濒危植物合柱金莲木开花结实物候及交配系统研究*
2023-03-01陈宗游柴胜丰汪俊芳
陈宗游,韦 霄,柴胜丰,邹 蓉,汪俊芳
(1.广西壮族自治区中国科学院广西植物研究所,广西植物功能物质与资源持续利用重点实验室,广西桂林 541006;2.四川大学生命科学学院,四川成都 610065;3.广西壮族自治区林业勘测设计院,广西南宁 530022)
合柱金莲木(Sauvagesiarhodoleuca)又名辛木,属金莲木科(Ochnaceae)合柱金莲木属(Sauvagesia)小灌木,为我国特有的单种属植物。合柱金莲木分布区域狭窄、分布零星,仅在广东和广西局部山区海拔200-1 000 m的中低山发现有少量分布[1,2]。合柱金莲木具有一定的药用价值,其根茎可以入药,具有止痒杀虫之功效[3]。因分类地位特殊,合柱金莲木对研究金莲木科植物的区系、地理分布及其发生与演化等具有重要的价值[1,4]。受生存环境恶化及森林砍伐、采根入药等人类活动的影响,合柱金莲木分布范围和数量日益减少,在不少分布区合柱金莲木濒临灭绝或已绝迹。在广西,龙胜[5]、象州[6]两县原记载有合柱金莲木分布,但经广西壮族自治区中国科学院广西植物研究所(以下简称广西植物研究所)科研人员实地调查,未发现该物种;在广东,合柱金莲木原见于怀集、连山、封开、广宁、海陵岛等地,但广东省林业局于1999-2001年组织有关专家对该省国家重点保护野生植物资源进行调查时,仅在肇庆的广东封开黑石顶省级自然保护区石门塘发现有合柱金莲木分布[7]。然而在2007年,笔者在开展合柱金莲木遗传多样性研究的野外采样时,发现连山、封开、广宁尚有合柱金莲木存在,只是其种群个体数量较少,其中广宁采样点只采到9个样[8]。但笔者在2013年进行合柱金莲木的伴生群落调查时[2],在广宁原采样点已找不到合柱金莲木植株。目前在《中国植物红皮书(第一册)》[9]和《国家重点保护野生植物名录》[10]中,合柱金莲木分别被列为二类保护植物和国家二级重点保护野生植物。
微卫星又称简单串联重复序列(Simple Sequence Repeat,SSR)或短串联重复序列(Short Tandem Repeat,STR),是以1-6 bp的碱基重复单元组成的串联重复序列。因具有数量多、丰富度高、易于检测等特点,微卫星被广泛用作遗传差异的标记[11]。目前已有不少基于微卫星技术的交配系统的研究,如罗芊芊等[12]运用12对微卫星引物对2个南方红豆杉(Taxuswallichianavar.mairei)天然居群的交配系统进行分析,结果发现南方红豆杉属于高度异交树种;Liesebach等[13]基于微卫星对挪威枫(Acerplatanoides)种子园亲本无性系及其种子样本进行基因分型检测,结果发现挪威枫有68%的异交后代、11%的自交后代和20%的外部授粉后代。
植物的开花结实物候及交配系统属于繁殖生物学范畴,是植物在自然状态下对环境长期适应而演化出的主要生殖特征。对于大多数濒危植物来说,在繁殖过程中通常会观察到生殖障碍[14],而生殖障碍是导致部分植物濒危的重要原因[15-18]。现有文献中有关合柱金莲木的研究报道十分有限,仅见有关形态[4]、生境条件[1]、药用价值描述[3],以及种子萌发特性[19]、扦插繁殖[20]和伴生群落特征[2]的研究,而繁殖生物学方面的开花结实物候及交配系统研究迄今未见报道。鉴于微卫星是分析交配系统较好的分子手段,本研究开展合柱金莲木的开花结实物候观测并利用微卫星分子标记测定其交配系统,拟从繁殖生物学角度探讨其濒危机制,为其保护和可持续利用提供科学依据。
1 材料与方法
1.1 分布点自然概况
合柱金莲木分布区位于南亚热带北缘及中亚热带南缘。分布区内雨量多、湿度大,年平均气温18-20 ℃,最冷月平均气温9-11 ℃,最热月平均气温约28 ℃,全年无霜期250-310 d,年降水量1 400-1 600 mm,相对湿度为75%-85%。土壤为山地红壤和谷底冲积土,成土母岩以花岗岩、页岩、砂岩为主,土层厚度0.6-1.2 m,腐殖质层厚5-30 cm[2]。合柱金莲木为林下阴生植物,不耐强光和干旱,常生于土壤湿润且郁闭度较高的常绿阔叶林中。目前经过调查确认广西融水(S1)、金秀(S2)、德保(S3)和广东封开(S4)、连山(S5) 5个县存在小块区域的合柱金莲木种群,各分布点概况见表1和图1。
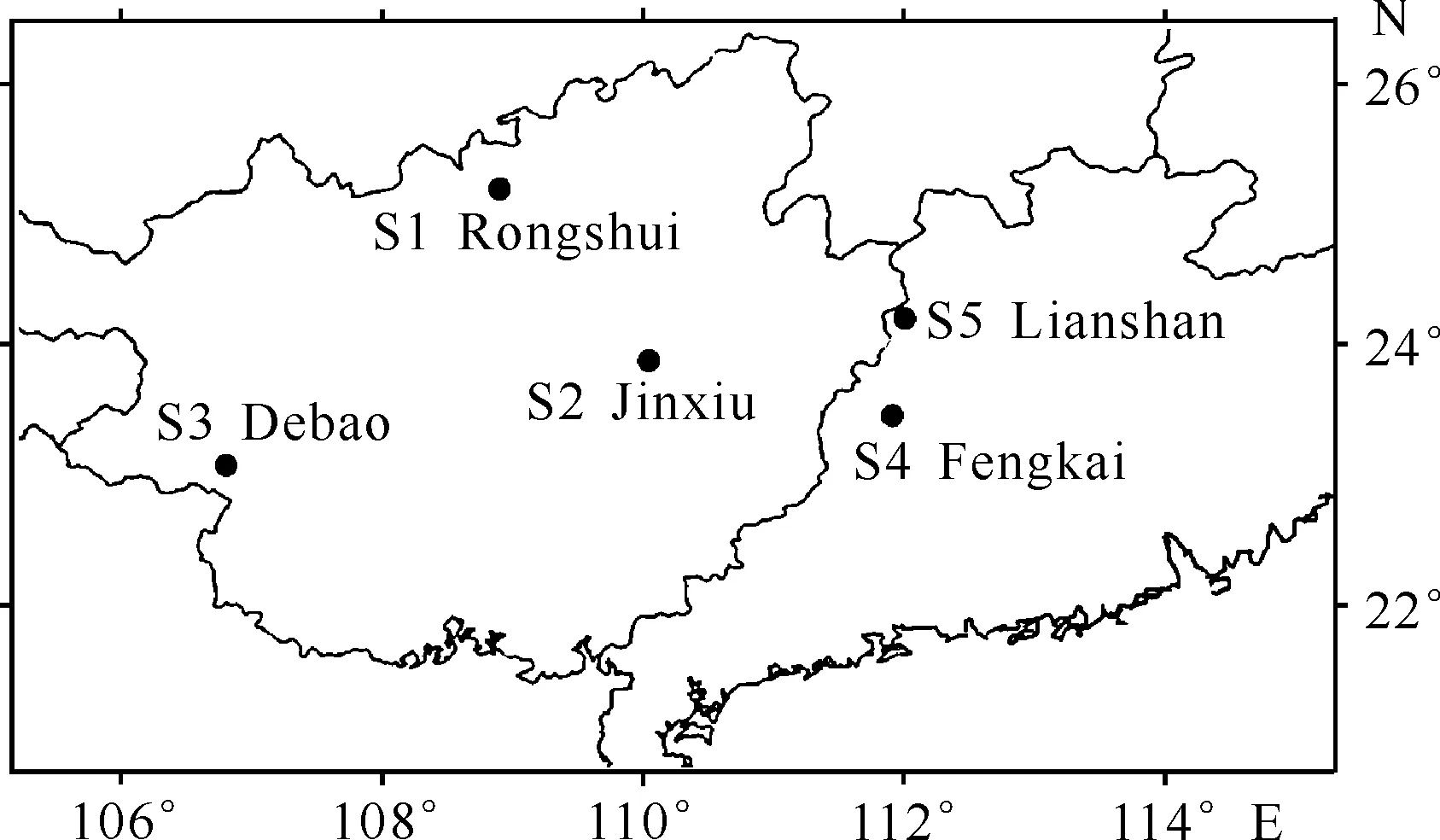
图1 合柱金莲木野外分布点
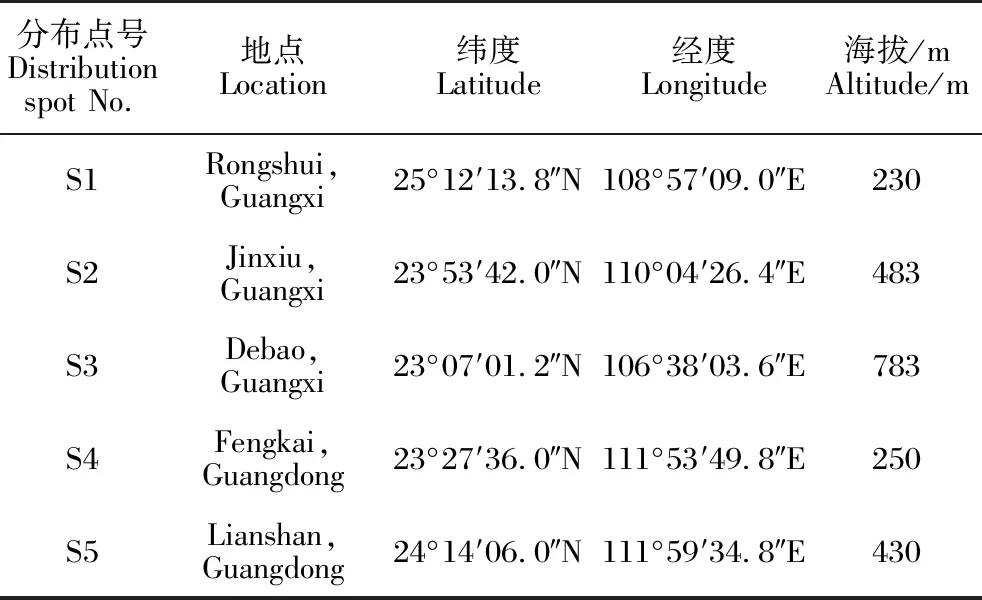
表1 合柱金莲木各分布点的基本概况
1.2 材料及试验地点
植株材料经广西植物研究所蒋运生研究员鉴定为合柱金莲木(Sauvagesiarhodoleaca)(图2)。本研究选择S2和S3两个合柱金莲木分布点作为野外试验点,成龄植株的基本形态特征及其开花结实物候期观测在上述两处试验点进行;单花开花动态、生殖构件特征的观测在S2分布点进行。交配系统研究在广西植物研究所实验室进行,样品(叶片和种子)分别采自S1、S2、S3、S4和S5 5个合柱金莲木分布点,引物为Li等[8]开发的9对合柱金莲木微卫星引物(SSR1、SSR3-SSR11),将其送到上海英骏生物技术有限公司合成荧光引物,采用羧基荧光素(Carboxyfluorescein,FAM)和六氯荧光素(Hexachloro-fluorescein,HEX )标记不同引物的正向引物。

1:fruit branch;2:enlargement of leaf;3:sepal;4:petal;5:external whorl of staminodes;6:second whorl of staminodes;7:inner whorl of staminodes;8:stamen;9:pistil;10:fruit;11:seed.
1.3 方法
1.3.1 成龄植株的基本形态特征及开花结实物候期观测
本研究将已花合柱金莲木视为成龄植株。在花蕾期,于观测点随机标记成龄合柱金莲木30株,统计丛生植株的比例,并测定各植株的地径(丛生植株测定其最大分枝)、株高、冠幅,统计每株植株的花序数。此外,初步观测和判断合柱金莲木的生育期。根据生育期,于次年3-10月以上年标记的植株为对象进行开花结实物候期观测,每5 d观察1次。结合Dafni[21]和柴胜丰等[22]的研究确定花期观察标准:约5%的植株抽出花序的时期为抽(花)序期;平均单株开花数约达5%的时期为始花期,约达50%为盛花期,大于90%为末花期。
1.3.2 开花动态观测
在合柱金莲木花蕾期,于观测点随机选取15株植株上的30串未开放的花序进行标记,观察花序单花的开放顺序和开花天数;随机选取成龄合柱金莲木5株,每株植株选定5朵刚现的花蕾作为单花开放动态的观测对象,每2-3 d观察1次,直至花朵开放。花朵开放当天,每隔10-15 min观测1次。此后,每天观测1次,直至花朵脱落。
1.3.3 生殖构件特征的观测
在合柱金莲木盛花期,随机选定花序30串,对花序柄长、花序轴长、花序宽(花序最宽处的值)及花序的单花数进行统计。采集成熟花蕾30朵,统计其苞长和苞宽(花苞最宽处的值)。采集盛开单花30朵,统计其花冠直径、花瓣数、花瓣长度和宽度(花瓣最宽处的值)、萼片数、萼片长度和宽度(萼片最宽处的值)、花柄长度及直径、子房直径等;在果期,随机选定成熟果序30串,对果序柄基径、果序柄长、果序轴长、果序宽(果序最宽处的值)及果序的蒴果数等进行统计。采集成熟蒴果30粒,统计果球纵横径、果柄长度及直径、蒴果的种子数、种子纵横径及种子千粒重等。以《中国植物志(第四十九卷第二分册)》[6]和《广西植物志(第一卷)》[5]有关合柱金莲木形态学特征的描述为参考,结合观测结果对合柱金莲木的生殖构件特征进行详细描述。
1.3.4 交配系统检测
于合柱金莲木种子成熟期,对S1、S2、S3、S4和S5 5个分布点进行样品采集:每个分布点选30株成龄植株采集鲜叶片和种子,每一单株及其种子组成一个家系(Family),采样时,对各家系的母株及种子进行标记和编号。单株鲜叶采集后迅速装入封口袋,加硅胶进行干燥;种子带回实验室用于培养幼苗,所培养的幼苗及其对应的母株叶片用于DNA提取。
采用十六烷基三甲基溴化铵法(Cetyltrimethylammonium Bromide,CTAB)[23]提取DNA,基于微卫星分子标记试验对合柱金莲木进行交配系统检测,利用9对SSR引物对所培养出的合柱金莲木幼苗及其对应母株的DNA进行荧光PCR扩增。PCR反应体系为15 μL,包括1×Buffer,2.0 mmol/L MgCl2,4种dNTP各0.3 mmol/L,正、反向引物各0.75 μmol/L,0.75 U Taq酶,10 ng模版DNA。扩增程序:95 ℃预变性5 min;94 ℃变性30 s,53 ℃退火30 s,72 ℃延伸45 s,进行35个循环;最后72 ℃延伸10 min。为节省经费,笔者把不同荧光标记引物混合在一起进行PCR扩增,或者当两个以上(含两个)引物PCR扩增产物长度片段相差至少50 bp时,同样荧光标记的引物也放在一起进行PCR扩增,接着将两种方式的扩增产物在3037xl DNA分析仪(美国赛默飞世尔科技公司)上进行毛细管电泳,最后采用Genemarker 2.2.0软件分析目标片段大小。
1.3.5 参数处理与数据分析
1.3.5.1 基本形态特征参数
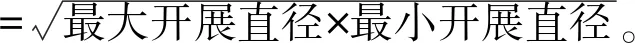
采用SPSS 13.0软件对S2和S3两个分布点的合柱金莲木成龄植株的基本形态指标进行t检验,显著性水平设为0.05。描述性统计值用平均值±标准误(Mean±S.E.)、平均值(Mean)表示。
1.3.5.2 交配系统参数的计算
用GenALEx 6软件[24]计算合柱金莲木的近交系数理论推算值(F1)以衡量其是否偏离Hardy-Weinbery平衡,并根据F1计算理论预期异交率(te),te=(1-F1)/(1+F1)[25];用MLTR 3.4统计软件[26,27]计算合柱金莲木母代和子代的多位点异交率(tm)和单位点异交率(ts)、双亲近交系数(tm-ts)、多位点相关度(rpm)、单位点相关度(rps)、异交率相关性(rt)、亲本近交系数当代估算值(F)以及亲本相关性(rp);根据Ritland[28]和赵耀等[29]的估算方法,使用tm和F值估计近交衰退系数(δ),δ=1-2Ftm/(1-tm)(1-F)。描述性统计值用平均值±标准差(Mean±S.D.)、平均值(Mean)表示。
2 结果与分析
2.1 成龄植株的基本形态特征及其开花结实物候期
从表2可知,成龄合柱金莲木植株的平均地径、平均株高和平均冠幅分别为0.70、129.67和81.98 cm,平均每株结花序15.56串,有一定比例的成龄植株为丛生植株。与S3分布点相比,S2分布点的成龄植株更大(地径、株高和冠幅的数值更大)、平均每株花序数更多,但差异不显著(P>0.05)。而S2分布点的平均每丛枝数显著大于S3(P<0.05)。

表2 不同分布点合柱金莲木成龄植株的基本形态特征
成龄合柱金莲木在3月下旬至4月中上旬进入抽(花)序期,4月中下旬进入现蕾期,5月中上旬为现蕾末期,4月下旬至5月上旬进入始花期,5月中上旬达到盛花期,5月中下旬为末花期,花期持续1个月左右。合柱金莲木于5月下旬至6月上旬进入果实发育期,经过60-70 d的生长,果实进入成熟期。不同的分布点,合柱金莲木的开花结实物候期存在一些差异,S3分布点的物候期比S2分布点提前10 d左右(表3)。

表3 合柱金莲木开花结实物候期
2.2 开花动态
合柱金莲木的花序单生于枝条上部的叶腋。进入抽序期后,随着花序轴的生长,其基部一般会产生侧枝,中上部及基部的侧枝不断离心产生丛生花蕾。合柱金莲木花蕾的开放顺序一般为花序下部的花蕾先开放,然后往上逐渐开放,偶见花序上部的花蕾先于下部花蕾开放的现象。合柱金莲木的单花开放过程可分为蕾松、初开、盛开和落花4个阶段。从现蕾开始,单花经过15-20 d的孕蕾期(图3:1-5)后膨胀到了极限,原来紧紧包裹的花瓣开始逐渐松开,花朵进入蕾松期(图3:6);花瓣继续松动,花蕾顶端裂开出现“小口”,此时花瓣与雌蕊之间的夹角呈10°左右,花瓣内拱,从“小口”可见退化雄蕊中轮的中上端,此为初开阶段(图3:7);花朵继续开放,花瓣向外伸展,与雌蕊的夹角呈70°左右,并呈碗状内拱,退化雄蕊中轮及外轮完全展现出来,此为盛开阶段(图3:8);花瓣外翻、脱落,部分单花开始凋落,部分单花转入子房膨大期,此为落花阶段(图3:9-11)。单花开放1 d内完成,从蕾松阶段到初开阶段约10 min,从初开阶段到盛开阶段约30 min,从盛开阶段到脱落阶段约5 h。脱落的单花从花瓣外翻到整个单花彻底脱落需要1-2 d,而不脱落的转入子房膨大期的单花(图3:12),其子房迅速膨胀,果实迅速生长,至花瓣外翻后的第15-20天,蒴果基本成型,花萼附在子房上相当长的时间内不萎蔫凋落(图4)。

1-5:budding stage; 6:bud loosening stage; 7:early flowering; 8:flowering stage; 9-11:wilt stage; 12:ovary growth stage.

图4 合柱金莲木单花子房膨胀进程
2.3 生殖构件特征
合柱金莲木的花序呈狭圆锥状,花序轴长约6.28 cm,花序宽约3.35 cm,花序柄长约2.91 cm,每串花序约有27朵单花;合柱金莲木的幼蕾呈圆锥形,成熟花蕾呈长圆锥形,苞长约7.61 mm,苞宽约3.12 mm;盛开单花的花冠直径约1.61 cm,花柄长约1.06 cm,花柄基径约0.21 mm。单花有萼片5枚,浅青色,呈卵形或披针形,长约4.49 mm,宽约1.78 mm,边缘有柔毛且呈锯齿状(图5:1);花瓣5枚,椭圆形,长约8.57 mm,宽约3.13 mm,其上部为白色,基部为橙黄色,白色部分占花瓣面积的5/6左右(图5:2)。盛开单花显示萼片与花瓣交互轮生(图3:8,9)。退化雄蕊3轮,外轮基部连合生成一短管,上部分离呈腺体状,高约2.21 mm,中轮和内轮各5枚,均为花瓣状,长椭圆形,上部为白色,基部为橙黄色,白色部分占中轮和内轮面积的5/6左右,中轮略大于内轮,中轮的高度和宽度分别为5.92 mm和1.18 mm,内轮的高度和宽度分别为5.53 mm和0.97 mm(图5:3-5)。合柱金莲木的单花同时具有雌蕊和雄蕊(图2:8,9;图5:6,7),为两性花。发育雄蕊5枚,花丝极短,花药长2.05 mm,宽0.91 mm(图5:6);子房卵形,纵径约1.86 mm,横径约0.91 mm,花柱长约5.25 mm,呈尾状渐尖,柱头小(图5:7)。发育雄蕊、子房及花柱被退化雄蕊的中轮和内轮所包裹,只有柱头尖端露出(图3:8-11)。
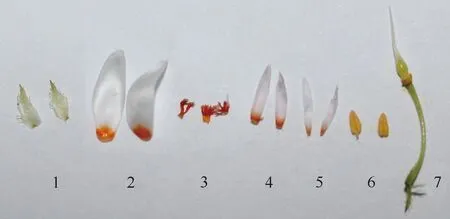
1:sepals;2:petals;3:external whorl of staminodes;4:second whorl of staminodes;5:inner whorl of staminodes;6:stamens (filaments,anthers);7:pistil,ovary,flower stalk.
合柱金莲木的果序轴长约8.75 cm,果序宽约5.08 cm,果序柄基径约0.83 mm,果序柄长约4.66 cm,果序轴上的分穗(丛)数(总轴上的分枝数与丛生蒴果丛数之和)约11穗(丛),每串果序约有18个蒴果;蒴果卵球形,先端具宿存的细长花柱,果皮上有3道凹槽,成熟蒴果开裂时,从其先端顺凹槽下裂,先端形成嘴状裂口,种子可从裂口掉出。蒴果果球纵径约4.94 mm,横径约4.36 mm,蒴果千粒重(干重)约18.24 g,单果种子数约21粒;种子椭圆形,纵径约1.52 mm,横径约0.78 mm。种皮硬而脆,呈暗红色。种子千粒重(干重)约0.61 g。
按照每株15.56串花序(果序)(表2),每串花序(果序)18个蒴果,每个蒴果21粒种子,则平均每株成龄合柱金莲木的种子数为15.56×18×21=5 881.68粒。
2.4 交配系统分析
2.4.1 荧光引物PCR的检测结果
采集用于交配系统研究的合柱金莲木种子,最终培养到35株幼苗,对应17株母株(即17个家系),所有家系的成员合计52个个体,9对荧光引物在52个个体上共检测到140个等位基因,每个等位基因的片段大小为107-345 bp不等。下图为其中一个合柱金莲木DNA样本分别在两对荧光引物(荧光素分别为FAM和HEX)上扩增的结果(图6),图6(a)和6(b)相对应的基因座分别为杂合子和纯合子基因型。
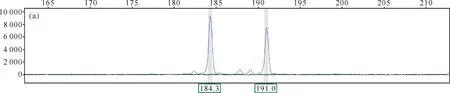

Numerical values in the vertical axis represent fluorescence intensities,and those in the horizontal axis represent the size of the amplified fragment.The appearance of two distinct peaks in (a) indicates that the locus is heterozygous,while the appearance of a single distinct peak in (b) indicates that the locus is homozygous.The horizontal axis values corresponding to each peak in the figure represent the size of their amplified fragments.
2.4.2 交配系统
经过培养得到的合柱金莲木幼苗对应的17个家系分别属于5个采样点,每个采样点的家系数目3-4个。鉴于每个采样点的家系数目太少,本研究将17个家系作为整体来分析合柱金莲木的交配系统,其结果见表4。
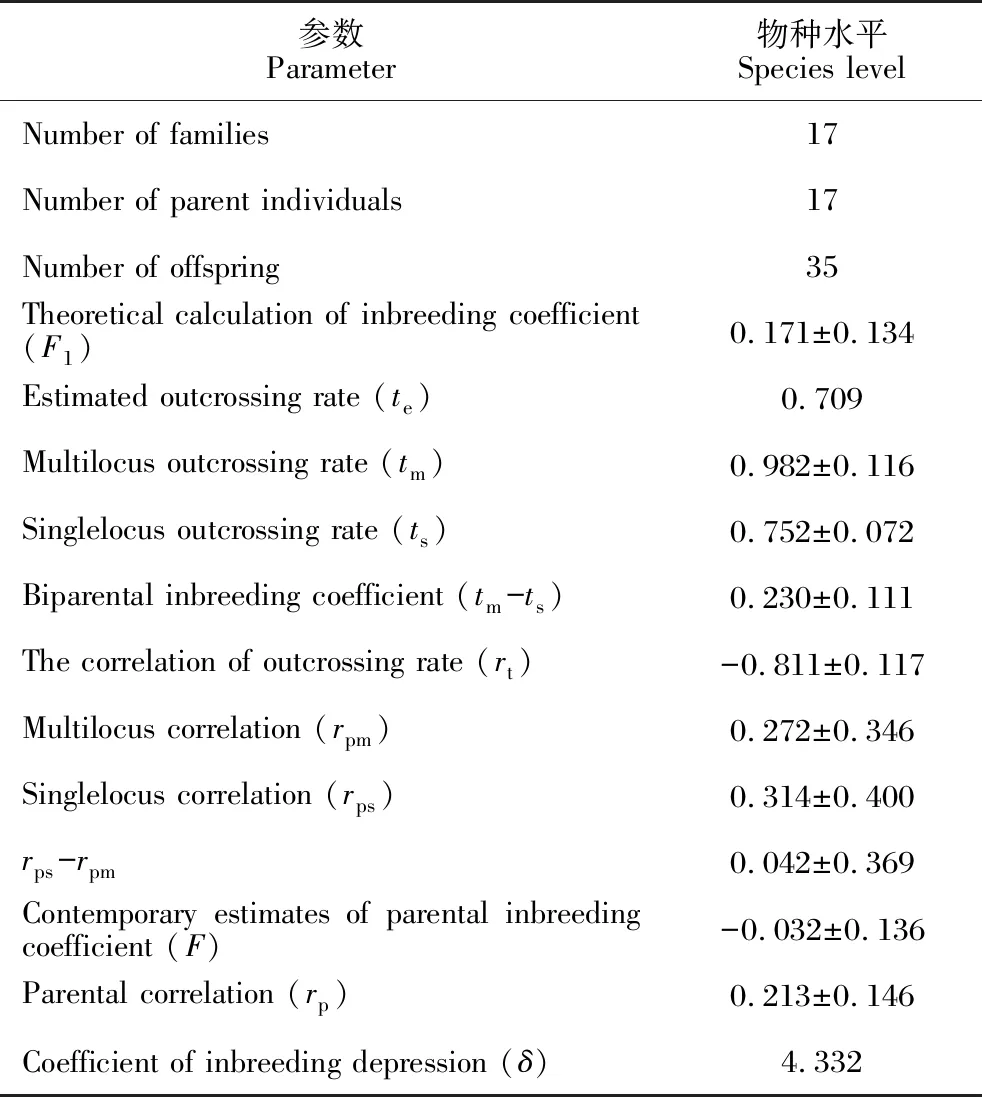
表4 合柱金莲木的交配系统参数
从近交系数理论推算值(F1=0.171)来看,合柱金莲木在理论上偏离了Hardy-Weinbery平衡,在物种水平上有过剩的纯合子;而从亲本近交系数当代估算值(F=-0.032)来看,合柱金莲木在物种水平上接近随机交配,但从该估算值的标准差(0.136,该数值比较大)来看,可知该物种的部分种群偏离Hardy-Weinbery平衡,部分种群存在近亲交配。
合柱金莲木的预期异交率(te)、多位点异交率(tm)和单位点异交率(ts)分别为0.709、0.982和0.752。tm与ts的差值反映双亲近交程度,合柱金莲木多位点异交率高于单位点异交率,即tm-ts=0.230>0,说明在物种水平上合柱金莲木存在一定比例的近交(双亲的近交比例为0.230)。
合柱金莲木物种水平上的多位点相关度(rpm)和单位点相关度(rps)分别为0.272和0.314,其单位点相关度大于多位点相关度(rps-rpm=0.042>0),说明物种内部存在明显分化的种群遗传结构。异交子代间亲本相关性(rp)为0.213,表明部分子代共享了亲本。近交衰退系数(δ)大于0,说明合柱金莲木在物种水平上存在近交衰退。
3 讨论
合柱金莲木的花为圆锥花序,平均每株成龄植株的花序数量为15串左右,花序单花数平均达27朵。合柱金莲木的单花开放在1 d内完成,但因单花开放不同步,其花期长达1个多月。这种集成花序结构的花,开花持续时间长且不同步,一方面保证植株在较长的时间内能成功完成异花传粉受精过程,从而保证生殖成功;另一方面也可降低花期恶劣的自然环境对其生殖成功造成的不良影响,这是部分植物长期适应环境影响所形成的一种生殖策略[30]。从种子量来看,合柱金莲木的花采取的集成花序结构策略是成功的,合柱金莲木的平均每串果序的蒴果数约18个,每个蒴果的种子数约21粒,平均每株的种子数达5 881.68粒,种子数量极大。
合柱金莲木的发育雄蕊与雌蕊异长,花药远低于花柱,它们被包裹于退化雄蕊的中内轮当中,只有柱头露出(图3:8-11)。一个值得注意的问题是,笔者在对单花开放过程的观测中,未发现任何有退化雄蕊中内轮展开的单花,但偶见有蜂类访花,可未见访花者长有可插入退化雄蕊的中内轮当中的长喙,被访过的单花的退化雄蕊的中内轮亦未见有被咬破或咬开的情况。由于花药被包裹于退化雄蕊的中内轮当中,退化雄蕊的中内轮不展开且不被昆虫咬破或咬开,其花粉应是难以散发出来的,而其柱头露在退化雄蕊中内轮外面,花粉如何传授到柱头,这是一个目前未弄明白的问题。因其单花太小,笔者做两次套袋试验均未成功,通过一些传统的方法来确定其交配类型比较困难。由于合柱金莲木退化雄蕊中内轮将花药包裹得较为严实,风应难以将花粉吹散出来。因此,合柱金莲木是否存在不经授粉的无融合生殖或自花授粉(可能在雌蕊柱头伸出退化雄蕊中内轮之前已完成授粉)或其他笔者未观测到的异花授粉行为,还需进一步深入研究。
本研究结果显示,合柱金莲木在理论上偏离了Hardy-Weinbery平衡,在物种水平上有过剩的纯合子,部分种群存在近亲交配。这一研究结果支持了Li等[8]关于合柱金莲木种群存在近亲交配的研究结论。本研究显示合柱金莲木物种水平上的近交系数理论推算值为0.171,小于Li等[8]关于合柱金莲木各种群的近交系数理论推算值为0.464-0.950的研究结果。一般来讲,物种水平上的近交系数比种群水平上的低属正常;另外,由于Li等[8]的研究是采用变性聚丙烯酰胺凝胶电泳法结合人工读带对PCR产物进行相关分析,而本研究是采用毛细管电泳法结合Genemarker 2.2.0软件来分析目标片段大小,人工读带可能出现偏差从而使两个研究结果存在差异。
交配系统是物种的基本属性,交配系统中的自交(或近交)程度受很多因素的影响,而这些因素与物种种群的大小密切相关。小种群因交配对象少,导致自交和近交概率明显增加[31,32],多种小种群物种如蒜头果(Malaniaoleifera)[33]、细叶杨(Populusilicifolia)[34]、龙棕(Trachycarpusnanus)[35]存在自交或近交现象。合柱金莲木现有的几个分布点的种群规模都十分小,各分布点的个体数量十分有限。目前尚不清楚合柱金莲木的遗传物质如何在不同的种群间传播,但由于合柱金莲木分布区地形复杂,种群之间被连绵的山地或丘陵隔离,且存在一定的地理距离,地理上的物理隔离势必会给合柱金莲木种群间的遗传物质交流带来困难。合柱金莲木的种群间缺乏基因流,交配只能在个体数十分有限的种群内进行,有限的种群个体自然增加了合柱金莲木的自交或近交概率。另外,种群间缺乏基因流,使得合柱金莲木种群间难免出现遗传分化,从而使该物种内部出现明显分化的种群遗传结构。
许多珍稀濒危植物在恶劣环境下,为防止灭绝而采用近交策略来增加后代个体数[36]。对于植物来说,近交可以提高种群对片段化生境的适应性、减轻传粉媒介数量不足等影响[37],但近交也会导致遗传多样性丧失,最终失去生存和发展的基础[38,39]。长期近交会引起近交衰退从而使后代的生存能力下降,在遇到环境波动时很容易灭绝[40]。濒危植物的遗传多样性通常比较低,生态适应幅度狭窄,近交会致使其种群适应能力进一步下降,并使得有限的遗传多样性进一步丧失,从而加剧濒危程度。合柱金莲木遗传多样性低[8,41,42]且存在近交,且本研究检测到合柱金莲木在物种水平上存在近交衰退(δ=4.332>0),因此交配系统中存在一定程度的近交可能是合柱金莲木濒危的重要因素之一。
4 结论
成龄合柱金莲木植株平均地径、平均株高和平均冠幅分别为0.70、129.67和81.98 cm,平均每株结花序15.56串,有一定比例的成龄植株为丛生植株。与S3分布点相比,S2分布点的成龄植株更大、平均每株花序数更多。合柱金莲木于3月下旬至4月中上旬进入抽(花)序期,4月中下旬进入现蕾期,4月下旬至5月中下旬为开花期,5月下旬至6月上旬进入果实发育期,再经过60-70 d的生长,果实进入成熟期。合柱金莲木的单花开放在1 d内完成,其花序的单花数约为27朵/串,果序的蒴果数约为18个/串,结果植株平均每株的种子数达5 881.68粒,种子量极大。
合柱金莲木在理论上偏离了Hardy-Weinbery平衡,在物种水平上有过剩的纯合子。此外,合柱金莲木的多位点异交率(tm=0.982)高于单位点异交率(ts=0.752),说明在物种水平上合柱金莲木存在一定比例的近交,双亲的近交比例为0.230,且存在近交衰退(δ=4.332>0)。合柱金莲木在物种水平上的多位点相关度(rpm)和单位点相关度(rps)分别为0.272和0.314,其物种内部存在明显分化的种群遗传结构,其异交子代间亲本相关性(rp)为0.213,表明部分子代共享了亲本。交配系统中存在一定程度的近交可能是合柱金莲木濒危的重要因素之一。
