Sirt6通过调控P53/SLC7A11/GPX4通路抑制骨骼肌细胞铁死亡*
2023-03-10王瑶陈士坤段晨阳梁筱晗周成富秦珺侯东尧杜权
王瑶, 陈士坤, 段晨阳, 梁筱晗, 周成富, 秦珺, 侯东尧, 杜权,2△
Sirt6通过调控P53/SLC7A11/GPX4通路抑制骨骼肌细胞铁死亡*
王瑶1, 陈士坤1, 段晨阳1, 梁筱晗1, 周成富1, 秦珺1, 侯东尧1, 杜权1,2△
(1重庆医科大学附属第二医院麻醉科,重庆 400010;2重庆市老年医学临床研究中心,重庆 400010)
探讨骨骼肌细胞铁死亡的发生及sirtuin 6 (Sirt6)对其调控的分子机制。将C2C12小鼠成肌细胞分为对照组、erastin(Era;铁死亡诱导剂)组、Era+ferrostatin-1(铁死亡拮抗剂)组、Era+MDL-800(Sirt6激动剂)组和Era+OSS-128167(Sirt6抑制剂)组;通过细胞活力和C2C12成肌细胞肌源性分化情况观察铁死亡诱导剂及拮抗剂的影响;RT-qPCR和Western blot测定成肌细胞及肌管中Sirt6、肌萎缩标志物、铁死亡标志物的mRNA及蛋白表达水平;进一步检测P53蛋白乙酰化水平、胞内亚铁离子(Fe2+)、活性氧(ROS)、谷胱甘肽(GSH)及脂质过氧化指标Liperfluo;免疫荧光法检测肌管分化标志物肌球蛋白重链(MHC)荧光信号。Era降低成肌细胞活力和肌管分化质量,伴有Sirt6的mRNA和蛋白水平下降(<0.05),肌萎缩标志物肌肉环指蛋白1(MuRF1)和肌萎缩F盒蛋白(MAFbx)表达增加(<0.05)。激活Sirt6可抑制P53蛋白第381位赖氨酸乙酰化,降低P53表达水平,提高溶质载体家族7成员11(SLC7A11)和谷胱甘肽过氧化物酶4(GPX4)水平,改善ROS、GSH、Fe2+和脂质过氧化等铁死亡特异性指标(<0.05),逆转铁死亡导致的肌肉负性变化。反之,抑制Sirt6将进一步加重铁死亡。Sirt6可抑制P53蛋白乙酰化使其活性降低,通过调控P53/SLC7A11/GPX4信号通路抑制成肌细胞铁死亡,进而改善肌肉质量。
肌肉减少症;sirtuin 6;铁死亡;去乙酰化
随着老龄化社会的到来,年龄相关的进行性骨骼肌变化引发的一系列负面影响越发突出。肌肉减少症(sarcopenia;简称肌少症)作为一种以骨骼肌质量降低、力量下降、功能退化为特征的全身性肌肉综合征,是衰老的特征性表现[1-2]。患病老年群体自主活动能力下降、术后并发症增加及住院时间延长[3],给个人、家庭及社会带来巨大的心理压力和医疗经济负担。肌少症虽然已经被WHO定义为一种疾病,但对它的认识还远不清楚,值得我们进一步探索。
氧化应激、炎症反应、肌蛋白合成与代谢失调等均被认为是肌少症发生的原因[4-5]。骨骼肌是机体质量最大的组织,通过有氧代谢维持生命活动,其铁元素含量丰富并随年龄增长不断累积[6-8]。这一特性让铁死亡(ferroptosis)这种铁依赖的、胞内脂质过氧化物和活性氧簇(reactive oxygen species, ROS)急剧增加为特征的全新细胞死亡方式也参与到肌少症发生中[9-11]。铁死亡涉及多种基因信号表达变化,肿瘤抑制因子P53不仅参与肿瘤调控,近来也被发现在铁死亡诱导剂erastin (Era)作用后表达增强[12],抑制下游胱氨酸/谷氨酸逆向转运系统(System XC-)中的轻链亚基溶质载体家族7成员11(solute carrier family 7 member 11, SLC7A11)表达和胱氨酸摄取,导致谷胱甘肽(glutathione, GSH)依赖的谷胱甘肽过氧化物酶4(glutathione peroxidase 4, GPX4)活性降低,促进细胞膜上高表达的多不饱和脂肪酸发生脂质过氧化,从而发生细胞铁死亡[13-14]。
sirtuins是烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide, NAD+)依赖性脱乙酰酶,共有7个亚型。sirtuin 6 (Sirt6)是位于细胞核中的多效赖氨酸脱乙酰酶,在能量代谢、衰老、炎症、应激反应和癌症等多方面发挥重要作用[15-17]。近期有研究发现Sirt6能够调控胃癌[18]和胰腺癌[19]中的铁死亡,但在肌少症领域中鲜有报道。使用生物信息学分析发现P53蛋白是肌少症和铁死亡交叉的关键信号之一。P53表达受多种翻译后修饰影响,在其C末端区域中多个赖氨酸位点受乙酰化作用调控并提高P53稳定性[20]。Sirt6通过使P53的C末端区域关键乙酰化位点——第381位赖氨酸(Lys381)去乙酰化,降低P53活性,影响下游信号转导[21]。因此,本研究假设Sirt6通过去乙酰化作用降低P53表达,经P53/SLC7A11/GPX4信号通路抑制肌细胞铁死亡而改善骨骼肌质量,并进行实验验证。
材料与方法
1 实验材料及主要试剂
C2C12小鼠成肌细胞系购自中国科学院上海细胞库。C2C12专用培养液购自Procell;DMEM basic培养液和马血清均购自Gibco;铁死亡诱导剂Era、铁死亡特异性拮抗剂铁抑素1(ferrostatin-1, Fer-1)和Sirt6抑制剂OSS-128167 (OSS)均购自MedChemExpress;Sirt6激动剂MDL-800 (MDL)购自APExBIO;CCK-8试剂及抗Sirt6、肌肉环指蛋白1(muscle ring-finger protein 1, MuRF1)、SLC7A11、GPX4和GAPDH抗体均购自ABclonal;抗肌球蛋白重链(myosin heavy chain, MHC)抗体购自Santa Cruz;抗乙酰化P53 (Lys381)蛋白[Ac-P53 (Lys381)]抗体购自Immumoway;抗肌萎缩F盒蛋白(muscle atrophy F-box protein, MAFbx)抗体购自Proteintech;快速RNA提取试剂盒、Evo M-MLV反转录预混型试剂盒及SYBR Green PCR试剂盒均购自湖南艾科瑞生物工程有限公司;抗P53抗体购自Cell Signaling Technology;DCFH-DA荧光探针购自Biosharp;GSH测定试剂盒购自上海碧云天公司;胞内亚铁离子荧光探针FerroOrange及脂质过氧化物荧光探针Liperfluo均购自Dojindo。
2 细胞培养及分组
2.1细胞培养与肌管分化C2C12小鼠成肌细胞在37 ℃、5% CO2的培养箱中使用C2C12专用培养液进行培养,细胞密度达70%~80%进行传代或分化。分化培养液由DMEM basic培养液和2%马血清组成,每天更换分化培养液,分化5 d形成肌管用于实验。
2.2细胞、肌管治疗及分组将Era、Fer-1、OSS和MDL溶于二甲亚砜(dimethyl sulfoxide, DMSO)中,DMSO作为对照(control, Con)组。细胞和肌管用药物处理24或48 h,Fer-1、OSS或MDL提前预处理2 h。实验分组:Con组、Era (1 μmol/L)组、Era (1 μmol/L)+Fer-1 (1 μmol/L)组、Era (1 μmol/L)+OSS (100 μmol/L)组和Era (1 μmol/L)+MDL (5 μmol/L)组。
3 主要方法
3.1CCK-8实验按每孔5 000个细胞种板,细胞密度达80%加药反应不同时间。换液后每孔加入CCK-8试剂10 μL,反应0.5~4 h,使用多功能酶标仪测量450 nm波长下吸光度,用以反映细胞活力。
3.2肌源性分化形态学观察6孔板中细胞生长密度达80%,更换含不同药物的分化培养液,每天换液,共分化5 d。使用倒置荧光显微镜对肌管形态和数量进行可视化观察。
3.3肌管免疫荧光染色成肌细胞用含有不同药物的分化培养液处理5 d形成肌管。PBS溶液洗涤,冰甲醇固定20 min,0.5% Triton X-100冰上透膜15 min。5% BSA室温封闭1 h。MHC抗体(1:50)4 ℃孵育过夜,荧光二抗室温孵育1 h,滴加含DAPI抗荧光淬灭剂封片。使用正置荧光显微镜观察拍照,并用ImageJ软件分析。
3.4RT-qPCR实验使用快速RNA提取试剂盒从细胞或肌管中提取总RNA并测定浓度。再用反转录试剂盒去除gDNA,将RNA逆转录生成cDNA。最后加入SYBR Green PCR试剂,扩增反应程序为:95 ℃ 10 s;95 ℃ 5 s,60 ℃ 30 s,39个循环;95 ℃ 5 s,65 ℃ 5 s,95 ℃ 50 s,1个循环。采用2-ΔΔCt法分析数据。引物序列见表1。

表1 RT-qPCR引物序列
3.5蛋白印迹分析细胞或肌管药物处理48 h后,PBS洗涤,按100∶1∶1的比例加入细胞裂解液、蛋白酶抑制剂和去乙酰酶抑制剂混合物,提取总蛋白。BCA法定量蛋白浓度。配制12%聚丙烯酰胺凝胶电泳,恒压100 V湿转1 h,5%脱脂牛奶室温封闭2 h;加入抗Sirt6、P53、MAFbx、MuRF1、SLC7A11和GPX4抗体(1∶1 000),抗Ac-P53 (Lys381)抗体(1∶1 500),抗GAPDH抗体(1∶2 000),4 ℃孵育过夜;加入Ⅱ抗(1∶10 000),室温孵育1 h。使用增强型化学发光剂显影记录分析。
3.6ROS荧光强度测定96孔板中细胞密度达80%左右或肌管分化5 d,药物作用24 h。按1∶1 000稀释DCFH-DA荧光探针,在37 ℃、5% CO2培养箱中避光孵育30 min,PBS洗涤后使用酶标仪检测荧光信号(激发光波长:488 nm,发射光波长:524 nm)。
3.7GSH测定按说明书配制所需试剂,超声处理收集的细胞上清液,去除内源性GSH;混匀反应试剂,室温反应5 min;再加入50 μL NADPH溶液(0.5 g/L),室温反应25 min,用酶标仪检测。单点法绘制标准曲线并计算出GSH含量。
3.8胞内Fe2+测定96孔板培育细胞或肌管,密度达标后加药处理24h。去除培养液用PBS洗涤后,每孔中加入1 μmol/L FerroOrange工作液100 μL,在37 ℃、5% CO2培养箱中避光孵育30 min。吸尽工作液后使用多功能酶标仪(激发光波长:543 nm,发射光波长:580 nm)检测样品荧光强度,通过相对荧光强度反映胞内Fe2+变化。
3.9胞内脂质过氧化测定96孔板中药物处理细胞或肌管24 h。无血清培养液洗涤后,每孔中加入5 μmol/L Liperfluo工作液100 μL,在37 ℃、5% CO2培养箱中避光孵育30 min。PBS洗涤2次,使用多功能酶标仪(激发光波长:488 nm,发射光波长:545 nm)测定荧光强度。
4 统计学处理
利用GraphPad Prism 9.2软件进行分析作图,数值以均数±标准差(mean±SD)表示,≥3。两组间均数比较采用检验;多组间均数比较使用单因素方差分析,用Tukey法进行多组间两两比较分析。<0.05被认为具有统计学意义。
结果
1 铁死亡诱导剂降低成肌细胞活力及肌源性分化能力
为明确铁死亡是否影响成肌细胞活力,并确定后续实验药物作用浓度,采用不同浓度(0~5 μmol/L)的Era处理C2C12成肌细胞不同时间(12、24和48 h)。结果发现,随Era浓度的增加和作用时间的延长,细胞活力显著下降;不同浓度作用12 h细胞相对活力均大于50%;当药物浓度大于1 μmol/L时,作用24和48 h细胞活力过低,因此选择1 μmol/L作用48 h进行后续实验(图1A)。而Era引起的细胞活力降低能被铁死亡拮抗剂Fer-1逆转(图1B)。
在铁死亡对C2C12成肌细胞肌源性分化影响的研究中,细胞生长密度达到80%时更换分化培养液、含Era的分化培养液或含Era+Fer-1的分化培养液作用5 d,每天换液。显微镜下可见,与Con组相比,Era处理后C2C12成肌细胞数量和分化能力下降,胞核减少、肌管变短、形态异常、排列紊乱,但添加Fer-1后能部分恢复肌源性分化能力(图1C)。
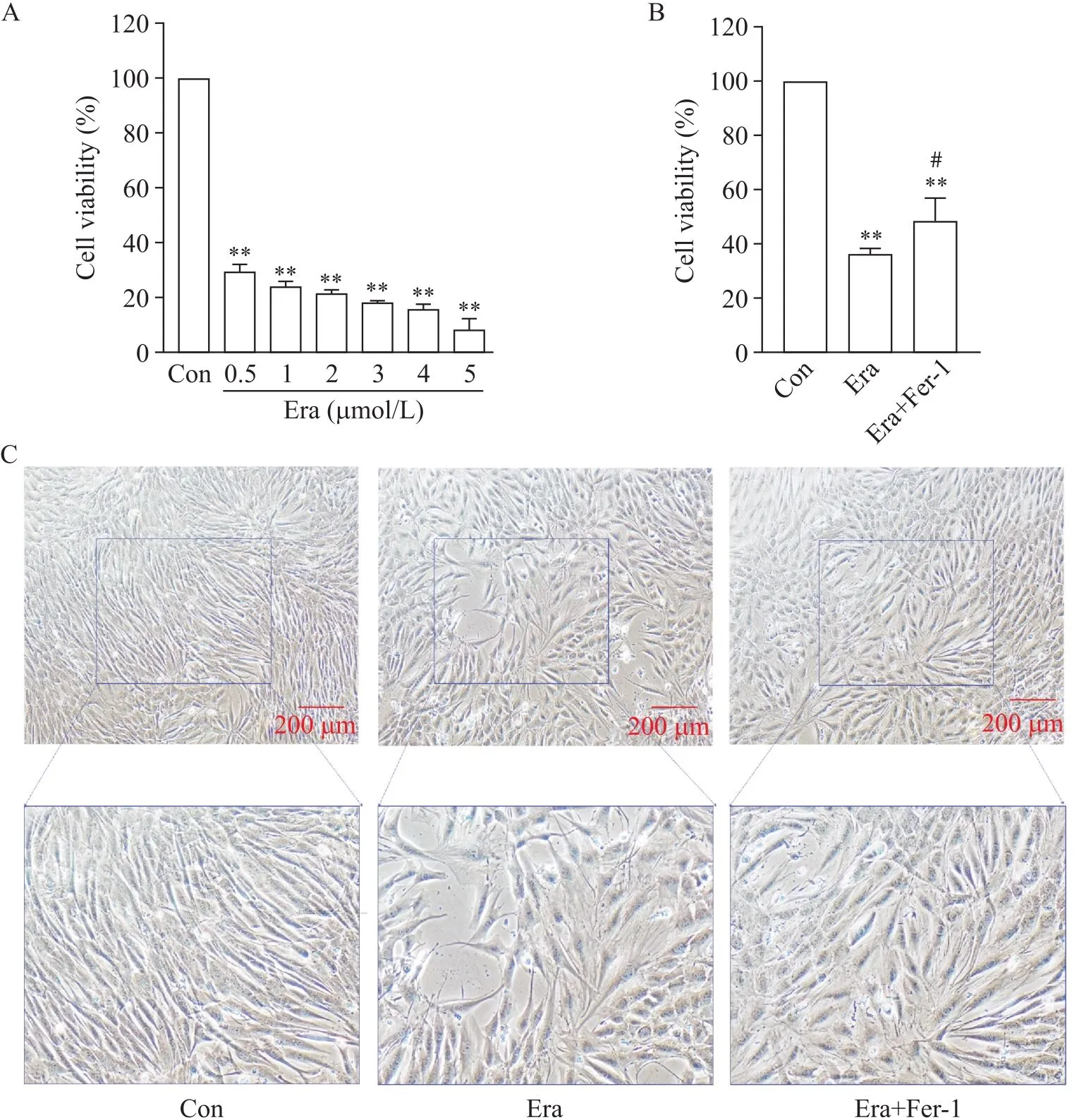
Figure 1. Effects of erastin (Era) on C2C12 myoblasts and myotubes. A: the viability of C2C12 myoblasts treated with different concentrations of Era for 48 h; B: the viability of C2C12 myoblasts at 48 h in control (Con), Era and Era+ferrostatin-1 (Fer-1) groups; C: the morphological changes of myoblasts differentiating into myotubes in different groups. Mean±SD. n=3. **P<0.01 vs Con group; #P<0.05 vs Era group.
2 Era作用的成肌细胞和肌管中Sirt6表达下降
加入Era作用C2C12成肌细胞48 h,RT-qPCR显示P53的mRNA表达水平升高,Sitr6及P53下游铁死亡相关分子SLC7A11和GPX4的mRNA表达水平却随之降低(图2A),且Sirt6会随药物作用时间延长(48 h和72 h比较)下降得愈发明显(图2A、B)。在蛋白层面也发现相同的变化(图2C)。

Figure 2. The mRNA and protein levels of Sirt6, P53, SLC7A11 and GPX4 in C2C12 myoblasts treated with different concentrations of erastin (Era). A: the mRNA levels of Sirt6, P53, SLC7A11 and GPX4 at 48 h; B: the mRNA level of Sirt6 at 72 h; C: the protein levels of Sirt6, P53, SLC7A11 and GPX4 at 48 h. Mean±SD. n=3. *P<0.05, **P<0.01 vs control (Con) or 0 μmol/L group.
在肌管中,Era会导致肌萎缩相关分子MAFbx和MuRF1的mRNA和蛋白水平升高,同时P53的蛋白水平升高,而Sirt6的mRNA和蛋白水平及铁死亡相关分子SLC7A11和GPX4蛋白水平下降(图3)。
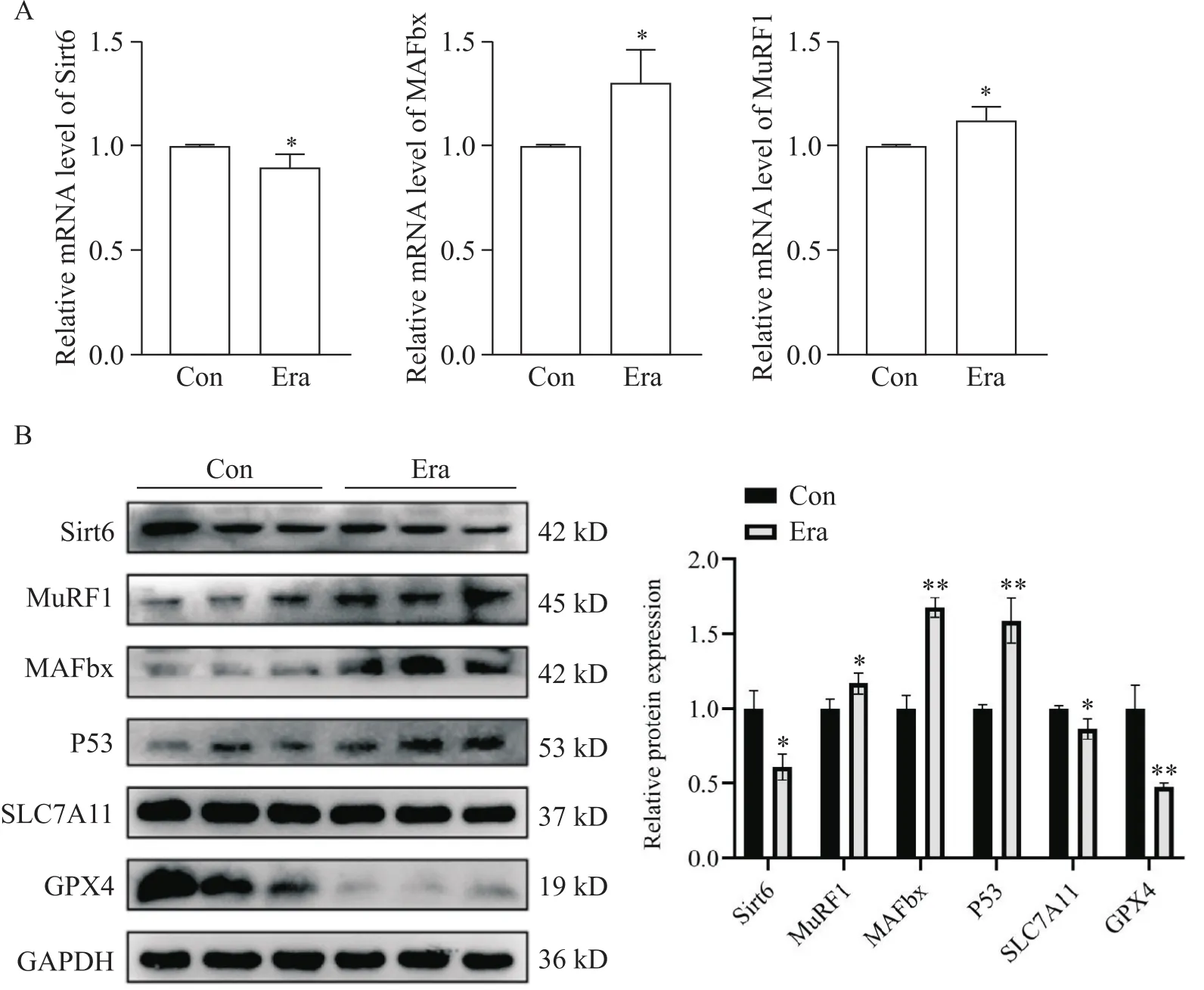
Figure 3. Myotrophic specific E3 ubiquitin ligases were up-regulated and Sirt6 declined after erastin (Era) treatment in C2C12 myotubes. A: the mRNA expression of Sirt6, MAFbx and MuRF1 in myotubes; B: the protein expression of MAFbx, MuRF1, P53, SLC7A11, GPX4 and Sirt6 in myotubes. Mean±SD. n=3. *P<0.05, **P<0.01 vs control (Con) group.
以上结果表明在铁死亡参与的肌肉病理改变中存在Sirt6的降低。Era作用使P53表达增加,可能通过影响P53/SLC7A11/GPX4铁死亡通路的信号变化,参与肌细胞和肌管铁死亡发生。
3 激活或抑制Sirt6对Era作用下成肌细胞活力、细胞肌源性分化和肌管的影响
3.1激活Sirt6能抑制Era引起的成肌细胞活力下降和肌管变化为探索Sirt6在铁死亡引起的肌质量下降中是否发挥作用,在Era处理的基础上预先加入Sirt6特异性激动剂MDL或抑制剂OSS处理成肌细胞和肌管。在成肌细胞中,MDL激动Sirt6会部分逆转Era引起的细胞活力降低(图4A),使P53的mRNA表达水平下降,SLC7A11和GPX4的mRNA表达水平升高(图4B),改善脂质过氧化等铁死亡指标(图5),与Fer-1作用类似;OSS处理后,伴随Sirt6表达下降,P53的mRNA表达水平升高,而SLC7A11的mRNA表达水平降低,肌细胞铁死亡指标反映铁死亡加重(图4B及图5)。以上结果进一步证实在Era诱导的铁死亡细胞模型中,P53可抑制SLC7A11表达并出现GPX4下降,Sirt6通过P53/SLC7A11/GPX4通路抑制肌细胞铁死亡。

Figure 4. The cell viability (A) and the mRNA levels of ferroptosis markers (B) after adding ferrostatin-1 (Fer-1), OSS-128167 (OSS) or MDL-800 (MDL) based on erastin (Era) treatment in C2C12 myoblasts. Mean±SD. n=3. **P<0.01 vs control (Con) group; #P<0.05, ##P<0.01 vs Era group; &&P<0.01 vs Era+OSS group.
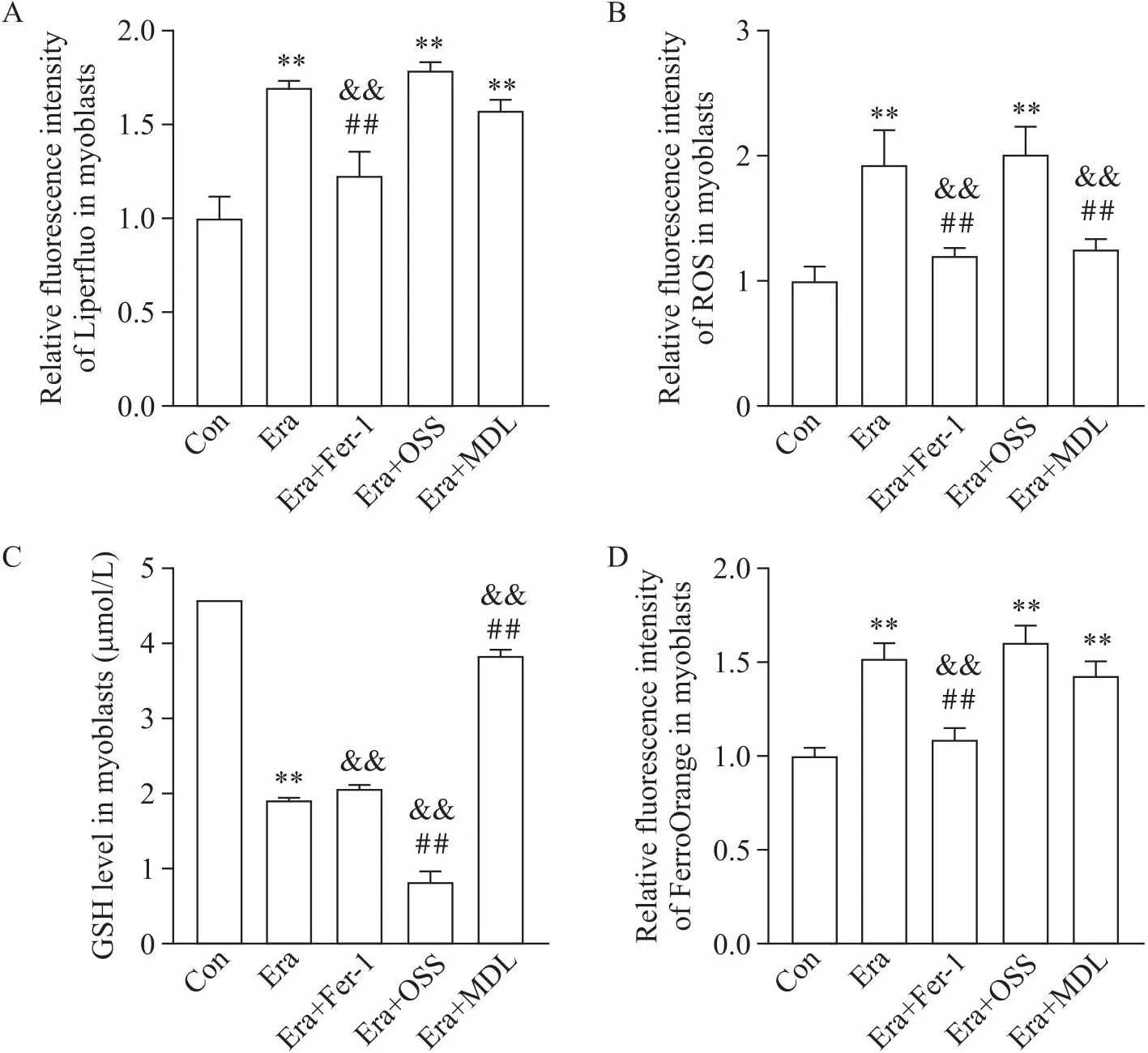
Figure 5. Lipid peroxidation indicators, Liperfluo (A), ROS (B), GSH (C) and FerroOrange/Fe2+(D), in C2C12 myoblasts treated with erastin (Era)-based addition of ferrostatin-1 (Fer-1), OSS-128167 (OSS) or MDL-800 (MDL). Mean±SD. n=3. **P<0.01 vs control (Con) group; ##P<0.01 vs Era group; &&P<0.01 vs Era+OSS group.
C2C12肌管用不同药物处理48 h,RT-qPCR显示Era使MAFbx和MuRF1的mRNA表达增加(图6A),铁死亡脂质过氧化指标和胞内Fe2+升高,GSH下降(图6B);OSS会加重铁死亡改变,但MDL与Fer-1相似,能部分逆转Era诱导的肌管变化(图6)。这些结果证实铁死亡降低肌管质量,使促肌蛋白分解分子表达增加,Sirt6的激活对此有抑制作用。
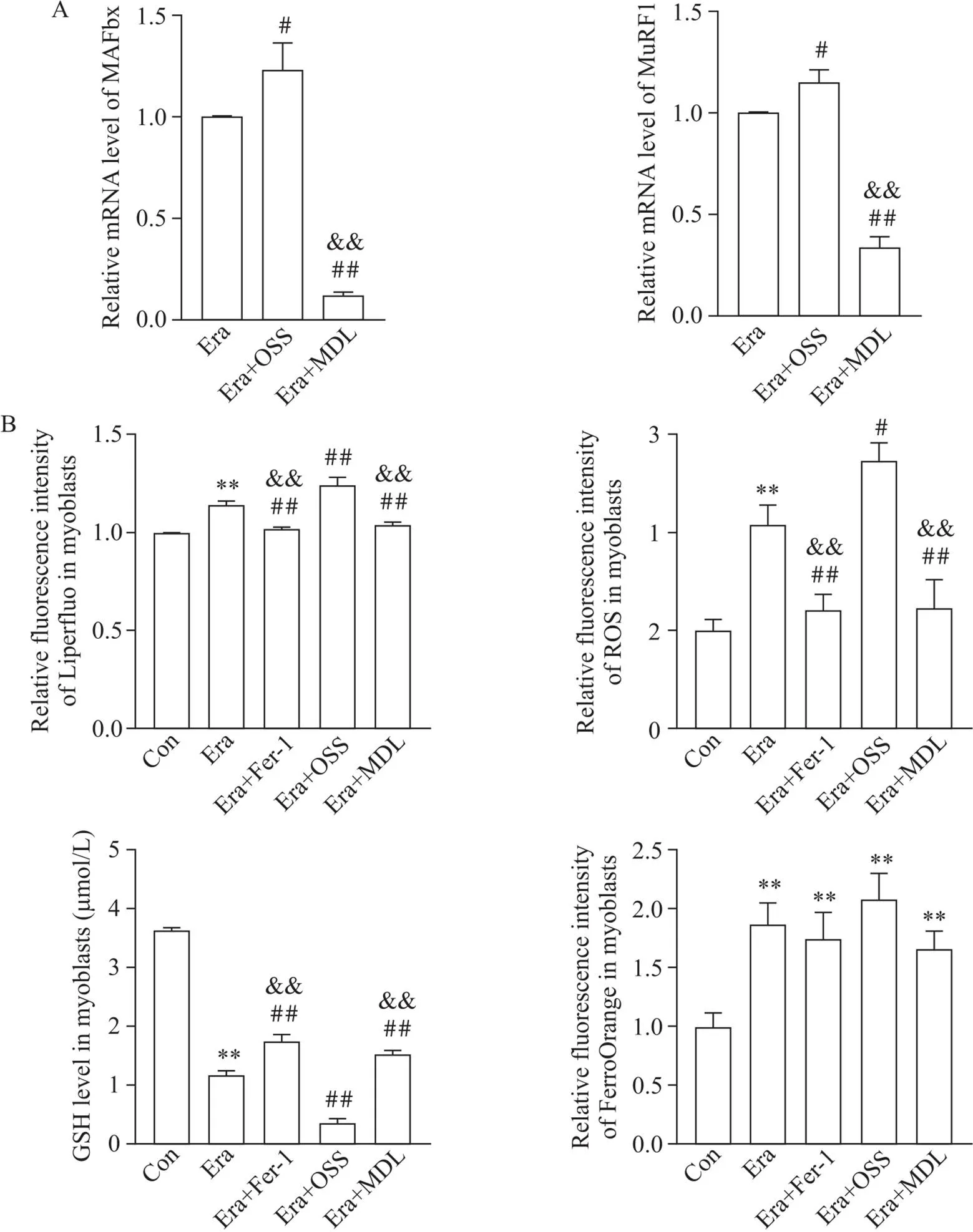
Figure 6. Changes of amatrophy-related genes (A) and lipid peroxidation indexes (B) in C2C12 myotubes after addition ferrostatin-1 (Fer-1), OSS-128167 (OSS) or MDL-800 (MDL) based on erastin (Era) treatment. Mean±SD. n=3. **P<0.01 vs control (Con) group; #P<0.05, ##P<0.01 vs Era group; &&P<0.01 vs Era+OSS group.
3.2Sirt6激活能部分恢复因Era受损的细胞肌源性分化能力使用含不同药物的分化培养液作用细胞5 d,利用免疫荧光染色检测肌分化末期标志分子MHC,观察肌管分化的数量和形态。结果发现Era引起肌管胞核融合下降、数量变少、长度变短、直径变细、排列紊乱,MHC荧光信号降低;而MDL激活Sirt6后,与Fer-1相似,能减轻上述肌管变化(图7A),说明Sirt6能抑制铁死亡引起的肌源性分化能力退化。
3.3Sirt6的去乙酰化作用可降低P53表达,通过P53/SLC7A11/GPX4信号通路抑制铁死亡肌管变化用Era处理肌管48 h,Sirt6蛋白水平下降,P53、Ac-P53 (Lys381)和肌萎缩标志蛋白(MAFbx和MuRF1)表达增强,SLC7A11和GPX4下降;在此基础上加入MDL,则P53、Ac-P53 (Lys381)、MAFbx和MuRF1蛋白降低,SLC7A11和GPX4升高,与Fer-1相似,均能逆转肌管铁死亡相关蛋白表达;而加用OSS使P53、Ac-P53 (Lys381)、MAFbx和MuRF1表达进一步增加,Sirt6、SLC7A11和GPX4表达减少(图7B)。这些结果表明,Sirt6抑制铁死亡和改善铁死亡相关肌管改变是通过使Ac-P53 (Lys381)去乙酰化而降低P53活性,进而影响下游P53/SLC7A11/GPX4信号通路来实现的。
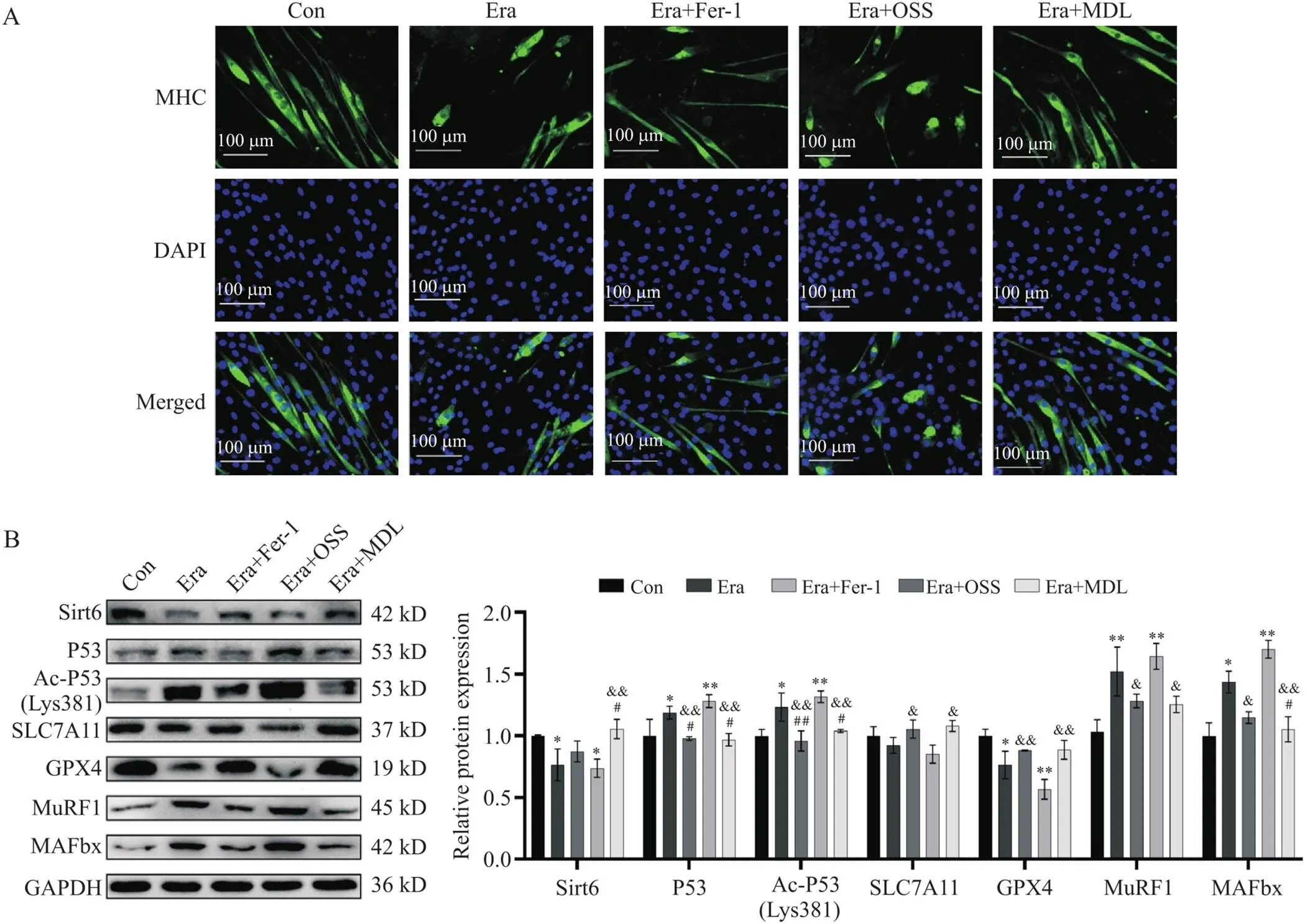
Figure 7. Effects of Sirt6 on MHC and P53/SLC7A11/GPX4 signaling pathway in C2C12 myotubes. A: immunofluorescence staining of MHC; B: the protein levels of P53, Ac-P53 (Lys381), SLC7A11, GPX4, MAFbx and MuRF1. Mean±SD. n=3. *P<0.05, **P<0.01 vs Con group; #P<0.05, ##P<0.01 vs Era group; &P<0.05, &&P<0.01 vs Era+OSS group.
此外,在Era作用C2C12成肌细胞或肌管后ROS水平增高;Fer-1及MDL使ROS生成有不同程度下降;联用OSS后ROS又进一步增加(图5B、6B)。因此,考虑Sirt6对肌细胞和肌管铁死亡变化的调控存在多个作用机制,除了去乙酰化作用外,对ROS生成的影响也可能辅助抑制铁死亡。
讨论
肌少症相关的研究已经成为老年化社会的一个重点方向,其发生机制已有大量研究报道[2, 22],随着对衰老研究的深入,铁死亡与肌少症的关联也走进我们的视野。C2C12细胞系属于卫星细胞(satellite cells, SCs),能够在充分刺激下从成肌细胞转化成可分化的肌纤维最终形成可伸缩的肌管,是用以研究肌肉分化和再生的理想体外模型。成肌分化在肌肉再生中起重要作用,通过高度协调的序列程序产生成熟的骨骼肌,因此成肌细胞活力、肌源性分化能力和肌管质量下降更能从源头反映肌肉功能、力量和质量改变[23]。本研究发现,使用Era处理后,C2C12成肌细胞发生铁死亡,肌管分化能力下降,可对骨骼肌产生负性影响。
铁作为人体重要的微量元素,伴随年龄增长在身体机能调节中成为一把双刃剑[9, 24],让铁死亡从多方面对骨骼肌质量产生影响。发生铁死亡的肌细胞中,转铁蛋白受体1(transferrin receptor 1, Tfr1)明显减少会影响SCs蛋白合成,使SCs不可逆消耗,导致肌细胞活力下降、骨骼肌再生能力变差甚至肌萎缩发生[25]。肌细胞中铁积累增加,GPX4和核因子E2相关因子2蛋白水平降低导致细胞脂质过氧化、ROS清除障碍,加重肌细胞损伤[26-27]。P53在细胞铁死亡中也发挥重要作用,应激条件下被大量激活,介导下游靶基因转录抑制,调节System XC-、GPX4促进肌细胞铁死亡变化[12, 28-29]。此外,作为肌肉蛋白分解代谢最重要酶类,部分E3泛素连接酶参与铁死亡发生[30],而肌萎缩特异性E3泛素连接酶(MAFbx和MuRF1)在衰老个体骨骼肌中随铁累积表达增加[9, 31]。研究中也发现Era处理后MHC表达下降,MAFbx和MuRF1含量大幅升高,铁死亡影响肌管分化,促进肌肉蛋白分解。
Sirt6是参与衰老、能量代谢、炎症等反应的重要脱乙酰酶,缺乏的个体更容易出现体重减少、脊柱前凸、肿瘤发生率增加、加速衰老,甚至过早死亡。目前Sirt6与铁死亡的关系多数是围绕肿瘤相关的报道,但在非肿瘤领域中也发现Sirt6通过抑制炎症,减少1型糖尿病小鼠前额叶皮层细胞的铁死亡,改善1型糖尿病小鼠的抑郁和焦虑[32]。
结合P53在铁死亡调控中的重要意义和乙酰化作用能加强P53蛋白表达的特性[19, 33],本研究针对Era处理后的C2C12成肌细胞或肌管引入Sirt6特异性激动剂MDL[34]和抑制剂OSS[35],发现激动Sirt6能够改善成肌细胞中铁死亡变化并降低肌萎缩特异性E3泛素连接酶的表达。
对P53蛋白C末端第381位赖氨酸这一关键乙酰化位点进一步研究发现,Sirt6激活使P53中第381位赖氨酸去乙酰化导致P53活性减弱,SLC7A11和GPX4的mRNA和蛋白表达增加,铁死亡和肌萎缩特异性指标下降,肌源性分化能力得到一定恢复。相反,Sirt6抑制会进一步加重铁死亡引起的变化。这些结果表明,Sirt6通过去乙酰化作用,可以影响P53/SLC7A11/GPX4信号通路表达,是铁死亡相关肌少症的重要调控分子。
总之,在这项研究中,我们证明了骨骼肌细胞中铁死亡发生与P53/SLC7A11/GPX4通路中信号变化关系密切,并伴有Sirt6表达下调,肌管质量下降,最终可能导致肌少症发生,引起肌肉功能障碍;并从机制的角度阐明Sirt6对成肌细胞和肌管铁死亡的干预是通过促进Ac-P53 (Lys381)去乙酰化,从而影响P53/SLC7A11/GPX4信号通路实现的。我们的研究结果为铁死亡相关肌少症这一新型“流行病”治疗提供了新的思考方向和治疗策略。
[1] Cruz-Jentoft AJ, Sayer AA. Sarcopenia[J]. Lancet, 2019, 393(10191):2636-2646.
[2] Cruz-Jentoft AJ, Bahat G, Bauer J, et al. Sarcopenia: revised European consensus on definition and diagnosis[J]. Age Ageing, 2019, 48(1):16-31.
[3] Woo EC, Rodis B. Sarcopenia in elderly surgery[J]. Ann Acad Med Singap, 2019, 48(11):363-369.
[4] Wiedmer P, Jung T, Castro JP, et al. Sarcopenia: molecular mechanisms and open questions[J]. Ageing Res Rev, 2021, 65:101200.
[5] Kinoshita H, Orita S, Inage K, et al. Skeletal muscle cell oxidative stress as a possible therapeutic target in a denervation-induced experimental sarcopenic model[J]. Spine (Phila Pa 1976), 2019, 44(8):E446-E455.
[6] Alves FM, Kysenius K, Caldow MK, et al. Iron accumulation in skeletal muscles of old mice is associated with impaired regeneration after ischaemia-reperfusion damage[J]. J Cachexia Sarcopenia Muscle, 2021, 12(2):476-492.
[7] Zhao G. Is iron accumulation a possible risk factor for sarcopenia?[J]. Biol Trace Elem Res, 2018, 186(2):379-383.
[8] Alves FM, Ayton S, Bush AI, et al. Age-related changes in skeletal muscle iron homeostasis[J]. Gerontol A Biol Sci Med Sci, 2023, 78(1):16-24.
[9] Huang Y, Wu B, Shen D, et al. Ferroptosis in a sarcopenia model of senescence accelerated mouse prone 8 (SAMP8)[J]. Int J Biol Sci, 2021, 17(1):151-162.
[10] Ikeda Y, Imao M, Satoh A, et al. Iron-induced skeletal muscle atrophy involves an Akt-forkhead box O3-E3 ubiquitin ligase-dependent pathway[J]. J Trace Elem Med Biol, 2016, 35:66-76.
[11] Stockwell BR, Friedmann Angeli JP, Bayir H, et al. Ferroptosis: a regulated cell death nexus linking metabolism, redox biology, and disease[J]. Cell, 2017, 171(2):273-285.
[12] Jiang L, Kon N, Li T, et al. Ferroptosis as a p53-mediated activity during tumour suppression[J]. Nature, 2015, 520(7545):57-62.
[13] Wang SJ, Li D, Ou Y, et al. Acetylation is crucial for p53-mediated ferroptosis and tumor suppression[J]. Cell Rep, 2016, 17(2):366-373.
[14] 张丽媛, 李芙蓉, 王超, 等. p53对铁死亡的调节作用及潜在应用[J]. 中国病理生理杂志, 2019, 35(12):2299-2304.
Zhang LY, Li FR, Wang C, et al. Emerging roles of p53 in ferroptosis and its potential application[J]. Chin J Pathophysiol, 2019, 35(12):2299-2304.
[15] Chang AR, Ferrer CM, Mostoslavsky R. SIRT6, a mammalian deacylase with multitasking abilities[J]. Physiol Rev, 2020, 100(1):145-169.
[16] Liu G, Chen H, Liu H, et al. Emerging roles of SIRT6 in human diseases and its modulators[J]. Med Res Rev, 2021, 41(2):1089-1137.
[17] 陶娜娜, 周洪钟, 任吉华, 等. Sirtuin 6对肝癌细胞增殖的影响[J]. 中国病理生理杂志, 2016, 32(6):1031-1036.
Tao NN, Zhou HZ, Ren JH, et al. Effect of sirtuin 6 on proliferation of hepatocellular carcinoma cells[J]. Chin J Pathophysiol, 2016, 32(6):1031-1036.
[18] Cai S, Fu S, Zhang W, et al. SIRT6 silencing overcomes resistance to sorafenib by promoting ferroptosis in gastric cancer[J]. Biochem Biophys Res Commun, 2021, 577:158-164.
[19] Gong S, Xiong L, Luo Z, et al. SIRT6 promotes ferroptosis and attenuates glycolysis in pancreatic cancer through regulation of the NF-κB pathway[J]. Exp Ther Med, 2022, 24(2):502.
[20] Xia Z, Kon N, Gu AP, et al. Deciphering the acetylation code of p53 in transcription regulation and tumor suppression[J]. Oncogene, 2022, 41(22):3039-3050.
[21] Ghosh S, Wong S K, Jiang Z, et al. Haploinsufficiency ofdramatically extends the lifespan of Sirt6-deficient mice[J]. ELife, 2018, 7:e32127.
[22] Chen LK, Liu LK, Woo J, et al. Sarcopenia in Asia: consensus report of the Asian Working Group for Sarcopenia[J]. J Am Med Dir Assoc, 2014, 15(2):95-101.
[23] Yaffe D, Saxel O. Serial passaging and differentiation of myogenic cells isolated from dystrophic mouse muscle[J]. Nature, 1977, 270(5639):725-727.
[24] Tang D, Chen X, Kang R, et al. Ferroptosis: molecular mechanisms and health implications[J]. Cell Res, 2021, 31(2):107-125.
[25] Ding H, Chen S, Pan X, et al. Transferrin receptor 1 ablation in satellite cells impedes skeletal muscle regeneration through activation of ferroptosis[J]. J Cachexia Sarcopenia Muscle, 2021, 12(3):746-768.
[26] Gong Y, Wang N, Liu N, et al. Lipid peroxidation and GPX4 inhibition are common causes for myofibroblast differentiation and ferroptosis[J]. DNA Cell Biol, 2019, 38(7):725-733.
[27] Wang Y, Yu R, Wu L, et al. Hydrogen sulfide guards myoblasts from ferroptosis by inhibiting ALOX12 acetylation[J]. Cell Signal, 2021, 78:109870.
[28] Hassannia B, Vandenabeele P, Vanden Berghe T. Targeting ferroptosis to iron out cancer[J]. Cancer Cell, 2019, 35(6):830-849.
[29] Liu Y, Gu W. p53 in ferroptosis regulation: the new weapon for the old guardian[J]. Cell Death Differ, 2022, 29(5):895-910.
[30] Meng Y, Sun H, Li Y, et al. Targeting ferroptosis by ubiquitin system enzymes: a potential therapeutic strategy in cancer[J]. Int J Biol Sci, 2022, 18(14):5475-5488.
[31] Bodine SC, Baehr LM. Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1[J]. Am J Physiol Endocrinol Metab, 2014, 307(6):E469-E484.
[32] Wang Y, Wang S, Xin Y, et al. Hydrogen sulfide alleviates the anxiety-like and depressive-like behaviors of type 1 diabetic mice via inhibiting inflammation and ferroptosis[J]. Life Sci, 2021, 278:119551.
[33] Xia Z, Kon N, Gu AP, et al. Deciphering the acetylation code of p53 in transcription regulation and tumor suppression[J]. Oncogene, 2022, 41(22):3039-3050.
[34] Shang JL, Ning SB, Chen YY, et al. MDL-800, an allosteric activator of SIRT6, suppresses proliferation and enhances EGFR-TKIs therapy in non-small cell lung cancer[J]. Acta Pharmacol Sin, 2021, 42(1):120-131.
[35] Jiang H, Cheng ST, Ren JH, et al. SIRT6 inhibitor, OSS_128167 restricts hepatitis B virus transcription and replication through targeting transcription factor peroxisome proliferator-activated receptors α[J]. Front Pharmacol, 2019, 10:1270.
Sirtuin 6 suppresses ferroptosis in skeletal muscle cells by regulating P53/SLC7A11/GPX4 pathway
WANG Yao1, CHEN Shikun1, DUAN Chenyang1, LIANG Xiaohan1, ZHOU Chengfu1, QIN Jun1, HOU Dongyao1, DU Quan1,2△
(1,,400010,;2,400010,)
To investigate the occurrence of ferroptosis in skeletal muscle cells and its molecular mechanisms.Mouse C2C12 myoblasts were divided into control group, erastin (Era; ferroptosis inducer) group, Era+ferrostatin-1 (ferroptosis antagonist) group, Era+MDL-800 [sirtuin 6 (Sirt6) agonist] group, and Era+OSS-128167 (Sirt6 inhibitor) group. The mRNA and protein expression levels of Sirt6, muscle atrophy markers and ferroptosis markers in C2C12 myoblasts or myotubes were determined by RT-qPCR and Western blot. Furthermore, acetylation levels of P53 protein, intracellular ferrous ion (Fe2+), reactive oxygen species (ROS), glutathione (GSH) and lipid peroxidation index Liperfluo were examined. Immunofluorescence revealed the intensity of myosin heavy chain (MHC; a biomarker of myotube differentiation) fluorescence signal.Treatment with Era reduced the viability of C2C12 cells and the quality of differentiated C2C12 myotube, accompanied by decreased Sirt6 mRNA and protein levels (<0.05). The mRNA and protein levels of muscle atrophy F-box protein (MAFbx) and muscle ring-finger protein 1 (MuRF1), markers of amyotrophy, were increased (<0.05). Activation of Sirt6 inhibited acetylation of lysine 381 site in P53 protein, and decreased P53 expression. The Sirt6 agonist also increased solute carrier family 7 member 11 (SLC7A11) and glutathione peroxidase 4 (GPX4) expression levels, and altered ferroptosis specific indicators such as ROS, GSH, Fe2+and lipid peroxidation (<0.05). Conversely, inhibition of Sirt6 promoted ferroptosis.Sirt6 inhibits the acetylation of P53 protein to reduce its activity, counteracts ferroptosis-associated cell death and improves muscle mass by regulating the P53/SLC7A11/GPX4 signaling pathway.
sarcopenia; sirtuin 6; ferroptosis; deacetylation
R322.7+4; R363.2
A
10.3969/j.issn.1000-4718.2023.02.016
1000-4718(2023)02-0335-10
2022-09-19
2022-11-28
[基金项目]重庆市出国留学人员创新自主项目(No. CX2021069);重庆科卫联合项目(No. 2020MSXM008)
Tel: 13883643222; E-mail: duquan100@sina.com
(责任编辑:林白霜,罗森)
