野牛草雌雄株对不同形态氮素的生理响应差异
2023-02-28郭丽珠孟慧珍范希峰滕珂滕文军温海峰岳跃森张辉武菊英
郭丽珠,孟慧珍,2,范希峰,滕珂,滕文军,温海峰,岳跃森,张辉,武菊英*
(1.北京市农林科学院草业花卉与景观生态研究所,北京 100097;2.西北大学生命科学学院,陕西 西安 710069)
由于长期的适应性进化和雌雄性别的分株表达,雌雄异株植物在生长特性、生理生化特征、资源分配、生殖格局和基因表达特性等方面具有一定的性别差异[1-3]。这些差异往往造成雌雄异株植物不同性别的植株对环境适应性不同,且使雌雄植株能在环境中分别获取各自最需要的资源进行营养生长与生殖活动,进而提高种群整体的繁殖率及种群对环境的适应性[4-6]。野牛草(Buchloe dactyloides)原产于北美中部温带和亚热带半干旱地区,是一种少有的雌雄异株草坪植物,其抗寒抗旱能力强、繁殖迅速、维护需求低,在我国引进后广泛应用于水土保持和园林绿化[7]。野牛草具有雌株、雄株及雌雄同株3种性别,不同性别植株的区分只有在进入生殖生长阶段才能进行分辨[8]。野牛草雌、雄株在匍匐茎生长能力[9]、对锈病的抗性[10]及不同生长阶段的保水能力[11]都存在显著的性别差异,这种差异势必影响不同性别野牛草植株的坪用性状有所不同。由于不同性别的植物具有不同的经济价值,因此研究雌雄异株植物在环境条件变化下的响应差异具有重要的生产实践意义,为有目的地选择具有经济、观赏等价值的雌雄异株植物的性别提供理论参考。
施肥是保障草坪草正常生长、维持和提高草坪质量的重要措施[12-13]。硝态氮(NO3--N)和铵态氮(NH4+-N)是植物可以直接从土壤中吸收和利用的两种主要无机氮素形态[14-15],不同氮素形态不仅影响植物的形态建设、光合作用、根系生长[16-18],对植物氮素代谢途径中关键酶活性及相关基因表达也会产生影响[19]。植物对氮素的吸收主要是通过根系完成的,植物根细胞吸收NO3--N后,首先在根系中被硝酸还原酶(nitrate reductase,NR)和亚硝酸还原酶(nitrite reductase,NiR)同化为NH4+,进而被同化为氨基酸和蛋白质。与吸收NO3--N不同,植物吸收NH4+-N后,直接在谷氨酰胺合成酶(glutamine synthetase,GS)/谷氨酸合成酶(glutamate synthase,GOGAT)循环反应的催化下形成谷氨酰胺。不同植物种类对氮素营养环境的长期适应形成对不同氮源的偏向选择性,并形成了不同的氮素利用分配策略。研究表明,硝态氮作为唯一氮源能够增加谷类作物的分蘖数[20],山核桃幼苗(Carya illinoinensis)在生长过程中表现出对铵态氮的优先吸收[17],而黄瓜(Cucumis sativas)在以铵态氮或高比例的铵态氮作为氮源时会受到明显的生长抑制[21],当氮肥中硝态氮的比例较高时则会促进甜椒(Capsicum annuum)、韭菜(Allium tuberosum)、芹菜(Apium graveolens)等作物的生长[22-24]。大量研究表明农作物对氮素形态吸收具偏向选择性,然而氮素形态影响草坪植物的研究十分有限[25-26],因此亟须弥补不同形态氮肥在草坪植物野牛草施用研究上的空白。前人研究表明野牛草草坪综合质量随氮素用量的增加而提高[27-29],但已有研究尚未从氮素形态角度出发探究野牛草对氮素的响应,且很少有研究区分野牛草雌雄株的表现差异,进而从性别差异的角度出发探究野牛草不同性别植株草坪建植中的氮肥管理差异。本研究以不同性别野牛草植株为试验材料,研究不同形态氮素对野牛草雌、雄株在生长、生物量、叶绿素含量、叶片氮含量及氮代谢相关酶的影响,揭示野牛草不同性别植株对不同形态氮素的偏好,为人工建植野牛草单性植株草坪合理施氮、提高野牛草对氮的吸收和利用具有十分重要的理论和现实意义。
1 材料与方法
1.1 试验材料
从野外选取长势良好的野牛草雌、雄单株,通过分株克隆繁殖,获得了大量遗传背景一致、基因型相同的单性别无性系群体。试验时选取生长状态一致的距植株生长顶端第4个克隆分株(一般为8片叶子,图1)进行扦插。

图1 实验对象选取示意图Fig.1 Schematic diagram of experiment object selection
1.2 试验设计
于2021年3月在北京市农林科学院智能温室中进行盆栽试验,盆栽试验的盆钵上口直径21 cm、底部直径15 cm、高27 cm。每盆栽植野牛草雌株或雄株匍匐茎上的无性分株1株,以没有营养成分的黄砂作为培养基质,每盆黄砂重3.5 kg。试验设计4个氮素配比处理:1)不加氮处理(CK);2)单一添加硝态氮N1(NO3--N);3)单一添加铵态氮N2(NH4+-N);4)混合态氮N3(NO3--N∶NH4+-N=1∶1),不同形态氮素的添加主要通过营养液的方式实现,营养液组成如表1所示。为防止试验过程中硝化作用,在N2、N3处理的营养液中添加了硝化抑制剂双氰胺(dicyandiamide,DCD),并通过盐酸和氢氧化钠将营养液的pH调至6.0左右。盆栽幼苗每5~7 d用500 mL营养液浇灌一次,每次浇灌营养液时随机移动花盆位置,保证所有植株光照情况相同,直到试验结束。每个处理12盆,每盆1株,不同性别野牛草植株共计96盆。植物生长3个月后试验结束,收获完整植株。首先采集部分叶片鲜样,测定鲜重后用液氮冷冻保存于-80℃冰箱中用于测定后续生理指标;其次将剩余各部位用蒸馏水进行冲洗,擦干后进行杀青,后置于65℃烘箱中烘干至恒重。
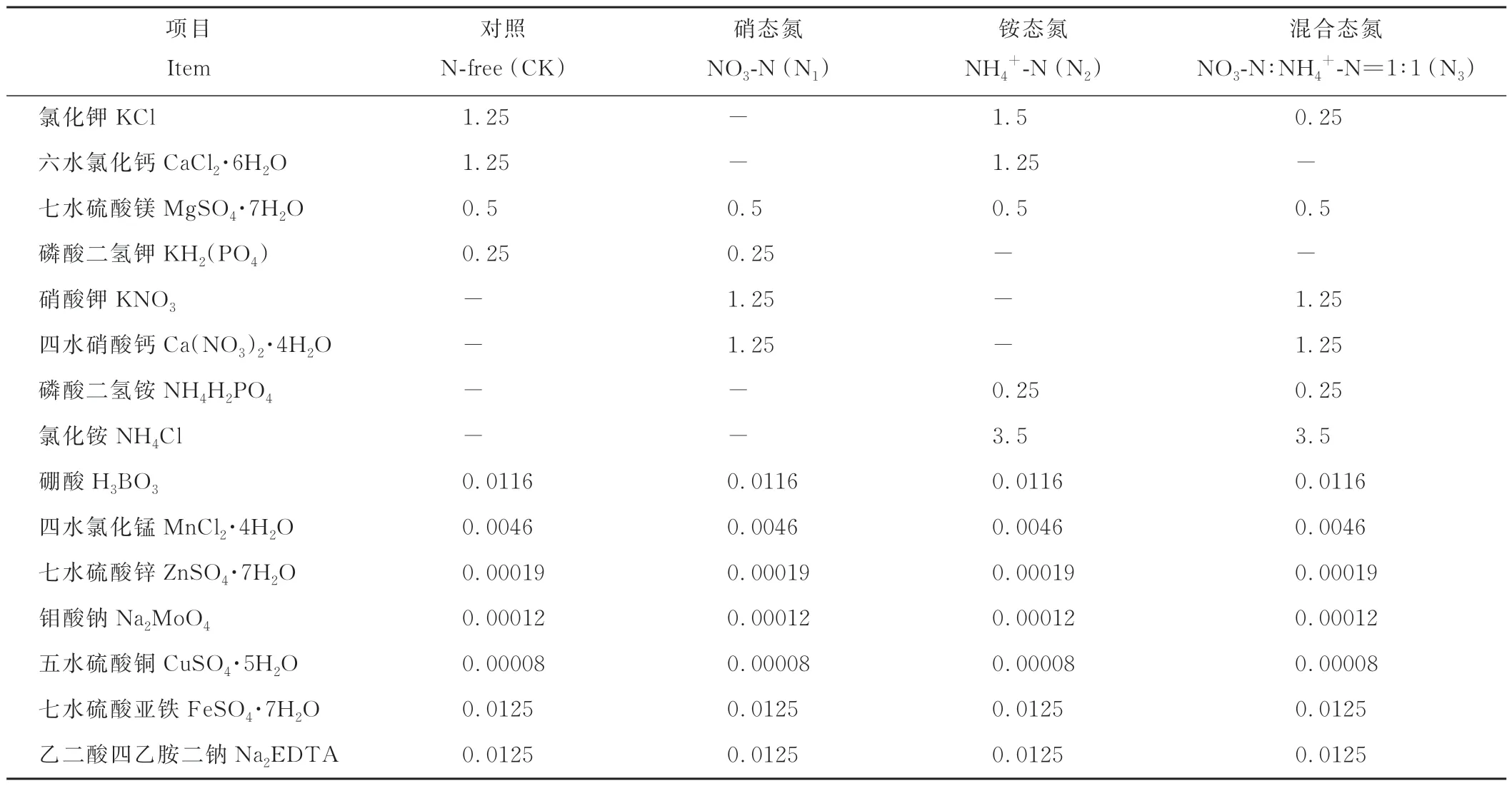
表1 不同形态氮素处理的营养液配方Table 1 Nutrient solution composition of nitrogen form treatments(mmol·L-1)
1.3 试验方法
1.3.1 生长性状指标测定 植株收获后每个处理选取12株测量生长性状指标。匍匐茎长度(cm):用直尺测量匍匐茎尖端到植株底部拉直长度;匍匐茎直径(mm):用游标卡尺测量从枝条底端开始第1、2、3段间隔子直径,取平均值;匍匐茎节数:从枝条尖端到底端全部间隔子的数量;间隔子长度(cm):从枝条底端开始第1、2、3段间隔子长度的平均值。
1.3.2 生物量指标测定 将野牛草雌株或雄株全株分为根、母株、匍匐茎(分株、间隔子),烘干后分别测量各部位生物量干重,并计算地下部分(根)和地上部分生物量(母株、分株、间隔子)以及各部位的生物量分配比例。
根生物量分配比例(%)=根生物量/(根生物量+母株生物量+分株生物量+间隔子生物量)×100
母株生物量分配比例(%)=母株生物量/(根生物量+母株生物量+分株生物量+间隔子生物量)×100
分株生物量分配比例(%)=分株生物量/(根生物量+母株生物量+分株生物量+间隔子生物量)×100
间隔子生物量分配比例(%)=间隔子生物量/(根生物量+母株生物量+分株生物量+间隔子生物量)×100
1.3.3 叶绿素含量的测定 称取野牛草新鲜叶片0.1 g,剪碎后于10 mL乙醇/丙酮(体积比1∶1)混合液内避光浸泡提取18 h,之后使用酶标仪(Multiskan FC,美国)测定在665和649 nm波长下的吸光值。
叶绿素a含量(mg·g-1)=(13.95×A665-6.88×A649)×0.01/0.1
叶绿素b含量(mg·g-1)=(24.96×A649-7.32×A665)×0.01/0.1
总叶绿素含量(mg·g-1)=(6.63×A665+18.08×A649)×0.01/0.1
1.3.4 叶片氮含量的测定 采用碳氢元素分析仪(Elementar vario MACRO cube,德国)测定分株叶片的全氮(total nitrogen,TN)含量。
1.3.5 氮代谢相关酶活性测定 采用北京索莱宝(Solarbio)科技有限公司试剂盒测定硝酸还原酶(nitratereductase,NR)、谷 氨 酰 胺 合 成 酶(glutamine synthetase,GS)和 谷 氨 酸 合 成 酶(glutamate synthase,GOGAT)活性,用酶标仪(Multiskan FC,美国)在340,540和340 nm波长下测量吸光值并计算酶活性。
NR(U·g-1)=5.359×△A340÷W(△A340为反应30 min前后两个吸光值的差值,W为样品重量g)
GS(U·g-1)=38×△A540÷W(△A540为测定管与对照管的吸光值差值,W为样品重量g)
GOGAT(U·g-1)=321.5×△A340÷W(△A340为反应5 min前后两个吸光值的差值,W为样品重量g)
1.4 数据处理
利用Microsoft Excel 2010对原始数据进行记录整理。采用SPSS 21.0对数据进行分析,对不同形态氮素处理下野牛草雌雄株生长性状、生物量及生物量分配、叶绿素含量、叶片氮含量及氮代谢相关酶活性的影响进行单因素方差分析(One-way ANOVA)并使用LSD法进行多重比较。使用Origin 2018作图。
2 结果与分析
2.1 不同形态氮素对野牛草雌雄株生长性状的影响
不同形态氮素处理对野牛草雌、雄株的生长性状影响不同(图2)。野牛草雌、雄株的间隔子长度都在N2、N3处理下显著短于N1和CK,特别是在N2处理时野牛草雌、雄株的间隔子长度都是最短,分别为5.79和2.86 cm,而CK与N1之间没有显著差异。野牛草雌、雄株的间隔子直径与间隔子长度表现相近。野牛草雌株的匍匐茎节数在N2和N3处理时,同样显著少于CK和N1,然而雄株匍匐茎节数在N1处理时显著大于其他3种处理。野牛草雌株的匍匐茎长度在N2处理时最短(44.67 cm),而雄株匍匐茎长度在N1处理时显著高于其他3种处理。
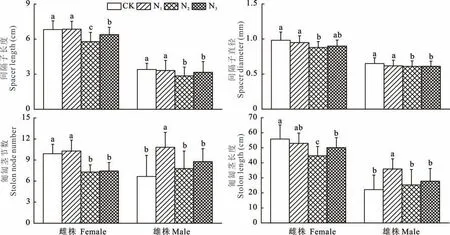
图2 不同氮素形态对野牛草雌雄株生长性状的影响Fig.2 Effects of nitrogen forms on growth traits of female and male buffalograss
2.2 不同形态氮素对野牛草雌雄株不同部位生物量及生物量分配的影响
由图3可知,野牛草雌株子株、根的生物量在N1处理下显著大于其他处理,而N1处理下母株和间隔子的生物量与CK没有显著差异但显著大于其他形态氮素处理。雄株各部位生物量表现与雌株并不一致。雄株的母株生物量在N3处理下最小,子株、间隔子、根生物量在N1处理下最大,其中子株和间隔子的生物量在N1处理和N3处理间无显著差异而根的生物量在N1处理时显著大于N3处理。此外,野牛草雌株和雄株的地上生物量、地下生物量及总生物量均在N1处理时最大。其中雌株总生物量在CK时显著大于N2和N3处理,但地上生物量和地下生物量在N2处理时显著小于CK和N3处理;雄株总生物量在N2处理时显著大于CK,但雄株地上、地下生物量在CK、N2、N3处理之间无显著差异。野牛草雌、雄株均将生物量主要分配于子株。雌株的间隔子生物量分配在N3处理时显著大于N1和CK处理,根生物量分配在N1时显著大于N2;雄株的母株生物量分配在N1、N2处理时显著大于N3,子株生物量分配在N3处理时显著大于N2。
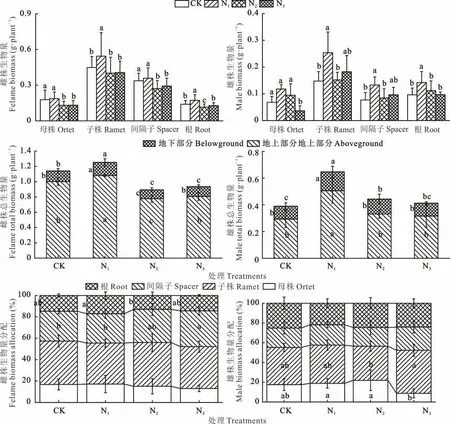
图3 不同氮素形态对雌雄异株野牛草不同部位生物量的影响Fig.3 Effects of nitrogen forms on biomass in different parts of female and male buffalograss
2.3 不同形态氮素对野牛草雌雄株叶绿素、N含量的影响
从表2可以看出,不同形态氮素处理对雌株、雄株叶片氮含量的影响不同,雌株叶片氮含量在N3处理时显著大于其他处理,而雄株在N1处理时最大。此外,不同形态氮素处理主要对野牛草叶片的叶绿素a含量及叶绿素总量的影响显著。雌株、雄株的叶绿素a含量均在N1处理时显著大于其他处理,雄株叶绿素a含量在CK时最低,雌株叶绿素a含量在CK、N2、N3之间无显著差异。雌、雄株的总叶绿素含量在N1处理时均最高,但雄株在N1时显著大于N3,而雌株在N1和N3处理之间无显著差异。总的来说,无论差异显著与否,野牛草雌、雄株的叶绿素含量均在N1处理时高于其他处理。
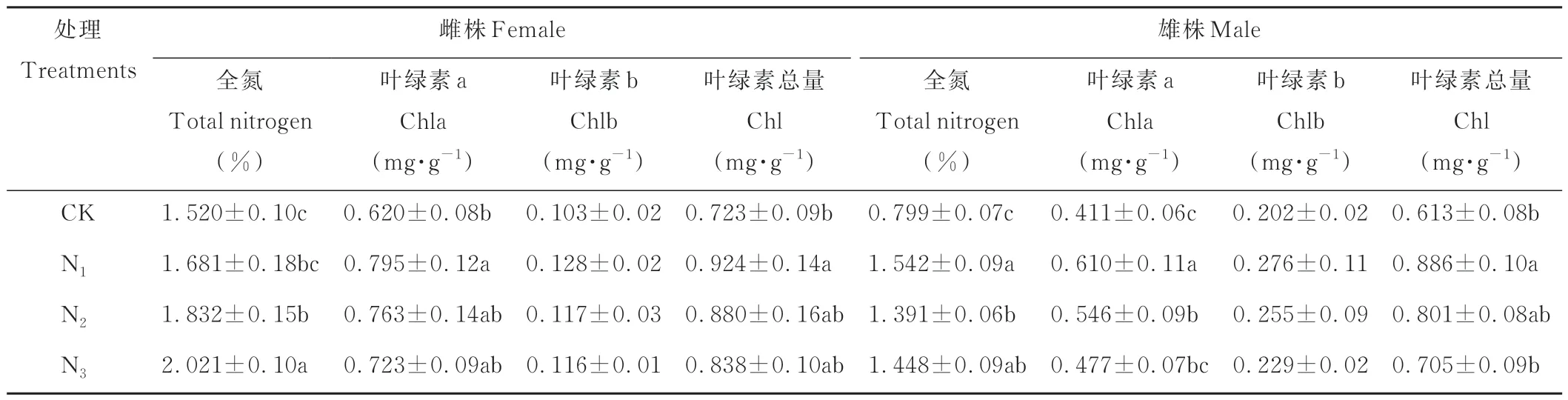
表2 不同形态氮素对野牛草雌雄株叶绿素含量、N含量的影响Table 2 Effects of nitrogen forms on chlorophyll and nitrogen content of female and male buffalograss(mean±SD)
2.4 不同形态氮素对野牛草雌雄株氮代谢相关酶活性的影响
如图4所示,野牛草雌株的硝酸还原酶、谷氨酰胺合成酶及谷氨酸合成酶的活性均在N3处理时显著大于其他处理,其中雌株的NR、GOGAT活性在N1处理时与CK无显著差异、N2与CK有显著差异而GS活性在CK、N1、N2处理之间无显著差异。野牛草雄株NR活性在N1处理时显著大于N2和CK处理,但与N3处理无显著差异,GOGAT活性在N3处理时显著大于CK但与N1和N2无显著差异,GS活性在N1处理时显著大于其他处理。
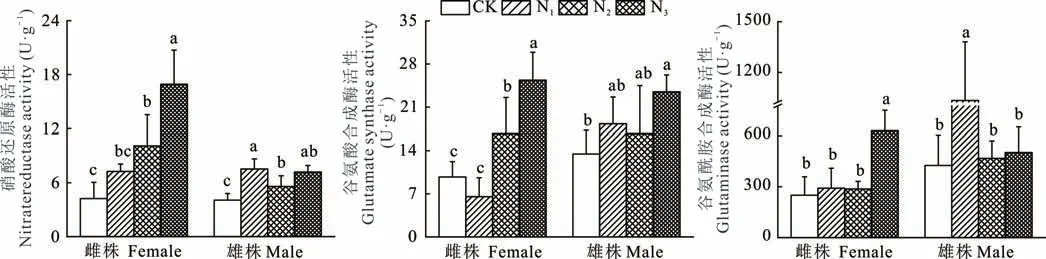
图4 不同形态氮素对野牛草雌雄株氮代谢相关酶活性的影响Fig.4 Effects of different nitrogen forms on activities of nitrogen metabolizing enzymes in female and male buffalograss
3 讨论
自Darwin[30]首次认识到雌雄异株植物的生殖差异将导致性别特化和对资源的不同需求以来,植物界中雌雄植株间的各种差异逐步引起人们的注意。野牛草是禾本科植物中少有的雌雄异株植物,性别分化使野牛草单性别植株草坪的建植成为可能。通常,野牛草进入花期后,雄株的花轴高于株丛,花粉成熟后草坪呈现明显的黄色,因此雌株逐渐成了坪用型野牛草。此外,有研究表明野牛草雌株匍匐茎的生长能力要强于雄株[9]、对锈病的抗性更强[10],因此建植野牛草纯雌株草坪可能比现存混合性别草坪更具性价比。然而,早期学者更多地关注野牛草性别分化比例、性别形态学差异及性别决定等研究[10,31],对野牛草响应不同环境条件的性别差异研究并不多。
氮肥是草坪草需要量最大的营养元素,在草坪植物的生长质量和产量形成中扮演着关键角色。前人研究结果表明,野牛草草坪综合质量随氮素用量的增加而提高[27-29],而本研究发现,在野牛草克隆分株培养过程中N的施加效果并未像其他研究那么明显。这主要是因为:与其他草坪草相比,野牛草是一种低养护、管理粗放的草坪草,其植株生长可能对N养分的丰缺在生长前期并不十分敏感;其次,本试验中施用N素浓度并不高,进而导致N素的施加与对照对野牛草植株生长的影响差别不明显。通常,氮肥对维持和改善草坪质量的作用效果除与施用量有关外[32-33],还受氮素形态的影响[26]。大量研究表明,植物对不同形态氮素的偏好性首先表现为生物量积累的差异[34],这可能是因为植物对不同形态氮素的偏好性与植物的光合能力对不同形态氮素响应差异有关。本研究发现野牛草雌株或雄株均在硝态氮处理(N1)时其总生物量、地上生物量、地下生物量都显著最大,表明野牛草雌、雄株的生长均偏喜硝态氮。尽管如此,野牛草雌、雄株在响应铵态氮作为唯一氮源时的表现却不一致,雌株的地上、地下生物量在铵态氮处理时均显著小于铵硝混合氮处理及对照,而雄株反应不明显,这表明野牛草雌株在以铵盐为唯一氮源的情况下可能产生铵盐毒害症状。通常,NH4+-N抑制植物生长主要与供应NH4+-N导致养分(如K+、Ca2+、Mg2+等阳离子)的吸收减少有关,进而打破植物细胞内的离子平衡[35-36]。此外,本研究还发现氮素形态对野牛草生长性状的影响与其生物量结果表现相近。
植物生长代谢、光合作用和同化物质的基础之一是叶绿素,在一定程度上,叶绿素含量与光合作用的强度关联密切。氮素形态几乎影响了光合作用的各个环节,包括通过影响叶片叶绿素含量、光合速率、暗反应主要酶活性以及光呼吸等,直接或间接影响光合作用。相比对照处理,施氮可提高野牛草雌、雄植株叶绿素含量,其中叶绿素a含量变化比叶绿素b更加明显。硝态氮处理(N1)时野牛草雌、雄株的叶绿素a含量都显著大于CK,说明硝态氮处理显著提高了野牛草的光合作用,有利于光合产物的积累,从而促进其生物量的积累;而铵态氮或铵硝混合态氮处理对叶绿素a含量的影响效果相较CK并不显著。铵态氮营养对叶绿素的增加影响小的原因可能是铵离子在体内积累,产生毒害作用,从而破坏类囊体结构,导致质子形成受阻,且易形成氧化磷酸化,破坏碳水化合物的形成,降低植物的光合特性[37]。此外,氮素是叶绿素分子结构的构成要素,供应充分才能保证叶绿体形成和累积,提高叶绿素含量。野牛草雌株叶氮含量在铵硝混合态氮(N3)处理时显著大于单一氮形态处理,表明在铵态氮中加入硝态氮可以缓解铵态氮对野牛草雌株生长的抑制作用,而混合态氮对雄株的影响没有雌株明显。
氮代谢是植物生命活动和生长发育必不可少的代谢活动之一,而硝酸还原酶(NR)、谷氨酰胺合成酶(GS)、谷氨酸合成酶(GOGAT)是植物体内从无机氮变为有机氮过程中最重要的氮代谢相关酶[19]。NR活性大小表示氮代谢能力的强弱,故它在整个植物氮代谢的过程中起到关键调控作用。影响NR活性的因素有很多,一方面该酶活性高低会随底物浓度的升高而增加,即NR活性受硝酸盐的正向调节;另一方面也有研究认为作物在铵硝混合施用时的NR活性比单一NO3--N营养高,氮素代谢强[38]。本研究结果显示野牛草雄株的NR活性在硝态氮处理时最大,而雌株却在混合态氮处理时最大,表明影响野牛草雌、雄株NR活性的因素并不一致。此外,GS是植物氮素同化中的另一个重要酶类,和GOGAT共同完成氨的初始同化。在GS作用下,NH4+和谷氨酸反应形成谷氨酰胺,然后由GOGAT催化谷氨酰胺与а-酮戊二酸作用形成两分子的谷氨酸,其中一分子的谷氨酸作为GS的底物继续同化NH4+,另一分子谷氨酸可用于合成蛋白质、核酸等含氮化合物[39]。GS对NH4+有高度亲和性,可以使植物避免铵积累所造成的毒害[40]。野牛草雌株在混合态氮(N3)处理时其GS活性最大,这可能是由GS催化形成的产物谷氨酰胺能够反馈抑制GS活性,由于雌株对铵敏感、NH4+处理对雌株GS的活性可能产生了下调作用。然而,并非所有植物GS基因都是由NH4+诱导的,NO3-亦能诱导GS基因的表达[41]。野牛草雄株GS活性在硝态氮处理时达最大,这与其植株体内可能存在不受铵态氮调控的GS相关基因表达有关。GOGAT也是谷氨酰胺合成酶/谷氨酸合成酶循环途径中起催化作用的关键酶,野牛草雌株在混合氮处理时其GOGAT活性显著大于单一硝态氮或铵态氮处理,说明混合氮处理可以减轻铵态氮对雌株的铵毒作用;而雄株GOGAT活性在3种形态氮处理下无显著差异,表明雄株在硝态氮处理下仍可以保持与铵态氮或混合氮处理时相近的GOGAT活性,确保外源NO3-被植物根系吸收后通过木质部运到叶中还原为NH4+而被GS/GOGAT循环同化。总的来说,雌株的氮代谢相关酶在N3处理时均最高,而雄株则是在N1处理时最大,该结果与叶片N含量的结果一致。
4 结论
综上,野牛草雌、雄株的生长均偏喜硝态氮,雌株对铵态氮作为唯一氮源异常敏感而雄株反应不明显。野牛草雌雄株对不同形态氮素的响应存在明显差异,可为人工建植野牛草单性植株草坪合理施氮、提高野牛草对氮的吸收和利用提供科学依据。
