微RNA-132-3p靶向第10号染色体缺失的磷酸酶和张力蛋白同源物调控葡萄膜黑色素瘤细胞增殖与凋亡
2023-03-14许志波
许志波
葡萄膜黑色素瘤是一种罕见的癌症, 也是最多见的一种成人原发性眼内肿瘤[1]。虽然通过手术摘除或放射治疗是控制眼内肿瘤的好方法, 但高达一半的病人会发展为远处转移性疾病, 致使病人预后很差[2-3]。对于转移性葡萄膜黑色素瘤尚无有效的治疗方法, 大多数病人转移后存活不到12个月[3]。目前, 葡萄膜黑色素瘤的发病机制仍不明确, 并且缺乏有效的生物标志物能够识别高风险病人。因此迫切需要筛选肿瘤特异性预后因子以预测病人预后。微RNA(miRNA/miR)是一类高度保守的小分子内源性非编码RNA, 日益成为肿瘤等各种疾病发生的重要调节因子[4], 也可作为葡萄膜黑色素瘤的潜在生物标志物[5]。有研究对不同病理类型的葡萄膜黑色素瘤组织和正常葡萄膜黑色素组织进行miRNA的差异分析, 发现microRNA-132-3p(miRNA-132-3p, miR-132-3p)在梭形和上皮细胞型葡萄膜黑色素瘤组织中表达上调[6], 然而miR-132-3p对葡萄膜黑色素瘤细胞生物行为的影响及其机制尚未见相关报道。本研究于2018年2月至2019年7月利用体外细胞实验, 观察miR-132-3p在葡萄膜黑色素瘤细胞增殖和凋亡中的作用, 并进一步探索其可能的分子机制。
1 材料与方法
1.1 材料正常葡萄膜上皮细胞系ARPE-19和葡萄膜黑色素瘤细胞系SP6.5、M23购自武汉大学医学实验中心, 胎牛血清、洛斯维·帕克纪念研究所-1640(RPMI-1640)培养基购自美国Gibco公司, Lipofectamine 2000试剂购自美国Invitrogen公司, 膜联蛋白V(Annexin V)-异硫氰酸荧光素(FITC)/碘化丙啶(PI)凋亡检测试剂盒购自江苏凯基生物公司, 甘油醛-3-磷酸脱氢酶(GAPDH)、细胞周期蛋白D1(cyclin D1)、第10号染色体上缺失的磷酸酶和张力蛋白同源物(PTEN)、B细胞淋巴瘤-2(Bcl-2)、周期素依赖激酶抑制剂p21(P21)、Bcl-2相关X蛋白(Bax)一抗购自美国Cellular Signaling Technology公司, 辣根过氧化物酶(HRP)标记的二抗购自上海碧云天生物技术研究所, miR-132-3p、miR-132-3p干扰质粒(anti-miR-132-3p)、PTEN过表达质粒(pcDNA3.1-PTEN)、荧光素酶报告载体购自上海吉玛生物有限公司。
1.2 细胞培养与转染正常葡萄膜上皮细胞系ARPE-19、葡萄膜黑色素瘤细胞系SP6.5、M23细胞中接种至含10%胎牛血清的RPMI-1640培养基内, 培养于饱和湿度、37 ℃、5%二氧化碳的培养箱。取对数生长期的SP6.5细胞, 按2×105个/mL接种至6孔板, 利用Lipofectamine 2000试剂, 将miR-132-3p、anti-miR-132-3p、pcDNA3.1-PTEN及其相应的阴性对照转染至SP6.5细胞, 转染完成48 h后进行后续研究。
1.3 定量聚合酶链反应(qPCR)检测miR-132-3p表达取对数生长期的ARPE-19、SP6.5、M23细胞, TRIzol法提取总RNA并将其逆转录为互补DNA(cDNA), 进行qPCR反应。miR-132-3p正向引物5'GCGCGCGTAACAGTCTACAGC-3', 反向引物5'-GTCGTATCCAGTGCAGGGTCC-3'。U6正向引物5'-CTCGCTTCGGCAGCACATATACT-3', 反向引物5'-CGCTTCACGAATTTGCGTGT-3'。以U6为内参, 根据2-ΔΔCt法计算miR-132-3p的相对表达量。
1.4 MTT法检测细胞增殖收集各组SP6.5细胞, 加胰酶消化并调整细胞密度为2×104个/毫升。将细胞悬液接种至96孔板(100微升/孔), 常规培养24 h、48 h、72 h后, 分别加入100 μL的MTT溶液孵育4 h, 加入200 μL的二甲基亚砜后置于摇床上孵育10 min。多功能酶标仪测定各孔490 nm波长处SP6.5细胞吸光度值, 吸光度值越大, 表明细胞活性越强。
1.5 流式细胞术检测细胞凋亡收集各组SP6.5细胞, 加入500 μL结合缓冲液悬浮细胞, 分别加入5 μL Annexin V-FITC染色液和5 μL PI染色液, 室温条件下避光反应15 min, 通过流式细胞仪检测细胞凋亡。
1.6 蛋白质印迹法检测PTEN、cyclin D1、P21、Bcl-2、Bax蛋白表达收集各组SP6.5细胞, 分别加入放射免疫沉淀法(RIPA)裂解液, 提取总蛋白, 将蛋白沸水浴变性后进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE), 转膜。5%脱脂奶粉常温封闭1 h, 加入一抗(稀释比1∶1 000)4 ℃孵育24 h, 加Tris-HCl-Tween缓冲盐溶液(TBST)漂洗3次, 加入二抗(稀释比1∶2 000)37 ℃孵育1 h, 再次用TBST进行充分漂洗后加化学发光液进行显影。以GAPDH为内参, 计算PTEN、cyclin D1、P21、Bcl-2、Bax蛋白相对表达量。
1.7 双萤光素酶报告实验利用TargetScan数据库预测miR-132-3p的靶基因, 发现miR-132-3p与PTEN的3'非编码区(3'UTR)存在结合位点。构建含miR-132-3p结合位点的PTEN 3'UTR野生型(WT- PTEN)及突变型(MUT- PTEN)报告基因载体, 将其分别与miR-132-3p、miR-132-3p阴性对照(miRNC)共转染至S65细胞, 48 h后根据双萤光素酶报告基因检测试剂盒操作步骤, 检测细胞双萤光素酶相对活性。
1.8 统计学方法采用SPSS 22.0软件进行统计。计量数据表示为xˉ ±s, 两组间比较用两独立样本t检验;多组间比较用单因素方差分析, 多组间两两比较比较用SNK-q检验。P<0.05为差异有统计学意义。
2 结果
2.1 miR-132-3p在细胞ARPE-19、SP6.5和M23中的表达qPCR检测结果显示, miR-132-3p在葡萄膜黑色素瘤细胞SP6.5、M23及正常葡萄膜上皮细胞ARPE-19中表达量为0.94±0.09、0.81±0.08、0.26±0.02(F=78.70,P<0.001);SP6.5、M23均高于ARPE-19(P<0.05)。选择差异最显著的SP6.5细胞开展后续研究。
2.2 抑制miR-132-3p对细胞SP6.5增殖、凋亡的影响在SP6.5细胞中转染anti-miR-132-3p, 与antimiR-NC组比较, miR-132-3p表达量大幅减少, 24 h、48 h、72 h的细胞活性明显降低, 细胞凋亡率显著增加, cyclin D1、Bcl-2蛋白表达量降低而P21、Bax水平提高, 上述差异有统计学意义(P<0.05)。见图1, 2;表1, 2。
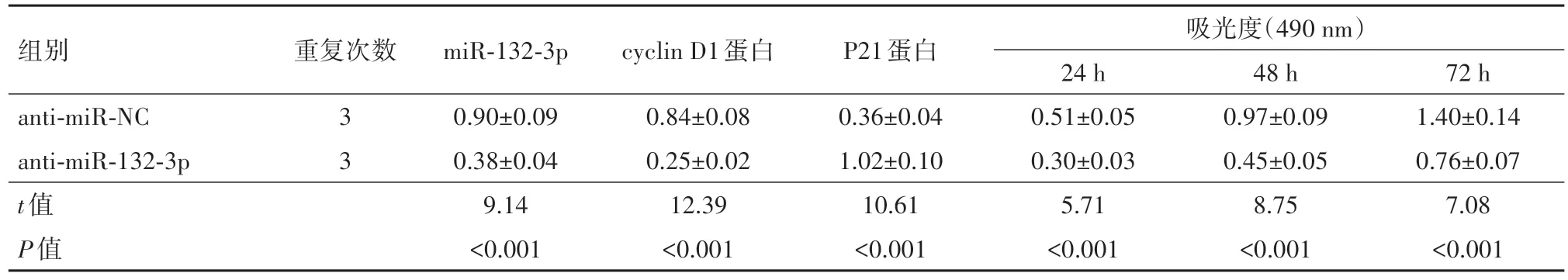
表1 抑制miR-132-3p对葡萄膜黑色素瘤细胞SP6.5增殖的影响/xˉ ±s
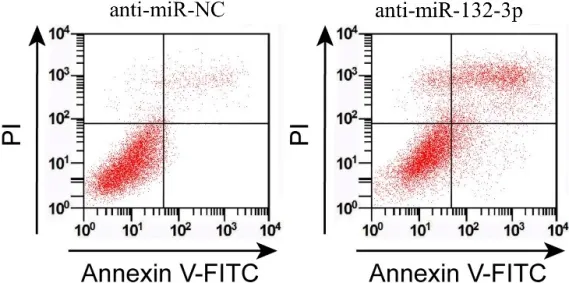
图1 抑制miR-132-3p对葡萄膜黑色素瘤细胞SP6.5凋亡的影响
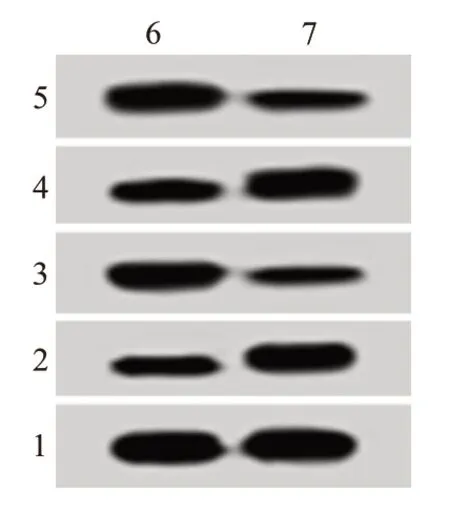
图2 抑制miR-132-3p对葡萄膜黑色素瘤细胞SP6.5增殖、凋亡蛋白表达的影响

表2 抑制miR-132-3p对葡萄膜黑色素瘤细胞SP6.5凋亡的影响/xˉ ±s
2.3 miR-132-3p靶向、调控PTEN生物学信息工具预测结果发现, PTEN的3'UTR部分碱基内含有miR-132-3p的结合位点, 见图3A。与miR-NC组比, miR-132-3p组WT- PTEN荧光素酶相对活性显著降低(1.02±0.10比0.21±0.02,t=13.76、P<0.05), MUT- PTEN荧光素酶相对活性差异无统计学意义(1.00±0.10比0.98±0.09,t=0.26、P>0.05)。与miR-NC组比, miR-132-3p组PTEN表达量显著降低(0.22±0.02比0.07±0.01,t=11.62、P<0.05), 与anti-miR-NC组比, anti-miR-132-3p组PTEN表达量显著升高(0.19±0.02比0.60±0.06,t=11.23、P<0.05), 见图3B。

图3 miR-132-3p靶向、调控PTEN:A为PTEN的3'非翻译区(3'UTR)含有miR-132-3p的互补序列;B为miR-132-3p调控PTEN的表达
2.4 过表达PTEN对细胞S65增殖、凋亡的影响在S65细胞中转染pcDNA3.1-PTEN, 结果发现, 相比于pcDNA3.1-PTEN阴性对照(pcDNA3.1)组, PTEN蛋白表达量明显升高, 24 h、48 h、72 h的细胞活性逐渐降低, 细胞凋亡率有所增加, Bcl-2、cyclin D1水平显著降低而P21、Bax表达量明显提高, 上述差异有统计学意义(P<0.05)。见表3, 4。
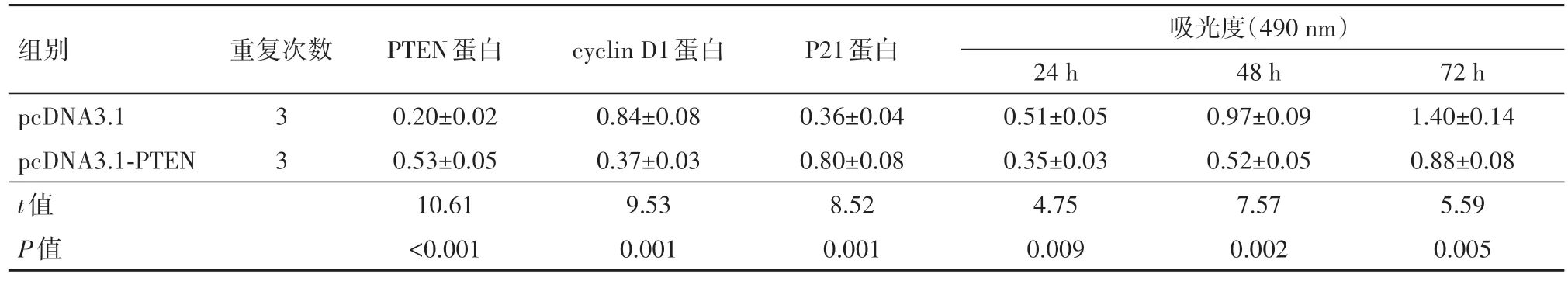
表3 过表达PTEN对葡萄膜黑色素瘤细胞SP6.5增殖的影响/xˉ ±s
2.5 抑制PTEN可逆转抑制miR-132-3p对细胞SP6.5增殖、凋亡的影响与anti-miR-NC组比较, 抑制miR-132-3p显著影响SP6.5细胞PTEN蛋白表达量、24 h、48 h、72 h的细胞活性、细胞凋亡率和cyclin D1、P21、Bcl-2、Bax蛋白水平(P<0.05), 结果同“2.2”“2.3”。与anti-miR-132-3p+si-NC组比较, antimiR-132-3p和si-PTEN共转染入SP6.5细胞, 24 h、48 h、72 h的细胞活性和cyclin D1、Bcl-2蛋白表达水平提高, 细胞凋亡率和PTEN、P21、Bax蛋白表达水平降低(P<0.05)。见表5, 6。

表5 抑制PTEN可逆转抑制miR-132-3p对细胞SP6.5增殖的影响/xˉ ±s
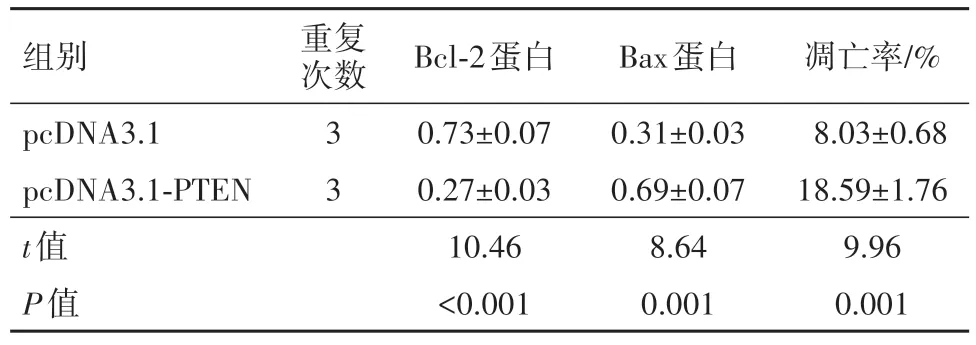
表4 过表达PTEN对细胞SP6.5凋亡的影响/xˉ ±s

表6 抑制PTEN能逆转抑制miR-132-3p对细胞SP6.5凋亡的影响/xˉ ±s
3 讨论
miRNA是一种短链非编码RNA, 长度从18到25个核苷酸不等, 在葡萄膜黑色素瘤等肿瘤进程中发挥抑癌或致癌作用[7], 如miR-21[8], miR-9[9], miR-181[10]。既往研究表明, miR-132-3p参与结直肠癌[11]、血管平滑肌脂肪瘤[12]、胃癌[13]等肿瘤发生发展过程。miR-132-3p在神经胶质瘤内皮细胞中的表达明显上调[14], miR-132-3p在结直肠癌细胞中的表达下调, 上调miR-132-3p的表达可显著抑制肿瘤细胞增殖、转移和迁移并促进细胞凋亡[15]。miR-132-3p过表达显著抑制骨肉瘤MG-63细胞增殖, 并使细胞凋亡率明显增加, 在骨肉瘤中起肿瘤抑制因子作用[16]。本研究中, miR-132-3p在SP6.5、M23细胞中表达上调, 与前人研究[7, 15]结果一致。同时, 抑制miR-132-3p表达显著降低24 h、48 h、72 h的SP6.5细胞活性及cyclin D1、Bcl-2蛋白表达水平(P<0.05), 提高细胞凋亡率和P21、Bax蛋白表达水平(P<0.05), 表明抑制miR-132-3p表达可以抗葡萄膜黑色素瘤细胞增殖并诱导其凋亡, 具有抗葡萄膜黑色素瘤作用。
PTEN是一种著名的抗肿瘤基因[17], 在葡萄膜黑色素瘤中具有抑癌作用[18]。Abdel-Rahman等[19]第一次证明PTEN是一种肿瘤抑制因子, 参与葡萄膜黑色素瘤的发病机制, 可能与临床结果有关。PTEN载体的过度表达抑制葡萄膜黑色素瘤MUM-2B细胞的侵袭[20]。在恶性黑色素瘤中, PTEN表达量明显降低, 其缺失与恶性黑色素瘤进展密切相关[21]。本研究中, 过表达PTEN明显抑制24 h、48 h、72 h的SP6.5细胞活性和Bcl-2、cyclin D1水平(P<0.05), 提升细胞凋亡率和P21、Bax表达量(P<0.05), 与抑制miR-132-3p表达结果相同。PTEN抑制葡萄膜黑色素瘤的作用与前述报道相同, 为PTEN作为肿瘤抑制因子增加了新的证据。
大量资料显示, miRNA通过与靶mRNA的3'UTR互补位点碱基配对, 导致mRNA降解、裂解或翻译抑制, 从而调控基因表达[22]。研究表明, 在乳腺癌细胞中, PTEN是miR-132和miR-212的直接靶点, miR-132或miR-212处理显著降低乳腺癌细胞MCF-7中PTEN表达[23]。在阿尔茨海默病中, miR-132关键靶点之一的PTEN表达上调[24]。在胶质瘤内皮细胞中, PTEN被鉴定为miR-132-3p的直接和功能性下游靶标[15]。本研究的生物信息学和双萤光素酶报告实验也证实了miR-132-3p对PTEN的靶向结合关系。另外, 抑制PTEN能逆转抑制miR-132-3p对SP6.5细胞抗增殖、促凋亡的作用, 表明miR-132-3p可能是通过调控PTEN表达影响葡萄膜黑色素瘤细胞增殖和凋亡的。
综上所述, miR-132-3p在葡萄膜黑色素瘤细胞中表达上调, miR-132-3p可通过靶向调控PTEN的表达抑制葡萄膜黑色素瘤细胞增殖并促进细胞凋亡, 这为葡萄膜黑色素瘤提供了新的候选预后生物标志物和治疗靶标。
