维生素A对一氧化氮诱导损伤的奶牛乳腺上皮细胞内乳蛋白合成下降的缓解作用
2023-03-07闫素梅刘锦涛杨青苗郭晓宇郭咏梅郑亚光赵艳丽
李 愿 闫素梅 刘锦涛 杨青苗 郭晓宇 郭咏梅 郑亚光 赵艳丽
(内蒙古农业大学动物科学学院,内蒙古自治区高校动物营养与饲料科学重点实验室,呼和浩特010018)
乳蛋白作为牛乳中的重要组成部分,是衡量乳品质的重要指标。奶牛饲粮营养水平、乳腺的健康状况以及其他调控乳蛋白合成的调控因子的变化,均会引起乳蛋白含量的变化。乳腺组织作为合成乳蛋白的重要场所,是奶牛体内代谢旺盛的组织之一,短时间内产生大量活性氧(ROS)、活性氮(RNS)自由基,超高水平的ROS等自由基导致乳腺内氧化还原失衡,产生氧化应激,进而破坏有机大分子发生代谢紊乱,诱导疾病产生[1]。长时间的氧化应激使奶牛乳腺代谢紊乱,导致产奶量、乳品质的下降,增加患乳腺炎的风险,影响经济效益。维生素A在维持机体正常生理功能中起着重要作用,通过对多种通路的调控影响抗氧化、免疫刺激、上皮细胞维持等功能[2]。体内外低浓度的一氧化氮(NO)有抗菌、抗肿瘤作用,但过高浓度的NO则会导致机体产生氧化应激[3]。有学者提出,维生素A对NO的调节可能是通过上调硫氧还蛋白还原酶(TrxR)抑制诱导型氧化氮合酶(iNOS)的活性,进而影响NO的生成,但其机理尚不清晰[4]。因此,探讨维生素A缓解奶牛乳腺上皮细胞(BMECs)氧化损伤以及乳蛋白合成下降的机理,对提高产奶量、乳品质以及延长奶牛的使用寿命具有重要意义。本课题组前期研究发现,与饲喂推荐剂量(110 IU/kg BW)的维生素A组相比,饲喂高剂量(220 IU/kg BW)的维生素A显著提高了奶牛血清中谷胱甘肽过氧化物酶(GPx)和TrxR的活性,并显著降低血清中ROS的含量,有增加乳蛋白含量的趋势[5]。体外研究显示,维生素A可缓解脂多糖对BMECs的氧化损伤,提高细胞内GPx、TrxR的活性和基因表达[6]。对BMECs的体外研究发现,维生素A具有促进乳蛋白及其合成相关基因表达的作用[7]。然而,维生素A是否能通过缓解氧化损伤促进乳蛋白合成还未见相关报道。因此,本试验采用不同浓度的维生素A进行预处理,来探究其对NO诱导损伤的BMECs内乳蛋白合成的影响及其机理,并筛选适宜的维生素A添加剂量,为深入研究乳蛋白合成的调控机理提供理论依据。
1 材料与方法
1.1 细胞培养及试验设计
原代BMECs的培养采用胶原酶消化法,步骤参照Qi等[8]的方法进行。采用单因子完全随机试验设计,以二乙烯三胺/一氧化氮聚合物(DETA/NO)作为氧化应激源,将第3代BMECs细胞混匀后分为对照组(CON组)、NO损伤组(NO组)以及NO损伤+不同浓度(0.05、0.10、0.20、0.50、1.00、2.00、4.00 μg/mL)维生素A预保护组(NA0.05、NA0.1、NA0.2、NA0.5、VA1、VA2和NA4组),每组4个重复。第3代BMECs细胞在5%CO2、37 ℃条件下培养,待细胞融合至80%~90%后,各组更换无血清培养基饥饿培养12 h后,CON组不添加维生素A和DETA/NO继续培养30 h;NO组以不添加维生素A的培养基培养24 h后,添加1 000 μmol/L DETA/NO继续培养6 h;NA0.05、NA0.1、NA0.2、NA0.5、VA1、VA2和NA4组分别以含0.05、0.10、0.20、0.50、1.00、2.00、4.00 μg/mL维生素A的培养基培养24 h后,添加1 000 μmol/L DETA/NO继续培养6 h。处理结束后,收集细胞以及细胞培养液进行抗氧化指标、乳蛋白合成相关蛋白含量和酶活性以及抗氧化和乳蛋白合成相关基因表达的测定。
1.2 配制试剂
维生素A原液:将50 mg的视黄酸(Sigma公司,美国)溶解于2.5 mL二甲基亚砜(DMSO,Sigma公司,美国),配制成20 mg/mL的维生素A原液,0.22 μm滤器过滤后避光冷冻保存。
NO贮备液:10 mg的DETA/NO溶于612.8 μL的无菌蒸馏水中,配成浓度为100 mmol/L的贮备液。
1.3 测定指标与方法
1.3.1 细胞活力
采用CCK-8法测定BMECs的细胞活力,结果用相对增殖率(RGR)表示。将细胞悬液接种于96孔板中,按照试验设计加入不同工作液处理细胞30 h后,每孔加20 μL的CCK-8溶液(碧云天),培养箱中继续培养1 h,全自动酶标仪检测450 nm波长处的吸光度(OD450 nm)值,计算RGR。
RGR(%)=(试验组OD450 nm值/
对照组OD450 nm值)×100。
1.3.2 抗氧化指标及乳蛋白合成相关蛋白含量和酶活性
将BMECs以3×105个/mL的密度接种于60 mm培养皿内,根据试验设计加入不同工作液处理细胞30 h后,收集1 mL细胞培养液,采用钼酸铵显色法测定总抗氧化能力(T-AOC),采用黄嘌呤氧化酶法测定总超氧化物歧化酶(T-SOD)活性,采用比色法测定过氧化氢酶(CAT)活性,采用酶联免疫吸附试验(ELISA)法测定NO、ROS含量和iNOS活性,具体操作步骤参照试剂盒说明书进行。
将培养皿置于冰上,弃去细胞上清,磷酸盐缓冲液(PBS)清洗细胞,加入适量细胞裂解液,冰浴30 min,刮取细胞转入1.5 mL Eppendorf离心管内,4 ℃、12 000 r/min离心10min,收集上清,采用二硫代二硝基苯甲酸法测定GPx活性,采用ELISA法测定TrxR、p70核糖体蛋白S6激酶1(S6K1)活性和哺乳动物雷帕霉素靶蛋白(mTOR)含量,具体操作步骤参照试剂盒说明书。
1.3.3 抗氧化和乳蛋白合成相关基因表达
将BMECs以2×105个/mL的密度接种于6孔板内,按照试验设计加入不同工作液处理细胞30 h后,使用Trizol法提取细胞内的总RNA[9],RNA反转录使用PrimeScriptRTReagentKit(TaKaRa)试剂盒并按说明书进行操作。利用实时荧光定量PCR仪(ABI-7500,ABI公司)测定细胞内抗氧化和乳蛋白合成相关基因的表达情况,详细步骤参照SYBY Premix Ex TaqTMⅡ(TaKaRa)试剂盒说明书。PCR所用引物序列见表1。以磷酸甘油醛脱氢酶(GAPDH)为内参基因,用2-△△Ct法计算目的基因[GPx1、TrxR、αs1-酪蛋白(CSN1S1)、β-酪蛋白(CSN2)、κ-酪蛋白(CSN3)、真核起始因子4E(eIF4E)、真核起始因子4E结合蛋白1(4EBP1)、mTOR、S6K1、酪氨酸激酶2(JAK2)、信号转导和转录激活因子5(STAT5)]的mRNA相对表达量。
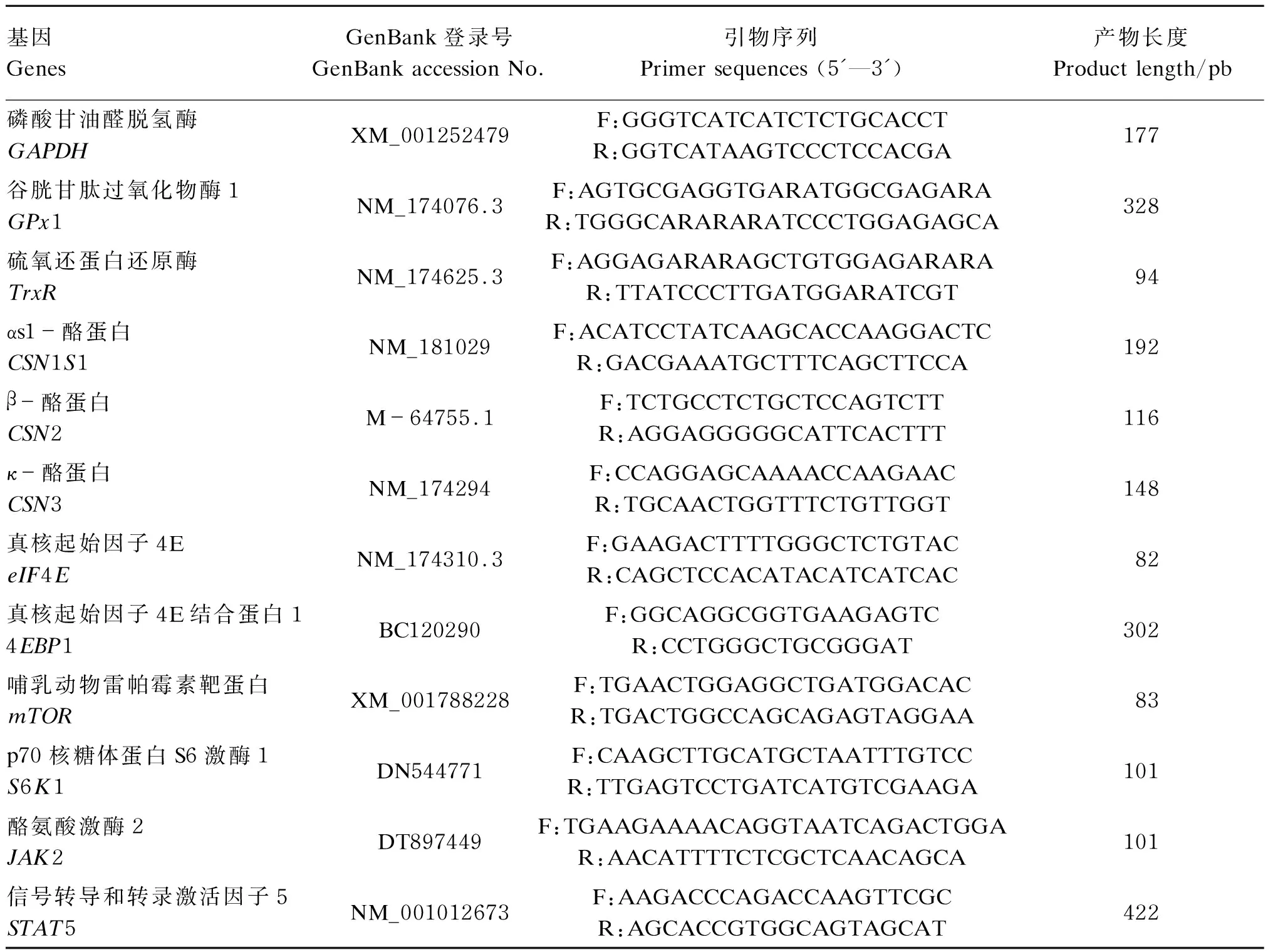
表1 引物序列
1.4 数据统计与分析
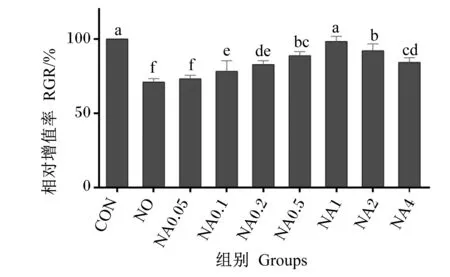
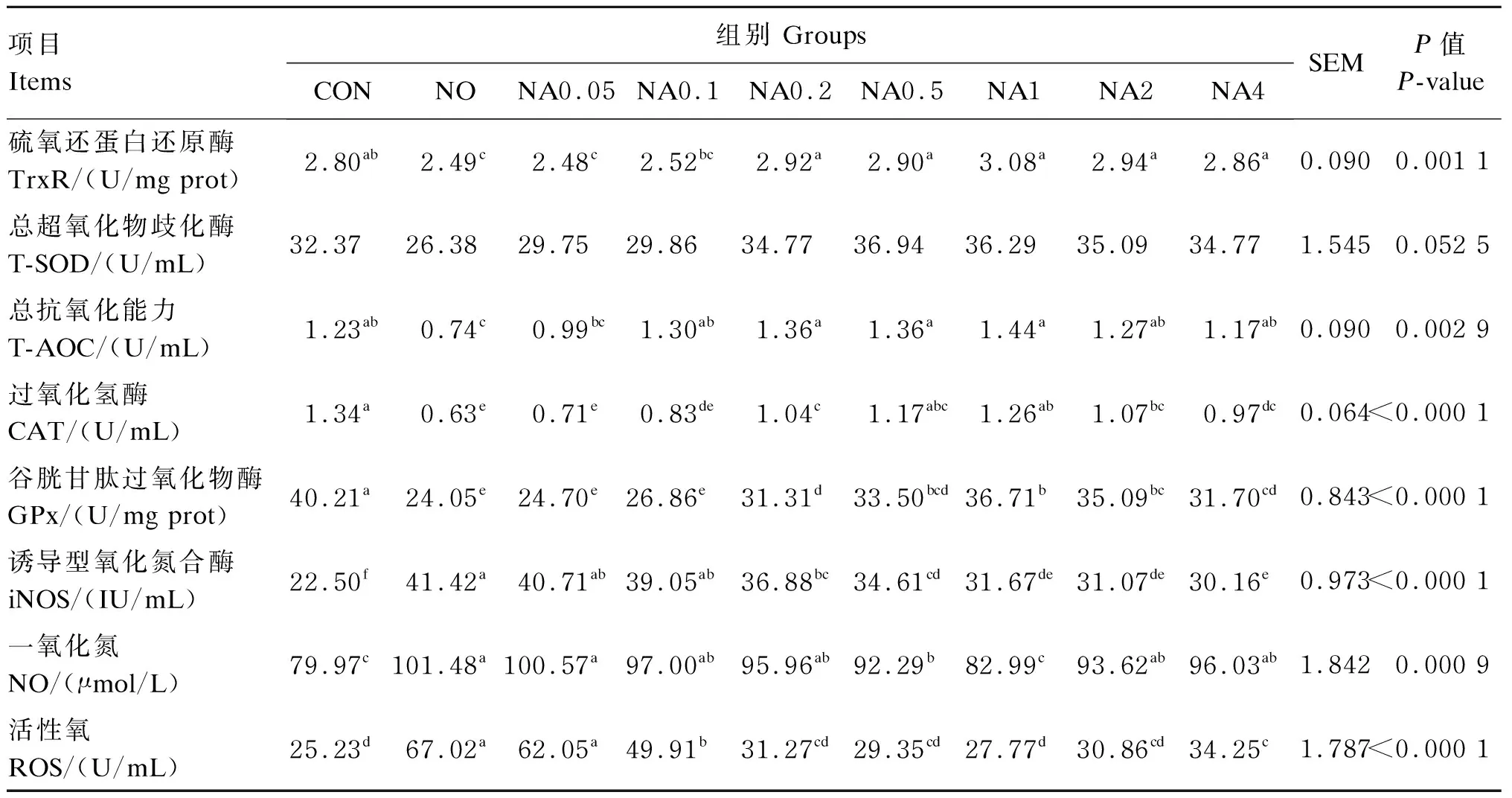
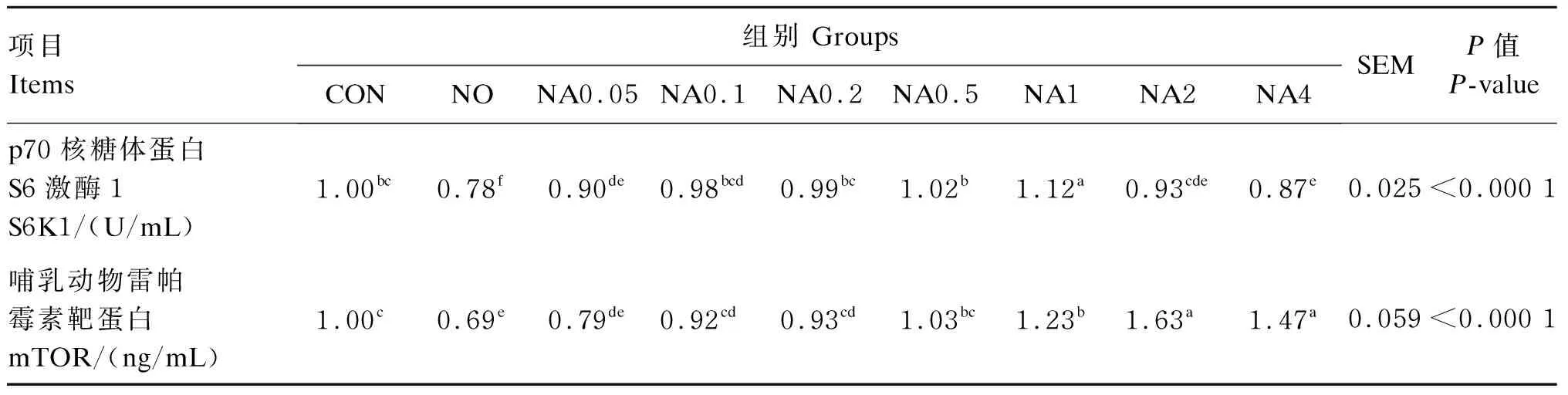
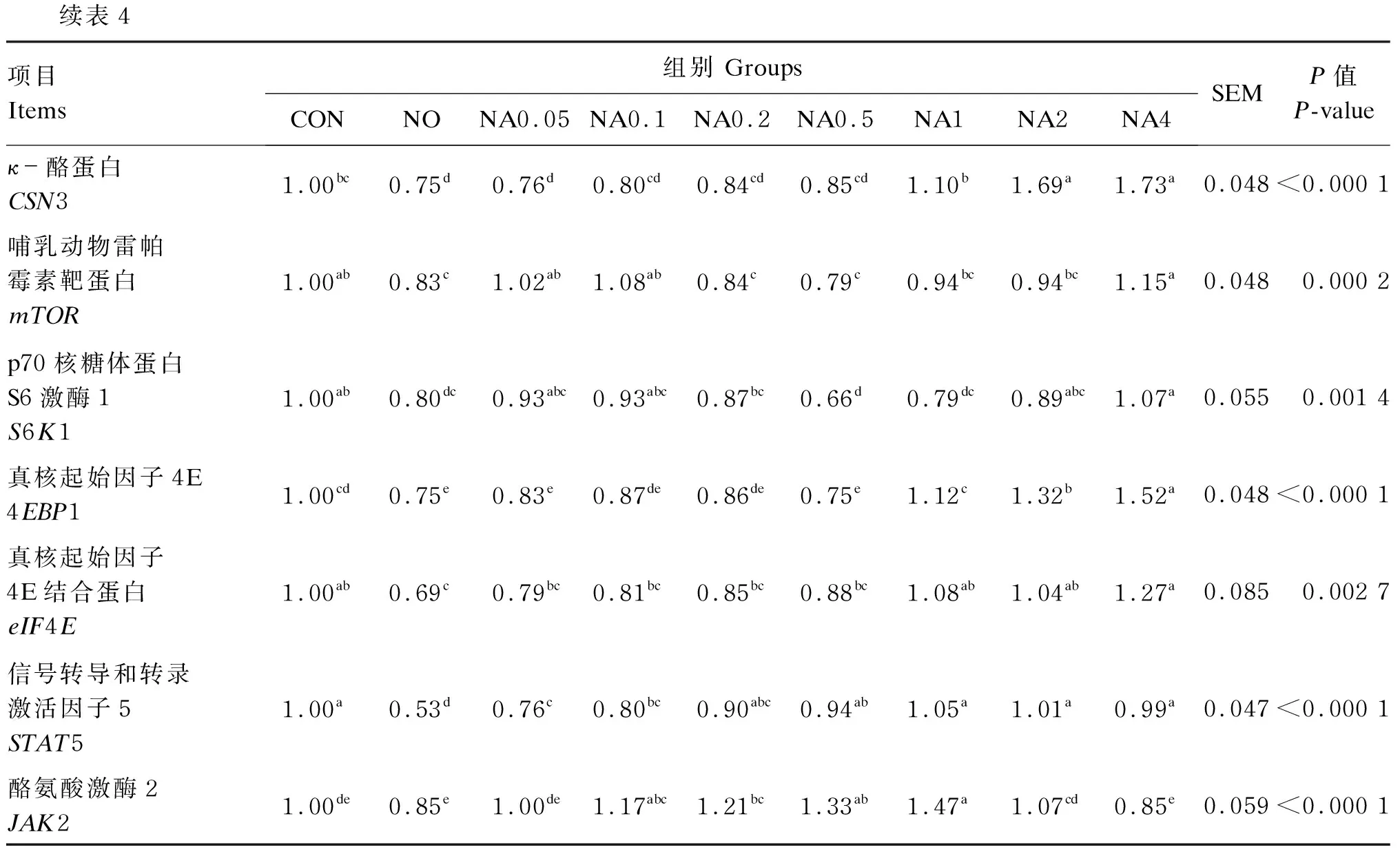
数据利用Excel 2019进行整理后,使用SAS 9.0软件进行单因素方差分析,以P≤0.05表示差异显著,P>0.05表示差异不显著,0.05 由图1可知,与CON组相比,NO组的RGR显著降低(P<0.05)。与NO组相比,NA0.1~NA4组的RGR显著升高(P<0.05),且以NA1组的RGR最高,达到CON组水平。 数据柱标注不同字母表示差异显著(P<0.05)。 由表2可知,与CON组相比,NO组的TrxR、CAT、GPx活性与T-AOC显著降低(P<0.05),iNOS活性与ROS、NO含量显著增加(P<0.05)。与NO组相比,NA0.2~NA4组的TrxR、CAT、GPx活性显著提高(P<0.05),NA0.1~NA4组的T-AOC显著提高(P<0.05),NA0.2~NA4组的iNOS活性显著降低(P<0.05),NA0.1~NA4组的ROS含量显著降低(P<0.05),NA0.5和NA1组的NO含量显著降低(P<0.05)。NA0.2~NA4组之间TrxR活性与T-AOC无显著差异(P>0.05);NA0.2~NA2组之间CAT活性无显著差异(P>0.05);NA1、NA2组的GPx活性无显著差异(P>0.05);NA4组的CAT、GPx活性显著低于NA1组(P<0.05);NA4组的NO、ROS含量显著高于NA1组(P<0.05);各组间T-SOD活性均无显著差异(P>0.05)。 表2 维生素A对NO诱导损伤的BMECs内抗氧化指标的影响 由表3可知,与CON组相比,NO组的S6K1活性和mTOR含量显著降低(P<0.05);NA1组的S6K1活性显著提高(P<0.05);NA1、NA2、NA4组的mTOR含量显著提高(P<0.05)。与NO组相比,NA0.05~NA4组的S6K1活性和mTOR含量均显著提高(P<0.05),且S6K1活性以NA1组最高,mTOR含量以NA2、NA4组较高。 表3 维生素A对NO诱导损伤的BMECs内乳蛋白合成相关蛋白含量和酶活性的影响 由表4可知,与CON组相比,NO组GPx1、TrxR、CSN2、CSN3、mTOR、S6K1、4EBP1、eIF4E、STAT5的mRNA相对表达量均显著降低(P<0.05),且均不同程度的低于NA0.05~NA4组。GPx1的mRNA相对表达量以NA1组最高,且显著高于其他组(P<0.05),同时NA4组显著低于NA0.1~NA1组(P<0.05);TrxR的mRNA相对表达量以NA1~NA4组较高,显著高于NO组和NA0.05~NA0.2组(P<0.05)。CSN1S1的mRNA相对表达量以NA0.2~NA1较高、CSN2的mRNA相对表达量以NA1组最高,均显著高于CON组和NO组(P<0.05);NA2、NA4组CSN3的mRNA相对表达量显著高于CON组、NO组和NA0.05~NA1组(P<0.05),NA1组显著高于NO组和NA0.05~NA0.5组(P<0.05);NA0.05、NA0.1和NA4组mTOR的mRNA相对表达量显著高于NO组(P<0.05),NA1~NA4组4EBP1、eIF4E的mRNA相对表达量显著高于NO组(P<0.05),mTOR、S6K1、4EBP1和eIF4E的mRNA相对表达量均以NA4组最高;NA0.5~NA4组STAT5的mRNA相对表达量显著高于NO组(P<0.05),NA0.1~NA2组JAK2的mRNA相对表达量显著高于NO组(P<0.05),STAT5和JAK2的mRNA相对表达量均以NA1组最高。 表4 维生素A对NO诱导损伤的BMECs内抗氧化和乳蛋白合成相关基因表达的影响 续表4项目Items组别 GroupsCONNONA0.05NA0.1NA0.2NA0.5NA1NA2NA4SEMP值 P-valueκ-酪蛋白CSN31.00bc0.75d 0.76d0.80cd0.84cd0.85cd1.10b 1.69a 1.73a 0.048<0.000 1哺乳动物雷帕霉素靶蛋白mTOR1.00ab0.83c 1.02ab1.08ab0.84c 0.79c 0.94bc0.94bc1.15a 0.0480.000 2p70核糖体蛋白S6激酶1S6K11.00ab0.80dc0.93abc0.93abc0.87bc0.66d 0.79dc0.89abc1.07a 0.0550.001 4真核起始因子4E4EBP11.00cd0.75e 0.83e 0.87de0.86de0.75e 1.12c 1.32b 1.52a 0.048 <0.000 1真核起始因子4E结合蛋白eIF4E1.00ab0.69c 0.79bc0.81bc0.85bc0.88bc1.08ab1.04ab1.27a 0.0850.002 7信号转导和转录激活因子5STAT51.00a 0.53d 0.76c 0.80bc0.90abc0.94ab1.05a 1.01a 0.99a 0.047<0.000 1酪氨酸激酶2JAK21.00de0.85e 1.00de1.17abc1.21bc1.33ab1.47a 1.07cd0.85e 0.059<0.000 1 乳腺组织泌乳代谢旺盛导致ROS、RNS生成的增加,诱导奶牛乳腺发生氧化应激。TrxR、T-SOD、CAT、GPx等酶是清除体内自由基的重要物质,其活性的降低导致机体的抗氧化能力下降,清除自由基的能力下降,致使大量自由基堆积[10-11]。前期本课题组研究显示,在奶牛饲粮中添加高于推荐剂量(220 IU/kg BW)的维生素A可明显提高奶牛的免疫、抗氧化能力,有提高乳蛋白含量的趋势[5];维生素A能通过提高TrxR、T-SOD、CAT、GPx等抗氧化酶的活性,降低ROS含量来缓解奶牛的氧化应激[12]。而有研究显示,添加高浓度的维生素A会导致妊娠大鼠的子宫和卵巢氧化损伤标志物含量增加,产生促氧化效应,使维生素A失去抗氧化能力[13]。本试验结果显示,维生素A浓度为4.00 μg/mL组CAT和GPx活性显著低于维生素A浓度为1.00 μg/mL组,GPx1的mRNA相对表达量显著低于维生素A浓度为0.10~1.00 μg/mL组,NO和ROS含量显著高于维生素A浓度为1.00 μg/mL组。这表明高浓度的维生素A可能通过降低抗氧化酶活性,增加ROS的含量,使维生素A失去抗氧化能力,进而导致维生素A缓解NO引起的氧化应激能力下降。体外研究验证了维生素A对过氧化氢(H2O2)、脂多糖(LPS)等外源性刺激诱导损伤的BMECs内的氧化应激有缓解作用[7,14]。本研究结果也得出相同的规律,即NO诱导损伤BMECs后,ROS、NO含量与iNOS活性显著升高,抗氧化酶TrxR和GPx等的活性降低,抗氧化相关基因GPx1和TrxR的表达下调,降低了细胞的活力和抗氧化能力;而添加不同浓度的维生素A进行预保护,不同程度地提高了由NO诱导损伤引起的细胞活力、抗氧化酶活性及抗氧化相关基因表达的下降,且效果以0.20~2.00 μg/mL较好,尤以1.00 μg/mL最好。 乳中的蛋白主要是酪蛋白,如CSN1S1、CSN2和CSN3,是衡量牛奶中乳蛋白含量的重要指标,与酪蛋白指数、乳蛋白的比例以及乳品质的稳定有关[15-17]。本试验中,NO诱导损伤导致乳蛋白合成相关基因CSN1S1、CSN2和CSN3的表达显著下调。酪蛋白基因表达下调可能会降低乳蛋白的比例以及乳品质的稳定。大量研究表明,蛋白质的合成主要通过转录和翻译2个方面从酪氨酸激酶/信号传导和转录激活因子(JAK/STAT)信号通路和mTOR信号通路进行调控。有研究发现阻断BMECs内的JAK/STAT信号通路,CSN2的表达显著下降,其他与乳蛋白合成相关的基因的表达均有下降趋势[8,18-20]。本试验结果表明,NO损伤导致了STAT5和JAK2表达的下调,添加不同浓度的维生素A进行预保护能缓解STAT5和JAK2表达的下调。mTOR信号通路主要从乳蛋白翻译水平发挥调控作用,下游通路的mTOR被激活后,磷酸化4EBP1使其与eIF4E分离,并同时激活S6K1,来促进蛋白质翻译及表达[21],尤其促进酪蛋白的翻译。Chen等[22]在培养多能性胚胎干细胞时添加维生素A后发现,其通过激活磷脂酰肌醇3-激酶(PI3K)调节维生素A介导的细胞增殖,进一步添加雷帕霉素和LY294002分别阻断mTOR及其上游PI3K,验证了维生素A可促进mTOR信号通路相关基因的表达。有研究发现添加维生素A促进了BMECs内TrxR的表达,进一步沉默TrxR后降低了JAK/STAT信号通路和mTOR信号通路相关基因的表达,以及mTOR信号通路的磷酸化水平[23]。Hu等[24]研究发现,培养小鼠骨髓间充质干细胞时敲除GPx7基因,mTOR信号通路相关因子S6K1和磷酸化S6K1蛋白的表达显著降低。也有研究表明JAK/STAT信号通路能调控炎症,葛根芩连汤可通过JAK/STAT信号通路降低ROS和其他炎症因子的产生,缓解小鼠结肠上皮的氧化应激[25]。这些结果表明,维生素A可能通过影响TrxR和GPx的活性进而调控JAK/STAT信号通路和mTOR信号通路。本试验结果发现,适宜浓度的维生素A对NO诱导损伤的BMECs内TrxR、GPx1、mTOR、S6K1、JAK2、STAT5、eIF4E、4EBP1、CSN1S1、CSN2和CSN3表达的下调具有缓解作用,可能通过上调TrxR和GPx的表达缓解JAK/STAT信号通路和mTOR信号通路活性的下降,进而缓解乳蛋白合成的下降。本研究也发现高浓度维生素A对GPx活性及GPx1表达下降的缓解作用减弱,对TrxR和mTOR表达下降的缓解作用较好,因此维生素A是如何通过缓解氧化应激调控JAK/STAT信号通路和mTOR信号通路还需进一步研究探讨。 综上所述,维生素A对NO诱导损伤的BMECs内乳蛋白合成下降的缓解作用与添加剂量相关,对JAK2、CSN1S1、CSN2表达下降的缓解作用以0.20~1.00 μg/mL较好,尤以1.00 μg/mL最好;对mTOR和S6K1表达下降的缓解作用以1.00 μg/mL最好;对eIF4E、4EBP1、STAT5以及CSN3表达下降的缓解作用以1.00~4.00 μg/mL较好。综合抗氧化指标、乳蛋白合成相关基因的表达来看,以0.20~2.00 μg/mL维生素A对NO诱导损伤的BMEC内乳蛋白合成下降的缓解作用较好,尤以1.00 μg/mL最好。 维生素A可以缓解NO引起的BMECs的细胞活力、抗氧化能力下降以及乳蛋白合成相关基因CSN1S1、CSN2、CSN3、STAT5、JAK2、mTOR、S6K1、eIF4E、4EBP1表达的下调,且缓解作用与添加剂量相关。综合比较,以0.20~2.00 μg/mL维生素A对NO诱导损伤的BMECs内乳蛋白合成下降的缓解作用较好,尤以1.00 μg/mL最好。2 结 果
2.1 维生素A对NO诱导损伤的BMECs的RGR的影响
2.2 维生素A对NO诱导损伤的BMECs内抗氧化指标的影响
2.3 维生素A对NO诱导损伤的BMECs内乳蛋白合成相关蛋白含量和酶活性的影响
2.4 维生素A对NO诱导损伤的BMECs内抗氧化和乳蛋白合成相关基因表达的影响

3 讨 论
4 结 论
