‘中山含笑’与台湾含笑母树及其无明显表型变异子代的抗寒性分析
2023-03-03殷云龙王芝权陈婷婷华建峰汪贵斌
邱 健, 殷云龙, 王芝权, 施 钦, 陈婷婷, 华建峰, 汪贵斌
〔1. 南京林业大学南方现代林业协同创新中心, 江苏 南京 210037; 2. 江苏省中国科学院植物研究所(南京中山植物园) 江苏省植物资源研究与利用重点实验室, 江苏 南京 210014〕
含笑属(MicheliaLinn.)植物为木兰科(Magnoliaceae)常绿灌木或乔木,该属植物形态优美,花色多彩艳丽,表型遗传多样性丰富,是中国极具开发潜力的观赏植物,也是北亚热带地区普遍引种的园林绿化植物[1-2]。20世纪80年代初,江苏省中国科学院植物研究所(南京中山植物园)对台湾含笑〔M.compressa(Maxim.) Sarg.〕进行引种并获得了成功[3]。在含笑属植物引种过程中,低温是影响其生长和地理分布的重要非生物胁迫之一,低温胁迫对植物会造成一系列生理和生化反应,包括组织含水量降低、脯氨酸和可溶性糖等物质的积累以及激素调节[4]。近年来,国内学者对台湾含笑抗寒性开展了较多研究,陈辉等[5]研究了4种常绿阔叶树种对低温变化的生理响应,揭示了台湾含笑等植物抗寒性与可溶性糖含量等生理指标间的内在联系。亓白岩等[6-7]通过对8种含笑属植物越冬形态观察、叶片生理指标检测以及解剖结构特征分析,评价了台湾含笑与含笑属其他植物间的抗寒性强弱,发现抗寒性越强的树种,其越冬期间部分生理指标的变幅越大,解剖结构指标间比值也越大。王伟伟等[8]通过计算植株半致死温度以及观察叶片解剖结构发现,台湾含笑叶片半致死温度最低,同时叶片厚度和角质层厚度均较厚,因而抗寒性最强。
‘中山含笑’(Michelia‘Zhongshanhanxiao’)是在台湾含笑自然授粉后代的育苗过程中发现的具有明显表型变异的子代,具有生长迅速、枝叶茂密、花大量多、叶大增厚等优良特性,其观赏利用价值明显优于无明显表型变异子代[9],但目前对‘中山含笑’的抗寒性并不清楚。在低温环境下,植物叶片形态结构和生理指标变化反映了植物对逆境的适应性及自我调节能力[10-11]。依据越冬率区分植物抗寒类型是常用且相对稳定的方法,但对于同一植物不同变种间存在的细微差异难以准确区分,因此需要结合植物的形态、生理和解剖结构等指标。Meng等[12]在对樟子松(Pinussylvestrisvar.mongolicaLitv.)及其自然变种彰武赤松(Pinusdensifloravar.zhangwuensisS. J. Zhang, C. X. Li et X. Y. Yuan)的研究中发现,渗透调节物质、酶活性和丙二醛等生理指标与抗寒性存在密切的相关性,且叶片气孔密度较小的品种抗寒性更强。
鉴于此,本研究采用越冬观察法,通过测定植物叶片的可溶性糖和可溶性蛋白质含量等生理指标,并使用显微镜观察叶片解剖结构,对‘中山含笑’与台湾含笑母树及其无明显表型变异子代的抗寒性进行比较,以期确定‘中山含笑’适宜栽培区,为丰富江苏常绿阔叶树种资源以及推进城市园林建设奠定基础,并为选育抗寒性强的含笑属树种提供科学依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于江苏省南京市江宁区禄口街道苗圃地(东经118°52′24″、北纬31°41′06″),气候类型以亚热带季风气候为主,年均温17.6 ℃,极端最高温37 ℃,极端最低温-7 ℃,年降水量1 200 mm,无霜期240 d。土壤类型为黄棕壤,质地为沙壤土,酸碱度为pH 4.5~pH 5.5,有机质含量12.52 g·kg-1,全氮含量0.82 g·kg-1,全磷含量0.94 g·kg-1,全钾含量28.5 g·kg-1。3种供试植物分别为20年生的台湾含笑母树、6年生的台湾含笑无明显表型变异子代和6年生的‘中山含笑’,基本生长信息见表1。使用布鲁莱斯测高器(精度0.1 m)测量株高;使用卷尺(精度1 mm)测量地径和胸径;使用量杆(精度1 cm)测量冠层东西向和南北向的投影直径,2个投影直径平均值即为冠幅。试验期间每种植物采用同样的田间管理方式(定期灌溉除草等),以保证各植物间生长条件基本一致。

表1 3种供试植物基本生长信息
1.2 研究方法
1.2.1 越冬适应性观察 试验时间为2021年12月1日至2022年2月25日,其中,12月最低温度-5 ℃、最高温度17 ℃、平均温度6.7 ℃;翌年1月最低温度-3 ℃、最高温度13 ℃、平均温度4.7 ℃;2月最低温度-4 ℃、最高温度22 ℃、平均温度4.5 ℃,2月10日后温度回升。试验参考李瑞雪等[13]的方法对3种供试植物进行越冬观察记录,依据叶面冻害情况分为4个等级:0级,无冻害;1级,少于10%的叶片(受冻叶面积小于25%)受冻,叶面出现少量细小褐斑或形成团状褐斑;2级,10%~50%叶片(受冻叶面积小于50%)受冻,叶面色泽加深,有大面积褐斑产生或褐斑数量增多,叶缘卷曲泛黄,个别叶片呈腐烂状;3级,大于50%叶片(受冻叶面积大于50%)受冻,叶片呈深褐色,出现大面积腐烂、汁液外渗和落叶的现象。在试验地内从各供试植物中随机选出10株长势良好、无病虫害的植株,对其冠层北侧中上部的当年生枝条中部的典型越冬成熟叶片进行观察并记录叶面冻害情况,每株观察100枚叶片。
1.2.2 生理指标测定 2021年12月至2022年2月,随机从各供试植物冠层北侧中上部当年生枝条中部采摘无病虫害的第3至第5枚成熟且完整的叶片,每次取3个重复样品,每半个月左右采集1次,共采集6次。叶片采集后迅速用湿纱布包裹,装入密封塑料袋后带回实验室,依次用自来水和去离子水仔细清洗并用滤纸吸干表面水分,称量后放入液氮中速冻,置于-80 ℃冰箱中保存,用于生理指标测定。采用蒽酮比色法[14]195测定可溶性糖含量;采用考马斯亮蓝G-250染色法[14]184测定可溶性蛋白质质含量;采用酸性茚三酮显色法[14]250测定脯氨酸含量;采用氮蓝四唑法[14]167测定超氧化物歧化酶(SOD)活性;采用硫代巴比妥酸法[14]测定丙二醛(MDA)含量。生理指标数据取3次测定结果的平均值。
1.2.3 叶片解剖结构观测 2022年2月中旬从各供试植物冠层北侧中上部当年生枝条中部采摘无病虫害的第3至第5枚成熟且完整的叶片,每种供试植物取6枚叶片。从叶片中脉两侧切取面积1 cm×1 cm的小块,用FAA固定液进行固定,24 h后按常规石蜡切片法[15]制成永久切片,在Olympus BX41光学显微镜(日本Olympus公司)下观察和拍照。每种供试植物取6个切片,每个切片选取3个视野观测,并使用Digimizer 4.5.1软件测定叶片解剖结构指标,包括角质层厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度和叶片厚度,并计算栅海比(栅栏组织厚度/海绵组织厚度)、细胞结构紧密度(栅栏组织厚度/叶片厚度)、细胞结构疏松度(海绵组织厚度/叶片厚度)。叶片解剖结构数据取18个观测视野的平均值。
1.3 数据处理和分析
使用EXCEL 2016软件对数据进行统计整理,使用SPSS 16.0软件进行聚类分析和相关性分析,并采隶属函数法[16]对各指标进行综合评价。

2 结果和分析
2.1 越冬适应性分析
3种供试植物自然越冬后均有不同程度的冻害表现(图1)。统计结果显示:‘中山含笑’在自然越冬期间整体长势良好,有4%的叶片叶面出现少量细小褐斑,典型叶片叶面产生小面积团状褐斑,大部分叶片完好无损,冻害级别为1级;台湾含笑母树有9%的叶片叶面出现少量细小褐斑或形成小面积团状褐斑,典型叶片叶面产生较大面积团状褐斑,冻害级别为1级;台湾含笑无明显表型变异子代受冻叶片最多,有16%的叶片受冻,叶片表现为叶面色泽加深,叶缘卷曲泛黄,叶面褐斑数量较多,典型叶片出现腐烂症状,冻害级别为2级。3种供试植物冻害程度表现为:台湾含笑无明显表型变异子代受冻程度最重,台湾含笑母树次之,‘中山含笑’受冻程度最轻。

Z: ‘中山含笑’Michelia ‘Zhongshanhanxiao’; M: 台湾含笑母树Mother tree of M. compressa (Maxim.) Sarg.; N: 台湾含笑无明显表型变异子代Progeny without obvious phenotypic variation of M. compressa.
2.2 生理指标变化及综合评价
自然越冬下3种供试植物叶片的渗透调节物质含量的变化见表2;3种供试植物叶片的超氧化物歧化酶活性和丙二醛含量的变化见表3;基于这5个生理指标的综合评价结果见表4。
表3 自然越冬下3种供试植物叶片超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量的变化

表4 3种供试植物生理指标的综合评价
2.2.1 渗透调节物质含量的变化 结果(表2)显示:自然越冬期间(2021年12月1日至2022年2月25日),3种供试植物叶片的可溶性糖含量呈先升高后降低的变化趋势,在2022年1月15日达到峰值,其中,‘中山含笑’的峰值最高(54.78 mg·g-1),台湾含笑母树次之(53.37 mg·g-1),台湾含笑无明显表型变异子代最低(51.89 mg·g-1),但三者间差异不显著。2021年12月1日至2022年1月15日,台湾含笑无明显表型变异子代叶片可溶性糖含量的增幅在3种供试植物中最大,为59.32%。2022年2月10日至2022年2月25日,3种供试植物叶片的可溶性糖含量略有下降,但降幅不明显,依然维持在较高水平。
结果(表2)还显示:自然越冬期间,3种供试植物叶片的可溶性蛋白质含量呈先升高后降低的变化趋势,在2022年2月10日达到峰值,其中,‘中山含笑’的峰值最高(1.81 mg·g-1),台湾含笑母树次之(1.65 mg·g-1),台湾含笑无明显表型变异子代最低(1.55 mg·g-1),且‘中山含笑’与后二者间差异显著。2021年12月1日至2022年2月10日,‘中山含笑’叶片可溶性蛋白质含量的增幅在3种供试植物中最大,为141.33%。2022年2月10日至2月25日,3种供试植物叶片的可溶性蛋白质含量呈下降趋势,其中,台湾含笑母树的降幅最大,为21.21%。
结果(表2)还显示:自然越冬期间,3种供试植物叶片的脯氨酸含量呈先升高后降低的变化趋势,在2022年1月15日达到峰值,其中,‘中山含笑’的峰值最高(67.85 μg·g-1),台湾含笑母树次之(62.41 μg·g-1),台湾含笑无明显表型变异子代最低(56.40 μg·g-1),且三者间差异显著。2021年12月1日至2022年1月15日,‘中山含笑’脯氨酸含量的增幅在3种供试植物中最大,为71.82%。2022年1月15日至2022年2月25日,3种供试植物叶片的脯氨酸含量呈下降趋势,其中,‘中山含笑’的降幅最大,为8.18%。
表2 自然越冬下3种供试植物叶片渗透调节物质含量的变化
总体上看,3种供试植物叶片的可溶性糖、可溶性蛋白质和脯氨酸含量的变化趋势相似;从峰值看,‘中山含笑’上述3个指标的峰值均最高,台湾含笑无明显表型变异子代均最低。
2.2.2 超氧化物歧化酶活性和丙二醛含量的变化 结果(表3)显示:自然越冬期间,3种供试植物叶片的超氧化物歧化酶(SOD)活性总体呈先升高后降低的变化趋势,在2022年2月10日达到峰值,其中,‘中山含笑’的峰值最高(156.83 U·g-1),台湾含笑母树次之(143.71 U·g-1),台湾含笑无明显表型变异子代最低(133.47 U·g-1),且三者间差异显著。2021年12月1日至2022年2月10日,台湾含笑母树叶片SOD活性的增幅在3种供试植物中最大,为46.45%。2022年2月10至2022年2月25日,3种供试植物叶片SOD活性呈下降趋势,其中,‘中山含笑’的降幅最大,为16.72%。
结果(表3)还显示:自然越冬期间,3种供试植物叶片的丙二醛(MDA)含量呈先升高后降低的变化趋势,在2022年2月10日达到峰值,其中,台湾含笑无明显表型变异子代的峰值最高(34.74 nmol·g-1),‘中山含笑’次之(32.53 nmol·g-1),台湾含笑母树最低(30.15 nmol·g-1),且三者间差异显著。2021年12月1日至2022年2月10日,台湾含笑无明显表型变异子代叶片MDA含量的增幅在3种供试植物中最大,为119.60%。2022年2月10日至2月25日,3种供试植物叶片MDA含量呈下降趋势,其中,台湾含笑无明显表型变异子代的降幅度最大,为18.57%。
总体上看,3种供试植物叶片的SOD活性和MDA含量的变化趋势相似;从峰值看,‘中山含笑’叶片SOD活性的峰值最高,MDA含量的峰值较低;台湾含笑无明显表型变异子代叶片SOD活性的峰值最低,而MDA含量的峰值最高。
2.2.3 基于生理指标的综合评价 运用隶属函数法对3种供试植物的上述5个生理指标进行综合评价,结果(表4)显示:‘中山含笑’的平均隶属函数值最大,为0.569,其中,可溶性糖含量、脯氨酸含量和超氧化物歧化酶活性的隶属函数值在3种供试植物中均最大,分别为0.661、0.625和0.621;台湾含笑无明显表型变异子代的平均隶属函数值最小,为0.431。通过生理指标隶属函数值排序可知,中山含笑’的抗寒性最强,台湾含笑母树次之,台湾含笑无明显表型变异子代最弱。
2.3 解剖结构分析
2.3.1 叶片解剖结构特征 3种供试植物的叶片解剖结构指标和解剖图分别见表5和图2。结果显示:‘中山含笑’叶片角质层最厚(4.54 μm),台湾含笑无明显表型变异子代叶片角质层最薄(3.20 μm);‘中山含笑’叶片角质层厚度与台湾含笑母树及其无明显表型变异子代间差异显著。3种供试植物叶片上、下表皮均由一层不规则细胞紧密排列而成,且上表皮厚度均大于下表皮厚度,其中,台湾含笑母树叶片的上、下表皮最厚,‘中山含笑’叶片上、下表皮最薄;3种供试植物间的上、下表皮厚度均差异显著,变异系数分别为17.35%和18.94%。叶片栅栏组织和海绵组织厚度以‘中山含笑’最厚,台湾含笑母树次之,台湾含笑无明显表型变异子代最薄;3种供试植物间叶片栅栏组织厚度差异显著,变异系数为6.65%;‘中山含笑’叶片海绵组织厚度与台湾含笑母树无显著差异,二者与台湾含笑无明显表型变异子代差异显著,3种供试植物海绵组织厚度的变异系数为6.92%。台湾含笑母树叶片最厚(347.92 μm),台湾含笑无明显表型变异子代叶片最薄(303.63 μm),3种供试植物间的叶片厚度差异显著,变异系数为6.51%。叶片栅海比、细胞结构紧密度和细胞结构疏松度的值基本上以‘中山含笑’最大,台湾含笑无明显表型变异子代次之,台湾含笑母树最小;3种供试植物间的栅海比无显著差异,细胞结构紧密度差异显著;细胞结构疏松度表现为‘中山含笑’与台湾含笑无明显表型变异子代无显著差异,二者与台湾含笑母树差异显著。
表5 3种供试植物叶片解剖结构指标的比较

cu: 角质层Cuticle; ue: 上表皮Upper epidermis; pp: 栅栏组织Palisade parenchyma; sp: 海绵组织Spongy parenchyma; le: 下表皮Lower epidemis; vb: 维管束Vascular bundle.
2.3.2 叶片解剖结构指标筛选 采用分层聚类的方法对上述9个叶片解剖结构指标进行分析,结果(图3)显示:在欧氏距离5处,9个指标分为3类:Ⅰ类包括细胞结构紧密度、细胞结构疏松度、栅海比、角质层厚度、下表皮厚度和上表皮厚度;Ⅱ类包括栅栏组织厚度和海绵组织厚度;Ⅲ类仅包括叶片厚度。
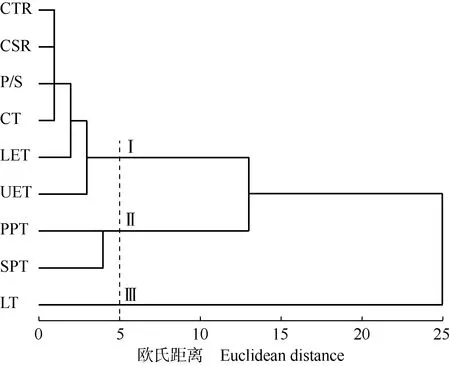
CTR: 细胞结构紧密度Cell tense ratio; CSR: 细胞结构疏松度Cell spongy ratio; P/S: 栅海比Palisade parenchyma and spongy parenchyma ratio; CT: 角质层厚度Cuticle thickness; LET: 下表皮厚度Lower epidermis thickness; UET: 上表皮厚度Upper epidermis thickness; PPT: 栅栏组织厚度Palisade parenchyma thickness; SPT: 海绵组织厚度Spongy parenchyma thickness; LT: 叶片厚度Leaf thickness.
进一步对各类中指标间的相关性进行分析,结果(表6)显示:细胞结构紧密度与角质层厚度和栅海比呈极显著正相关,与上、下表皮厚度呈极显著负相关;细胞结构疏松度与上表皮厚度和栅海比分别呈显著和极显著负相关,与下表皮厚度呈显著正相关;角质层厚度与上表皮厚度和下表皮厚度分别呈显著和极显著负相关;上表皮厚度与下表皮厚度呈极显著正相关。栅栏组织厚度和海绵组织厚度呈极显著正相关。

表6 3种供试植物叶片解剖结构指标间的相关性分析1)
根据各解剖结构指标间的相关性大小计算3类指标的相关指数。结果(表7)显示:Ⅰ类中细胞结构紧密度的相关指数最大,为0.335,选择为该类的典型指标;Ⅱ类中栅栏组织厚度和海绵组织厚度的相关指数相同,且海绵组织厚度和栅栏组织厚度的变异系数相差不大,因此选择更能反映叶肉组织结构特点的栅栏组织厚度作为该类的典型指标;Ⅲ类中仅叶片厚度1个指标,选择为该类的典型指标。
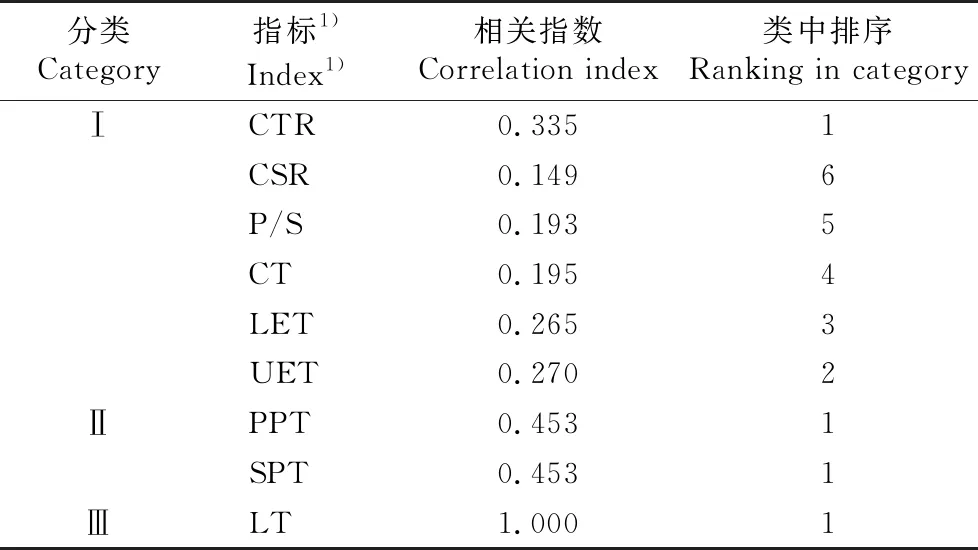
表7 3种供试植物叶片解剖结构指标的相关指数及排序
2.3.3 基于叶片解剖结构指标的综合评价 运用隶属函数法对3种供试植物的3个叶片典型解剖结构指标进行综合评价,结果(表8)显示:‘中山含笑’的平均隶属函数值最大,为0.686,其中,细胞结构紧密度和栅栏组织厚度的隶属函数值在3种供试植物中均最大,分别为0.679和0.770;台湾含笑无明显表型变异子代的平均隶属函数值最小,为0.267。通过叶片典型解剖结构指标隶属函数值排序可知,中山含笑’的抗寒性最强,台湾含笑母树次之,台湾含笑无明显表型变异子代最弱。

表8 3个供试植物叶片典型解剖结构指标的综合评价
3 讨论和结论
植物越冬适应性观察法是通过观察植株叶片受冻组织和未受冻组织的颜色,进而确定植物受冻害程度,能够很好地反映出植物在越冬期间的冻害情况[17]。高驰等[18]通过对7种重楼属(ParisLinn.)植物越冬观察发现,抗寒能力不同的树种叶片有不同的外观表现。本研究中,与同龄的台湾含笑无明显表型变异子代相比,在自然越冬期间(2021年12月1日至2022年2月25日)的各个阶段,‘中山含笑’均表现出了更强的抗寒性,未出现叶缘卷曲泛黄、腐烂的现象,仅少数叶片受冻产生褐斑。‘中山含笑’作为台湾含笑自然变异种,其不仅长势明显高于同龄的台湾含笑无明显表型变异子代,且多数生理指标和叶片解剖结构指标的隶属函数值也最大,而台湾含笑无明显表型变异子代的多数生理指标和叶片解剖结构指标的隶属函数值最小。因此,综合3种供试植物的越冬表现以及各指标隶属函数分析结果可知:‘中山含笑’的抗寒性最强,台湾含笑母树次之,台湾含笑无明显表型变异子代最弱。
植物遭受低温胁迫会产生一定的适应能力,而抗寒性强的植物在面对低温冻害时具有强大的生理适应机制,可使自身的发育和生理代谢能够正常运转[19]。研究结果表明:可溶性糖、可溶性蛋白质和脯氨酸是植物细胞内重要的渗透调节物质,能够促进细胞吸水和保水,其含量与植物的抗性呈正相关[20-21]。本研究中,自然越冬期间,3种供试植物叶片的渗透调节物质(可溶性糖、可溶性蛋白质和脯氨酸)含量均随着时间的推移呈先升高后降低的变化趋势,Pociecha等[22]认为这是一种普遍现象,说明3种供试植物在低温胁迫下可以通过增加可溶性糖等渗透调节物质的含量来调节细胞内电解质渗透压,防止细胞脱水。同时可溶性蛋白质含量的增加有助于合成逆境响应蛋白及保护性酶,这与曾光辉等[23]在茶树〔Camelliasinensis(Linn.) O. Ktze.〕上的研究结果是一致的。抗寒性强的树种遭遇低温胁迫时渗透调节物质含量要高于抗寒性弱的树种,且增幅也较大[24]。本研究中,‘中山含笑’叶片的渗透调节物质含量峰值均高于台湾含笑母树及其无明显表型变异子代,且增幅总体也较大,验证了‘中山含笑’抗寒性最强。王伟伟等[8]研究发现,含笑属植物的半致死温度在-16 ℃~-11 ℃,而本试验中样品采集地的最低温度仅为-5 ℃,在2022年2月10日后随着自然平均温度升高,3种供试植物叶片的渗透调节物质含量均出现了一定程度的下降,说明-5 ℃的低温胁迫未能对植物细胞膜造成不可逆转的损伤。
Campos等[11]研究认为,丙二醛含量的变化可以反映植物细胞膜的受损程度和逆境胁迫抵御能力。本研究中,自然越冬期间,3种供试植物叶片的丙二醛含量均随着时间的推移呈先升高后降低的变化趋势,这与马娟娟等[16]对北美冬青〔Ilexverticillata(Linn.) A. Gray〕的低温胁迫试验结果一致,说明3种供试植物细胞膜功能均受到了抑制。从12月份到翌年2月份,‘中山含笑’和台湾含笑母树叶片的丙二醛含量积累较慢,而台湾含笑无明显表型变异子代丙二醛含量迅速增加,其增幅最大且峰值最高,表明其活性氧自由基积累最多,细胞膜受损程度最大,抗寒性最差。植物在正常状态下时,体内活性氧自由基的生成与清除处于动态平衡,低温胁迫能够打破植物体内活性氧自由基的动态平衡,而植物为适应低温环境会提高超氧化物歧化酶活性来迅速清除体内积累的活性氧自由基,从而对自身进行保护[25-26]。本研究结果显示:3种供试植物叶片的超氧化物歧化酶活性随着时间的推移总体呈先升高后降低的变化趋势,且在2022年2月10日达到峰值,说明低温胁迫激发了植物体内抗氧化酶系统,对体内产生的活性氧自由基进行调节以抵御低温胁迫[27]。Airaki等[28]在对五彩椒(CapsicumannuumLinn.)的研究中发现,抗氧化酶活性在一定范围内与植物抗寒性呈正相关。本研究中,‘中山含笑’的超氧化物歧化酶活性在整个越冬期间均高于台湾含笑母树及其无明显表型变异子代,即抗寒性强的植物保护酶活性相对较高。
叶片在植物生长发育过程中会形成与当地环境相适应的外部形态和解剖结构[29],植物抗寒性与叶片解剖结构存在密切关系[7,30-31]。李瑞雪等[13]认为,含笑属植物叶片栅栏组织越厚,栅海比越大,抗寒性越强。本研究通过聚类分析和相关性分析,从9个解剖结构指标中筛选出细胞结构紧密度、栅栏组织厚度和叶片厚度作为3个典型指标,以消除冗余信息的影响。细胞结构紧密度为栅栏组织厚度与叶片厚度的比值,栅栏组织为薄壁细胞,有利于各种物质的进出,栅栏组织越厚,细胞结构紧密度越大,表现出的抗寒性也越强[32]。本研究发现,3种供试植物叶片解剖结构中,‘中山含笑’细胞结构紧密度和栅栏组织厚度均大于台湾含笑母树及其无明显表型变异子代,与抗寒性评价一致。谭殷殷等[33]研究发现,叶片厚度可以作为抗寒性鉴定的典型指标。本研究中叶片厚度虽然也作为反映抗寒性的主要指标之一,但3种供试植物的叶片厚度以台湾含笑母树最厚,这与其抗寒性评价结论略有不同,这可能与本研究中叶片厚度变异系数较小有关。‘中山含笑’具有枝叶茂密、花大量多、叶大增厚等优良特性,其角质层厚度、海绵组织厚度、栅海比和细胞结构疏松度均大于台湾含笑母树。但‘中山含笑’上、下表皮在3种供试植物中最薄,与其抗寒性评价结果相反,这可能是由于含笑属植物抗寒性不仅与上、下表皮有关,还与表皮上的气孔密度有关[34]。
综上所述,自然越冬期间3种供试植物均出现了一定程度的冻害,其中台湾含笑无明显表型变异子代受冻程度最重,‘中山含笑’受冻程度最轻。‘中山含笑’除丙二醛含量外的其余生理指标的峰值均为3种供试植物中最高,台湾含笑无明显表型变异子代叶片丙二醛含量的增幅最大且峰值最高,台湾含笑母树各生理指标介于二者之间。3种供试植物中,‘中山含笑’细胞结构紧密度和栅栏组织厚度均最大,台湾含笑无明显表型变异子代的栅栏组织厚度和叶片厚度均最小。综合自然越冬表现及各指标隶属函数分析结果可知:‘中山含笑’的抗寒性最强,台湾含笑母树次之,台湾含笑无明显表型变异子代最弱。低温对植物的影响是一个复杂的过程,植物的耐寒性是受多因子调控的,后续将对植物自然越冬期间内源激素变化和抗寒相关基因表达开展进一步的研究。
