层积过程中羌活种子种胚形态及生理生化变化
2023-03-03其乐木格段海婧张金保陈红刚赵文龙
张 娟, 其乐木格, 段海婧,b, 张金保,b, 晋 玲,b,c, 陈红刚,b,c,①, 赵文龙,b,c,①
(甘肃中医药大学: a. 药学院, b. 西北中藏药省部共建协同创新中心, c. 甘肃省珍稀中药资源评价与保护利用工程研究中心, 甘肃 兰州 730000)
羌活(NotopterygiumincisumTing ex H. T. Chang)为中药羌活的来源之一。中药羌活别名有羌青、护羌使者、胡王使者、羌滑、退风使者和黑药等;性温,味辛、苦,入膀胱、肾经,具有解表散寒、祛风除湿、止痛的功效,用于风寒感冒、头痛项强、风湿痹痛、肩背酸痛,主产于甘肃、四川、青海等地,是中、藏、羌医药体系中的常用药[1-2]。2013年,羌活在由中华人民共和国环境保护部和中国科学院联合编制的《中国生物多样性红色名录——高等植物卷》中被收录为近危(NT)物种。
据记载,羌活单味药及复方制剂涉及30多种剂型,260多个成方,约280个品种。近年来,随着中医药大健康产业的迅速发展,羌活的市场需求量急剧增加,野生羌活资源濒临枯竭。因羌活种子休眠严重[3-4](自然状态下发芽率仅0.52%),目前只有部分地区试种成功,尚无法进行大规模人工种植。层积处理是常见的打破种子休眠的方法,可使种子种皮软化,增强种子内部代谢活动,还可促进种子完成一系列生理生化变化和后熟,变温层积处理是用高温与低温交替进行的层积方法,可以加速种子发芽[5]。如大百合〔Cardiocrinumgiganteum(Wall.) Makino〕[6]和北五味子〔Schisandrachinensis(Turcz.) Baill.〕[7]种子经变温层积后,种胚完成形态后熟而萌发。变温层积过程中营养物质含量、呼吸途径关键酶活性以及内源激素含量的变化与种胚发育密不可分。如变温层积过程中种子可溶性糖含量增加,淀粉和可溶性蛋白质含量降低[8],葡萄糖-6-磷酸脱氢酶(G-6-PDH)活性升高[9],脱落酸(ABA)含量降低,赤霉素(GA3)和吲哚乙酸(IAA)含量升高[10],均是种子休眠解除的原因之一。
羌活种子的休眠特性是其资源濒临枯竭的主要原因,与常温层积相比,变温(15 ℃~25 ℃/2 ℃~5 ℃)层积可缩短其解除休眠的时间[4]。目前,对羌活种子层积过程中种胚形态及生理生化变化的相关研究较少,仅对羌活种子种胚后熟过程中内源激素含量变化有所研究[11],对贮藏物质和呼吸途径关键酶并没有进行系统的研究,不利于羌活种子休眠解除机制的研究。鉴于此,本研究对层积过程中羌活种子种胚发育和生理生化指标的变化进行了研究,并分析了种胚发育与生理生化指标间的相关性,旨在明确羌活种子种胚发育和生理生化变化与种子休眠解除的内在联系,探究羌活种子萌发机制,建立种子休眠解除方法,为羌活野生资源保护、引种驯化、规模化种植提供参考依据。
1 材料和方法
1.1 材料
供试羌活种子于2020年9月采自甘肃省甘南藏族自治州玛曲县欧拉秀玛乡(具体地理坐标为东经101°06′28.46″、北纬34°15′02.64″,海拔3 471 m),由甘肃中医药大学晋玲教授鉴定为羌活种子,采摘后去除杂质,先用质量分数0.2%次氯酸钠消毒5 min,再用蒸馏水冲洗3~5次,于室内通风处阴干,分装于10 cm×15 cm网袋(40目)中,共分装14袋,其中7袋每袋1 g,编号0-F至6-F,剩余7袋每袋3 g,编号0-J至6-J,备用。
河沙来源于兰州市黄河边(具体地理坐标为东经103°48′52.48″、北纬34°03′57.21″),用水洗去泥土及杂质,185 ℃烘干约3 h,过40目筛,高压灭菌锅121 ℃灭菌30 min,加入无菌水将沙子湿度保持在9%~11%,备用。
1.2 方法
1.2.1 变温层积处理 参考文献[9]中的方法进行变温层积处理。在容器(30 cm×40 cm聚丙烯盒)底部平铺厚度4 cm的沙子,将分装好的羌活种子平铺在其表面,再盖上一层沙子,用保鲜膜封住容器,用镊子打孔(12个)透气。先将容器置于室温(20 ℃~25 ℃)90 d,后移入3 ℃~5 ℃冰箱90 d。分别在层积0(即未层积处理,取出样品0-F和0-J)、30(取出样品1-F和1-J)、60(取出样品2-F和2-J)、90(取出样品3-F和3-J)、120(取出样品4-F和4-J)、150(取出样品5-F和5-J)和180 d(取出样品6-F和6-J)取样,取出后通风处阴干,将样品0-F至6-F置于4 ℃冰箱中保存,用于后续种胚形态及发芽指标的观测,样品0-J至6-J置于-80 ℃冰箱中保存,用于后续生理生化指标的测定。
1.2.2 种子种胚形态及胚率观测 样品0-F至6-F中,每份样品选取羌活种子30粒,蒸馏水浸泡12 h,先用OCT胶水进行常温包埋,然后置于-20 ℃条件下冷冻包埋5 min,采用Leica-CM1950型恒冷切片机(德国Leica公司)切片(厚度0.12 mm),在Leica DM500显微镜(德国Leica公司)下观察种子形态结构,并随机取20粒种子用显微镜测量种子的胚长(种胚底端至顶端的长度)和胚乳长(胚乳底端至顶端的长度),取平均值计算种子的胚率,计算公式为胚率=(胚长/胚乳长)×100%[10,12]。
1.2.3 种子生理生化指标测定 采用考马斯亮蓝法[16-17]测定羌活种子蛋白质含量,采用蒽酮比色法[16-17]测定可溶性糖和淀粉含量,采用丙酮酸激酶(PK)活性检测法[17-18]测定PK活性,采用硫酸甲酯吩嗪反应法[17-18]测定琥珀酸脱氢酶(SDH)活性,这5个指标均使用购自南京建成生物工程研究所的试剂盒测定,试剂盒批号分别为20210622、20210422、20210429、20210329和202210329。采用酶联免疫法[17-18]测定葡萄糖-6-磷酸脱氢酶(G-6-PDH)活性、赤霉素(GA3)含量、吲哚乙酸(IAA)含量和脱落酸(ABA)含量,这4个指标均采用购自上海钦成生物科技有限公司的ELISA试剂盒测定,试剂盒批号均为202103,并据此计算GA3/ABA比、IAA/ABA比和(GA3+IAA)/ABA比。每个处理重复测定3次。
1.2.4 种子发芽指标测定 预实验结果显示:层积后的羌活种子(空白对照)的发芽率不足10%,而添加400 mg·L-1氟啶酮处理的结果较好,故本试验选择400 mg·L-1氟啶酮处理羌活种子。样品0-F至6-F中,每份样品随机选取羌活种子150粒,用400 mg·L-1氟啶酮溶液(批号F809822、纯度99%,购于上海麦克林生化科技股份有限公司)浸泡24 h,蒸馏水冲洗3~5次,将种子上残留的氟啶酮溶液冲洗干净,播于沙床上,每培养皿(直径9 cm)播种50粒,3次重复。将培养皿置于光照度3 000 lx、光照时间12 h·d-1、温度20 ℃条件下培养,每日加入适量蒸馏水,记录萌发种子数(以胚根突破种皮为种子萌发标准),第12天统计发芽势,第25天计算发芽率,计算公式分别为发芽势=(第12天发芽种子数/供试种子数)×100%,发芽率=(发芽种子总数/供试种子数)×100%[12-15]。
1.3 数据统计和分析
采用EXCEL 2013软件对获得的实验数据进行整理和分析,采用SPSS 28.0软件进行单因素方差分析、多重比较和Pearson相关性分析,采用Origin 2021软件制图。
2 结果和分析
2.1 层积过程中羌活种子种胚生长发育动态
在整个层积过程中,羌活种子种胚不断发育,胚率总体持续升高,种胚形态变化见图1,胚率变化见图2。结果显示:未层积(层积0 d)时种胚为球形胚(图1-A),胚率为7.53%,种子仍处于休眠状态;层积30~90 d时,种胚发育缓慢(图1-B,C,D),胚率从13.33%升高至17.09%;层积120 d时,种胚发育为心形胚(图1-E),胚率显著升高至36.65%;层积150 d时,种胚分化出完整的胚芽、胚轴和胚根(图1-F),胚率显著升高至57.35%;层积180 d时,种胚发育为子叶型胚(图1-G),胚率显著升高至67.30%,种胚后熟过程完成。

Em: 种胚 Embryo.

不同小写字母表示不同层积时间处理在0.05水平差异显著Different lowercases indicate the significant difference among different stratification time treatments at 0.05 level.
2.2 层积过程中羌活种子生理生化指标的变化
2.2.1 种子营养物质含量的变化 结果(表1)显示:羌活种子的蛋白质含量在层积过程中总体呈降低的趋势,在层积0~120 d持续降低,在层积150 d时略升高,在层积180 d时降至最小值(22.39 mg·g-1),层积60~180 d的种子蛋白质含量与层积0 d(未层积)差异显著。可溶性糖含量在层积过程中波动较小,但在层积0~60 d呈降低的趋势,且在层积60 d时降至最小值(33.30 mg·g-1),此后波动升高,但均显著低于未层积处理。淀粉含量在层积过程中总体呈先降低后升高的趋势,在层积0~60 d呈降低的趋势,且层积60 d时降至最小值(12.93 mg·g-1),之后波动升高,但均显著低于未层积处理。说明层积处理可显著降低羌活种子中营养物质的含量,为种子发育提供物质和能量。
2.2.2 种子呼吸途径关键酶活性的变化 结果(表1)显示:羌活种子的丙酮酸激酶(PK)活性在层积30 d时较未层积处理显著升高,在层积30~90 d呈显著降低的趋势,且在层积90 d时降至最小值(79.25 U·g-1),之后逐渐升高,在层积180 d时升至最大值(264.17 U·g-1)。琥珀酸脱氢酶(SDH)活性在层积30 d时较未层积处理无显著变化,在层积60 d时显著高于层积0和30 d,在层积60~150 d呈波动降低的趋势,在层积180 d时升至最大值(39.70 U·g-1)。葡萄糖-6-磷酸脱氢酶(G-6-PDH)活性在层积30 d时较未层积处理显著降低,在层积30~120 d无显著变化,在层积150 d时降至最小值(10.06 U·g-1),在层积180 d时升至最大值(20.35 U·g-1)。说明层积处理可使羌活种子内部代谢活动增强,有利于种胚发育。
2.2.3 种子内源激素含量的变化 结果(表1)显示:羌活种子的赤霉素(GA3)含量在层积过程中总体呈先升高后降低的趋势,在层积0~120 d呈上升趋势,在层积150 d时降至最小值(59.15 ng·g-1),显著低于未层积处理,在层积180 d时升至67.90 ng·g-1。吲哚乙酸(IAA)含量在14.06~22.08 ng·g-1范围内小幅度波动,在层积60 d时达到最大值(22.08 ng·g-1),之后总体呈平缓降低的趋势。脱落酸(ABA)含量在层积过程中总体呈先升高后降低的趋势,在层积0~120 d无显著变化,层积150和180 d的ABA含量显著低于层积0~120 d,在层积150 d时降至最小值(82.39 ng·g-1),在层积180 d时升至91.11 ng·g-1,但与层积150 d差异不显著。
结果(表1)还显示:羌活种子的GA3/ABA比、IAA/ABA比和(GA3+IAA)/ABA比在层积0~90 d无显著变化,之后总体呈升高趋势;层积120~180 d的GA3/ABA比和(GA3+IAA)/ABA比显著高于层积0~90 d,但在层积120~180 d无显著变化。总体上看,在层积过程中,(GA3+IAA)/ABA比的变幅最大,GA3/ABA比的变幅次之,IAA/ABA比的变幅较小。说明层积处理可使羌活种子内促进种子萌发的激素含量升高,抑制种子萌发的激素含量降低,有利于种子萌发。
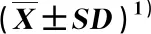
表1 层积过程中羌活种子生理生化指标的变化
2.3 层积过程中羌活种子发芽指标的变化
对层积过程中羌活种子发芽情况进行比较,结果见表2。结果显示:层积过程中羌活种子的发芽率和发芽势总体呈升高趋势,层积0~90 d的发芽率和发芽势较低,且均无显著变化,层积120~180 d的发芽率和发芽势显著高于层积0~90 d,发芽率和发芽势在层积180 d时升至最大值,分别为50.67%和35.33%。
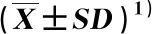
表2 层积过程中羌活种子发芽指标的变化
2.4 各指标间的相关性分析
对羌活种子的胚率和发芽率与各生理生化指标进行相关性分析,结果见表3。结果显示:种子胚率与脱落酸(ABA)含量呈显著负相关,相关系数为-0.800,与其余指标相关性不显著;发芽率与ABA含量呈极显著负相关,相关系数为-0.875,与其余指标相关性不显著。

表3 羌活种子胚率和发芽率与各生理生化指标间的相关系数
3 讨论和结论
3.1 羌活种子后熟与种胚形态的关系
因胚的发育程度造成的种子休眠可分为3种情况,即胚的形态休眠、生理休眠以及2种方式同时存在所引起的综合休眠[19]。综合休眠的种子在后熟阶段需要经历球形胚、心形胚、鱼雷型胚才能发育完全,这在柴胡(BupleurumchinenseDC.)[20]、滇重楼〔Parispolyphyllavar.yunnanensis(Franch.)Hand.-Mzt.〕[16]、刺楸〔Kalopanaxseptemiobus(Thunb.) Koidz.〕[21]等植物中已得到证实。本研究中,羌活种子先在室温(20 ℃~25 ℃)下层积90 d,后在低温(3 ℃~5 ℃)下继续层积90 d,观测发现羌活种子种胚发育具有胚后熟特性,且层积处理可有效促进种胚的发育,胚率总体持续升高,尤其是层积后期(低温层积)的胚率显著提高。张恩和等[4]的研究结果表明:羌活种子胚率仅6%,需经过一段时间的后熟,才可发育为具有胚根、胚轴和子叶的完整胚,且层积过程中低温可提高种胚的无氧呼吸作用,减少种胚的需氧量,从而克服包被组织造成的萌发障碍。由此推断,层积处理可打破羌活种子形态休眠,且低温对种胚形态休眠的解除至关重要。
3.2 羌活种子后熟与营养物质含量的关系
种子萌发过程中需要消耗能量,为了给种胚的生长发育提供原料和能量,种子中贮藏的淀粉、蛋白质和脂肪等大分子物质在相关酶的作用下会转化成种胚易于吸收的可溶性糖等小分子物质,这些小分子物质可为种胚的生长发育提供营养和能量[22-24]。黄精(PolygonatumsibiricumDelar. ex Redoute)[25]和百蕊草(ThesiumchinenseTurcz.)[26]种子在层积过程中,其淀粉含量逐渐降低,可溶性糖和蛋白质含量升高,大分子物质转化为小分子物质,为种胚的分化发育提供能量,促进了萌发。说明种子内贮藏物质的积累与消耗,可反应种子内部代谢活动的强弱[27]。本研究结果显示:在层积过程中羌活种子的蛋白质、可溶性糖和淀粉含量总体呈降低趋势。其中,蛋白质含量在层积0~120 d逐渐降低,在层积150 d时显著升高,在层积180 d时又显著降低,可能是因为在种子发育过程中蛋白质水解为氨基酸等其他小分子化合物,而后又和其他有机物质合成新的蛋白质,说明种子内部代谢活动增强[27]。可溶性糖和淀粉含量在层积0~60 d逐渐降低,为种胚伸长生长提供能量,在层积90~180 d波动升高,可能是为后期种子的萌发储存能量。
3.3 羌活种子后熟与呼吸途径关键酶活性的关系
糖酵解途径(EMP)、三羧酸循环途径(TCA)和磷酸戊糖途径(PPP)是植物体内常规的呼吸代谢途径,种子休眠和萌发与呼吸代谢途径密切相关,丙酮酸激酶(PK)、琥珀酸脱氢酶(SDH)和葡萄糖-6-磷酸脱氢酸(G-6-PDH)分别作为EMP途径、TCA途径和PPP途径的关键限速酶,一定程度上反映着相应呼吸途径的活化水平,通过对关键酶活性的测定,可以进一步了解种子休眠的历程[28-30]。如华重楼〔Parispolyphyllavar.chinensis(Franch.) Hara〕[31]种子在层积过程中G-6-PDH活性显著升高,其呼吸代谢途径从以TCA途径为主转为以PPP途径为主,通过调控种子呼吸代谢途径,促进种子解除休眠。本研究结果显示:在层积0~60 d, PK活性呈先升高后降低的趋势,TCA途径SDH活性呈明显升高的趋势,G-6-PDH活性先降低后略有升高,说明层积0~60 d期间以TCA途径为主。层积60~150 d,PK活性呈波动升高的趋势,SDH和G-6-PDH活性总体呈波动降低的趋势,说明层积60~150 d期间以EMP途径为主。在层积180 d时,PK、SDH和G-6-PDH活性均显著高于层积120 d,且层积150~180 d的羌活发芽率以及胚率均显著高于层积120 d,说明层积150~180 d的羌活种子内部代谢活动显著加强,利于种子休眠的解除。
3.4 羌活种子后熟与内源激素含量的关系
植物激素对种子萌发有至关重要的调控作用,脱落酸(ABA)可以促进种子休眠,抑制种子萌发。而赤霉素(GA3)可以解除种子休眠,促进种子萌发[32-33]。金银花(LonicerajaponicaThunb.)[34]和桃儿七〔Sinopodophyllumhexandrum(Royle) Ying〕[35]种子通过低温层积可使种子内部的GA3和IAA含量升高,ABA含量降低,降低了抑制种子萌发的内源激素含量,进而解除种子休眠。本研究结果表明:羌活种子的GA3和ABA含量在层积0~120 d内无显著差异,在层积150 d时显著降低,在层积180 d时又有所升高,而吲哚乙酸(IAA)含量在层积过程中变化不大。影响种子休眠和萌发的因子不仅与植物内源激素的绝对含量有关,还可能与种子内源激素的协调有关,尤其是与促进种子萌发和抑制种子萌发的激素有关[36],如高水平的GA3/ABA比、IAA/ABA比和(GA3+IAA)/ABA比有助于打破种子休眠[37-38]。本研究结果表明:羌活种子的GA3/ABA比和(GA3+IAA)/ABA比在层积过程中总体呈升高趋势,且层积120~180 d的GA3/ABA比和(GA3+IAA)/ABA比显著高于层积0~90 d,而IAA/ABA比在层积过程中无显著变化。相关性分析结果表明:种子胚率与ABA含量呈显著负相关,发芽率与ABA含量呈极显著负相关。说明层积处理可使种子内促进种子萌发的激素含量升高,抑制种子萌发的激素含量降低,提高种子的发芽率;且促进种子萌发与抑制种子萌发的激素比值显著升高对羌活种子休眠解除具有重要作用。
3.5 层积处理与羌活种子发芽情况的关系
种子的发芽情况可通过发芽率和发芽势等指标进行评价,发芽率和发芽势是检测种子质量的重要指标。氟啶酮为脱落酸(ABA)生物合成抑制剂,具有促进种子萌发的作用[39]。本研究结果表明:层积处理可促进种子萌发,将层积处理后的种子用氟啶酮处理,发芽势达到35.33%,种子发芽率达到50.67%。说明层积处理结合氟啶酮处理是打破羌活种子休眠的有效方法。
综上所述,羌活种子具有胚后熟特性,层积处理可打破羌活种子形态休眠,结合氟啶酮处理可提高种子发芽率;营养物质含量的降低,呼吸途径关键酶活性的波动变化,ABA含量的降低以及内源激素含量间的比值变化对羌活种子休眠解除有重要影响。该结果将为羌活引种驯化和规模化种植提供科学依据。
