42和0.41倍)。
在幼鱼的鳃和鳃上器官中,hbba相对其他globin基因具有较高的表达水平(FPKM>1 000),而hbae1.4、hbb2.4和ngb几乎检测不到表达(FPKM<1)。空气暴露胁迫后,hbae1(包括hbae1.1、hbae1.2和hbae1.3)、hbb2(包括hbb2.1和hbb2.2)、cygb和hbba的表达量发生显著变化(P<0.05)。其中在幼鱼鳃中,hbae1.1、hbb2.2、cygb和hbba的表达量在24 h显著升高(表达量为0 h的11.00、1.49、1.85和1.99倍),而hbae1.2的表达量在3 h显著降低(表达量为0 h的0.46倍)。在幼鱼鳃上器官中,hbae1.3和cygb的表达量在24 h显著升高(表达量为0 h的1.53和1.58倍),而hbb2.2和hbb2.1的表达量分别在3与6 h呈现显著降低的趋势(表达量为0 h的0.56和0.66倍)(见图5)。

(热图颜色表示表达量的高低,G表示鳃,S表示鳃上器官,表示显著差异(P<0.05)。Heatmap color denotes the level of expression, G denotes the gill, S denotes the suprabranchial organ, denotes ted the significant differences(P<0.05).)
3 讨论
Globin是一类小呼吸蛋白,能够通过血红素基团可逆地结合氧气,在脊椎动物中起到运输和储存氧气并进行氧化能量代谢的作用[32]。目前,globin基因家族已在高等脊椎动物中得到深入研究,但是在多数经济鱼类中尚未得到系统的研究。
鱼类的globin基因数量是脊椎动物中最多的,这有利于鱼类适应比陆生动物更为剧烈的环境和氧含量变化[33]。本研究中乌鳢的globin基因数量与其他硬骨鱼类相近,且相较于其他高等脊椎动物出现显著扩增。这种扩增现象可能是硬骨鱼类特有的第三次全基因组复制事件(WGD),即鱼类特异性基因组复制(FSGD)所导致的[34]。有学者认为,这种多重基因重复事件(Multiple gene duplication events)造成了globin基因家族复杂的进化过程[3]。串联复制是一种最常见的基因家族扩增机制[35]。在乌鳢的globin基因中,hbb2和hbae1基因均存在4个序列相似的拷贝,在Chr17上形成串联重复。同时,globin基因的系统进化分析和共线性分析表明,globin基因在进化上较为保守,乌鳢的globin基因与其他硬骨鱼类的globin基因具有较高的同源性。
与大多数脊椎动物相同,乌鳢的globin基因家族中具有Hb、Mb、Ngb和Cygb亚家族成员,而不存在GbE、GbY和GbX亚家族。通过对乌鳢globin基因的序列和结构进行分析,发现Hb亚家族成员在ORF长度、氨基酸数目、motif和结构域等方面保守性较高,而与其他亚家族成员的差异较大。表明在进化过程中不同亚家族间发生了功能分化,而亚家族内各成员功能较为保守。Hb是存在于红细胞中的呼吸蛋白,Hb起到结合氧气并提供能量的重要作用。有研究发现,鱼类可以通过Hb结构改变和基因拷贝数的增加来适应水中的低氧环境[36]。在乌鳢中,存在14个hb基因拷贝,包括hba、hbb、hbae、hbbc等不同类型,说明乌鳢的Hb结构与其他脊椎动物相似,是由2条Hbα链和2条Hbβ链组成的四聚体结构。此外,鉴定发现了一种血红蛋白β亚基阴极组件(Emoglobin cathodic subunit beta)基因(hbbc),该基因也存在于黄鳝、攀鲈和斑点雀鳝等其他可以进行空气呼吸的硬骨鱼类中。血红蛋白阴极组件与其他血红蛋白组分相比,具有更高的氧气结合能力[37]。据报道,翼甲鲶属(Pterygoplichthys)物种在缺氧的情况下,可以通过血红蛋白阴极组件所具有的较高氧气结合与运输能力,维持机体正常生命活动[38]。Mb和Ngb亚家族基因在大部分脊椎动物中只存在一个拷贝,具有较高的保守性。在具有空气呼吸能力的非洲肺鱼(Protopterusannectens)和蟾胡子鲶中,分别具有7和15个mb拷贝[23,39],这可能是它们适应低氧环境的关键策略。在金鱼(Carassiusauratus)中ngb的表达水平与耐低氧的能力呈正相关[40]。但在乌鳢中,Mb和Ngb均只有一个拷贝,且在鳃和鳃上器官中的表达水平无显著差异。此外据报道,由于第三次全基因组复制事件,硬骨鱼类具有2个cygb拷贝[41]。在乌鳢中也同样发现了2个cygb基因。
研究表明,在硬骨鱼类个体发育的不同阶段hb基因的表达会发生变化,这种现象称为血红蛋白的转换表达(Hemoglobin switching)[42]。根据仔稚鱼鳃上器官发育和幼鱼空气暴露胁迫转录组中globin基因表达的聚类热图,可以发现仔稚鱼和幼鱼中globin基因的转录表达模式也存在明显差异。对仔稚鱼鳃上器官发育中globin基因的表达分析表明,hba1、hbb1在不同发育时期的鳃上器官中均有较高的表达量,而hbae1、hbb2和hbaa等基因的表达量在发育中期和后期显著升高,可能在鳃上器官的生成过程中发挥着重要作用。hbae基因是硬骨鱼类globin基因家族中特有的一类调控胚胎期血红蛋白生成的基因,有研究对斑马鱼中hbae1.1进行敲除,发现缺失该基因会导致Hb含量明显下降[43]。在乌鳢中共发现4个hbae1基因的拷贝,其表达量在仔稚鱼中高于幼鱼,并在仔稚鱼的鳃上器官发育中期和后期呈逐渐升高的趋势,说明hbae1对鳃上器官中Hb的生成具有重要影响。此外,研究发现hbae1和hbb2基因成员在鳃上器官发育中具有相似的表达模式,推测是由于两者以头对头(3′-5′—5′-3′)的方式串联,共享启动子与增强子,存在协同表达机制[44]。对幼鱼鳃和鳃上器官的空气暴露胁迫中globin基因的表达分析表明,hbae1、hbb2、cygb和hbba等基因的表达量在暴露于空气24 h时显著升高,说明进行较长时间的空气呼吸可能导致乌鳢的鳃和鳃上器官中globin基因的表达水平升高。对各globin基因功能进行分析发现,cygb可能在鳃和鳃上器官的结缔组织形成中发挥着重要作用,而hbae1、hbb2和hbba等基因则可能与鳃和鳃上器官中Hb的生成有关。
4 结语
本研究对乌鳢的globin基因家族进行了鉴定,并对其进行了系统进化分析、基因结构分析和表达分析等相关研究。研究表明,乌鳢共有18个globin基因,分属于Hb、Mb、Ngb和Cygb四个亚家族,且在进化上较为保守。相较于高等脊椎动物,乌鳢的globin基因数量出现显著扩增。转录组测序数据分析结果表明,在仔稚鱼中,hbae1(包括hbae1.1、hbae1.2和hbae1.3)、hbb2(包括hbb2.1、hbb2.2和hbb2.4)和hbaa的表达量在鳃上器官发育过程中呈显著升高的趋势,而hba1和hbb1的表达量则在发育中期显著降低。在幼鱼中,hbae1(包括hbae1.1、hbae1.2和hbae1.3)、hbb2(包括hbb2.1和hbb2.2)、cygb和hbba的表达量在空气暴露胁迫后不同时间点的鳃和鳃上器官中发生显著变化。这些globin基因可能在乌鳢空气呼吸及低氧胁迫中发挥潜在的生物学功能。本研究为进一步探究globin基因在乌鳢耐低氧和空气呼吸中的功能机制提供了理论基础。
