海岛棉β-胡萝卜素异构酶GbD27-6基因克隆及表达分析
2023-02-21刘晨曦朱雨婷赵文杰
刘晨曦,朱雨婷,周 强,陈 瑾,赵文杰,郑 凯,2
(1.新疆农业大学农学院/农业生物技术重点实验室,乌鲁木齐 830052; 2.新疆农业大学博士后流动站,乌鲁木齐 830052)
0 引 言
【研究意义】植物株型是影响植物生长发育的关键因素,受到温度、光周期、营养条件等外界环境因素以及遗传和植物内源激素等内部因素调控,其中,遗传和内源激素占据主导地位[1]。此外,分枝发育对植物株型十分重要,该过程主要受到一类新型激素独脚金内酯(strigolacyone,SL)的精准调控[2]。独脚金内酯是一种多功能激素,有助于控制枝条分枝,并且作为植物与丛枝菌根真菌之间以及寄主植物与寄生独脚金植物之间的信号分子。独脚金是一类重要的植物激素,调控了植物的生长、发育和响应环境刺激的过程。其中D27基因使植物能够合成和调控独脚金激素水平,从而促进促进幼芽伸长进而增加植株高度、调控植物的花芽形成和开花过程、参与果实的发育和成熟过程、在种子的萌发和休眠状态的转变中发挥作用等。活性的独脚金内酯会进一步代谢为其他形式的独脚金激素,这些激素在植物的不同组织和器官种起到特定的生理调节作用。需要注意的是,独脚金合成途径是一个复杂的网络,包括多个酶、基因和反应。【前人研究进展】在缺乏磷的情况下,植物根会产生大量的独脚金内酯分泌物,是菌根形成共生过程重要的信号分子[3-5]。类胡萝卜素脱氧、氧化生成独脚金醇,继而转化成不同的独脚金内酯物质[6,7]。该合成通路涉及到许多基因,当这些基因发生变化时,植株的生长发育随之受到影响。独脚金内酯对番茄磷的吸收上发挥作用[8]通过调控赤霉素合成参与拟南芥种子萌发[9]、以及调控主根伸长及抑制侧根生长作用等[10]。棉花作为一种深根作物,根系十分发达[11],独脚金内酯通过调控根系的生长从而影响营养和水分的吸收[10],进而影响棉花纤维的发育。以玉米、高粱等植物的类胡萝卜素缺失突变体并加以类胡萝卜素生物合成抑制剂的处理,发现SL来源于类胡萝卜素生物合成途径[7,12],属于其衍生的化合物。拟南芥、水稻和豌豆等不同植物的SL缺失突变体表现出更多分枝的表型,并且该表型可以通过外源施加其人工合成类似物GR24得到恢复,从而揭示出植物体SL的生物合成起始于质体[13]。该过程至少存在5种酶的参与[1,14]。D27、CCD7与CCD8都定位于质体内部,P450则位于质体外部。独脚金内酯属于类胡萝卜素衍生的一种根际激素信号分子,而D27是其重要的转录基因,可以产生独脚金内酯前体或中间产物[1,6]。【本研究切入点】棉纤维发育的起始期、伸长期、次生壁增厚期、成熟期4个时期中,伸长期和次生壁增厚期同时开始,产生了交叉的生长时域[15]。棉花纤维的遗传机制极其复杂。现在已知独脚金内酯是调控植物分枝发育的关键因子,但是关于独脚金内酯在棉花纤维发育中作用尚不清楚,其功能机制、转导途径等方面还不清楚[10,16,17]。需研究GbD27-6基因在海岛棉分枝和侧枝生长中的影响作用。【拟解决的关键问题】通过对海岛棉GbD27-6基因的克隆和表达分析,研究其在棉纤维的表达特性及独脚金内酯在海岛棉纤维生长发育过程中的作用,为进一步研究D27基因的功能奠定理论基础。
1 材料与方法
1.1 材 料
海岛棉材料PimaS-7由新疆农业大学农学院作物遗传改良与种质创新重点实验室棉花分子遗传育种课题组所提供。海岛棉种植于新疆巴音郭楞蒙古自治州农科所试验站。
1.2 方 法
1.2.1 海岛棉GbD27-6基因克隆及生信
使用天根生物植物总RNA提取试剂盒,用于提取海岛棉PimaS-7在5、10、15、20、25、30 d各个时期的棉纤维总RNA(以开花当天作为纤维发育0 d)。将提取的RNA进行质量检测并在1%琼脂糖凝胶电泳鉴定,合格后使用反转录试剂盒生成cDNA第一链。多数物种D27基因家族都存在一个名为DUF4033的蛋白保守结构域,将其导入棉花基因组数据库在线比对得到D27基因蛋白保守序列,利用此保守结构域序列在海岛棉基因组数据库中进一步比对得到海岛棉GbD27-6基因。
以海岛棉PimaS-7的cDNA为模板,使用50 μL扩增体系:TransStartTaqDNA Polymerase 250 U(全式金)高保真酶25 μL、cDNA 2 μL、Forward primer和Reverse primer各1 μL,其余加Nucleasw-free Water 21 μL补齐体系。扩增目的基因反应程序为预变性94℃ 5 min;变性94℃ 30 s,退火62.4℃ 30 s,延伸72℃ 1 min,35个循环;延伸72℃ 10 min获得目的基因片段。表1
利用琼脂糖凝胶回收试剂盒,回收特异性扩增的目的基因,与T5线性化载体25℃连接30 min,重组载体转入到大肠杆菌感受态细胞中通过卡纳霉素标记进行筛选,并挑选单菌落于37℃摇床培养。菌液PCR阳性鉴定后,将阳性菌液送至生工生物工程(上海)股份有限公司进行测序。

表1 引物序列
1.2.2 海岛棉GbD27-6基因的表达特性
分别提取海岛棉PimaS-7在不同纤维发育时期、不同花器官的总RNA,采用反转录试剂盒进行反转录。采用荧光定量试剂盒按说明书对荧光定量体系进行配置[13]。选择UBQ7作为内参基因。实时荧光定量PCR分析平台为美国应用生物系统公司产品。单次试验中每个样本设置3次生物学重复,以内参基因UBQ7作为对照。
1.3 数据处理
利用诸多在线软件进行生物信息学分析,用SPSS 19.0软件进行差异显著性分析,P<0.05表示差异显著。表2
2 结果与分析
2.1 海岛棉GbD27-6基因的克隆
研究表明,条带大小为850 bp左右,与预测大小一致。GbD27-6基因长为816 bp,编码271个氨基酸。PCR阳性菌液产物显示符合上述预测及分析结果。图1,图2

表2 生信分析软件
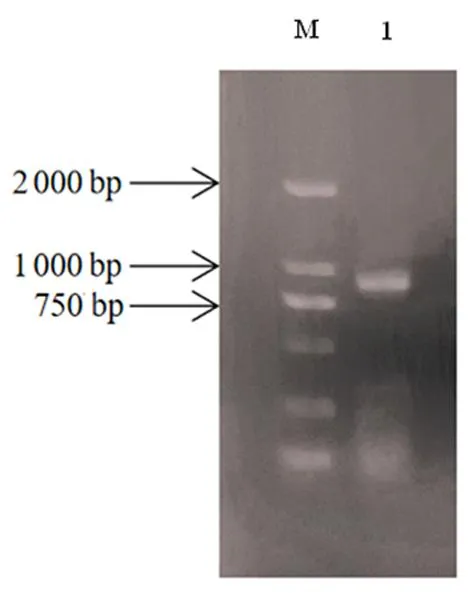
注:M:DL2000 DNA maker;1:GbD27-6基因的扩增产物
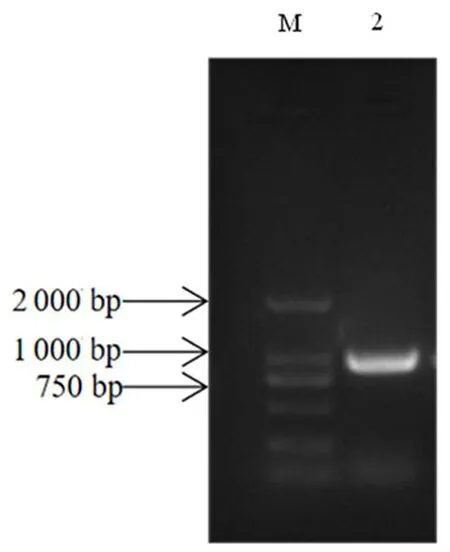
注:M:DL2000 DNA maker;2:GbD27-6基因菌液的阳性鉴定
2.2 海岛棉GbD27-6蛋白理化性质及亚细胞定位预测
研究表明,GbD27-6蛋白含有8个外显子和7个内含子,分子式为C1335H2134N350O389S24,分子量为30.081 18 kDa,理论等电点(pI)为8.40(>7),脂肪系数为85.57,不稳定系数为50.15(>40),该蛋白为不稳定蛋白。疏水氨基酸残基数目多于亲水氨基酸残基数。该蛋白属于疏水性蛋白。GbD27-6蛋白主要在细胞膜分布,其为分泌性蛋白。图3
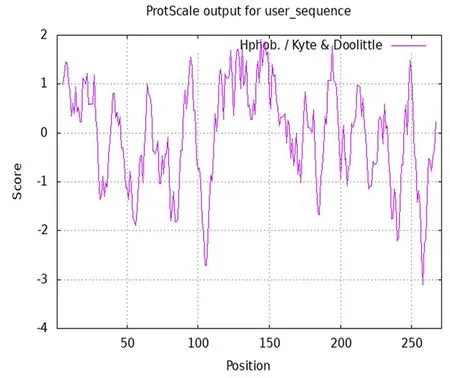
图3 海岛棉GbD27-6的亲疏水性Fig.3 Hydrophilic and hydrophobic analysis of GbD27-6 in Gossypium barbadense
2.3 海岛棉GbD27-6蛋白质高级结构预测
研究表明,使用在线软件对GbD27-6蛋白二级结构预测,二级结构中有133个无规则卷曲(占49.08%)、88个α-螺旋(占32.47%)、37个延伸链(占13.65%)、13个β-转角(占13.80%)。该蛋白属于异质二聚体结构蛋白,推测该蛋白需要与其他蛋白质结合发挥功能。表3,图4
2.4 海岛棉GbD27-6蛋白质信号肽、跨膜区、保守结构域及磷酸化位点预测
研究表明,该基因不具有信号肽,GbD27-6蛋白不存在跨膜结构。图5,图6
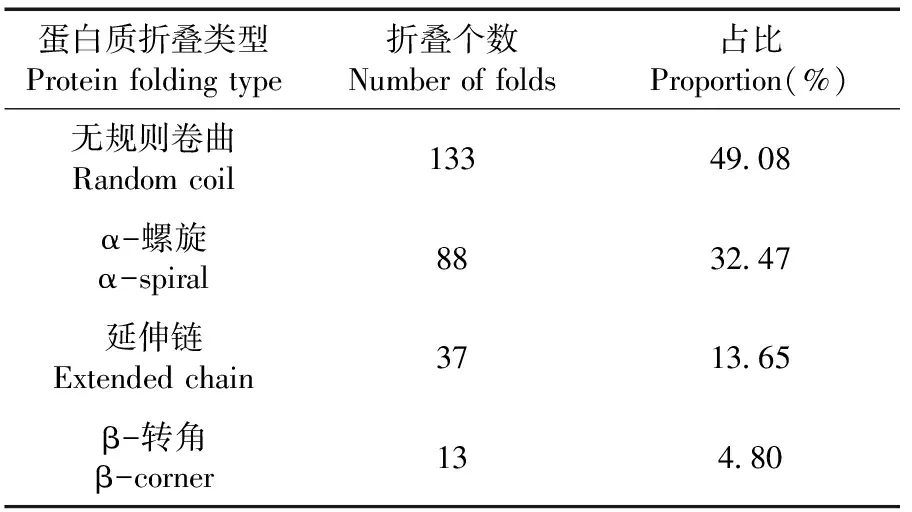
表3 GbD27-6蛋白二级结构预测
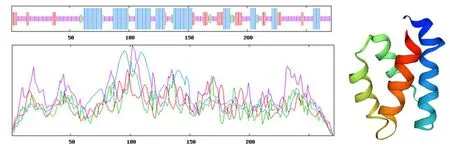
图4 GbD27-6蛋白三级结构预测Fig.4 Tertiary structure prediction of GbD27-6 protein
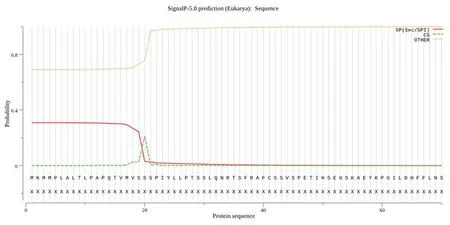
图5 海岛棉GbD27-6的信号肽预测Fig.5 Prediction of signal peptide of GbD27-6 in Gossypium barbadense
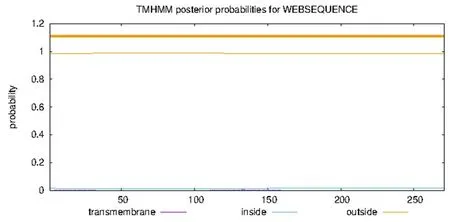
图6 海岛棉GbD27-6的跨膜区预测Fig.6 Prediction of transmembrane region of GbD27-6 in Gossypium barbadense
GbD27-6蛋白是DUF4033超家族的成员,含有DUF4033结构域的蛋白质,与拟南芥叶绿体β-胡萝卜素异构酶D27高度类似,通过催化全反式β-胡萝卜素中C9-C10双键的异构化导致9-顺式β-胡萝卜素并为CCD7提供底物,参与链内酯的生物合成。其结合位点出现于靠近多肽链N端一端。结构域与蛋白质完成生理功能有着密切的关系,预测该结构域占据重要的作用。图7
该蛋白质磷酸化位点中丝氨酸磷酸化多于苏氨酸多于酪氨酸。图8

图7 海岛棉GbD27-6的保守结构域Fig.7 Conserved structural domain of GbD27-6 in Gossypium barbadense
2.5 海岛棉GbD27-6基因保守基序、多序列比对和系统进化树
研究表明,筛选NCBI数据库的信息,选取7条同源性较近的物种序列(巴西海岛棉(KAB2003250.1)、陆地棉(XP 016695906.1)、戴维逊棉(MBA0617518.1)、松散棉(MBA0715122.1)、旱地棉(MBA0686082.1)、可可树(XP 007051131.2;EOX95288.1)),都含有3个相同的保守基序;经DNAMAN进行多序列比对,结合保守结构域预测分析结果找出DUF4033蛋白结构域。图9,图10
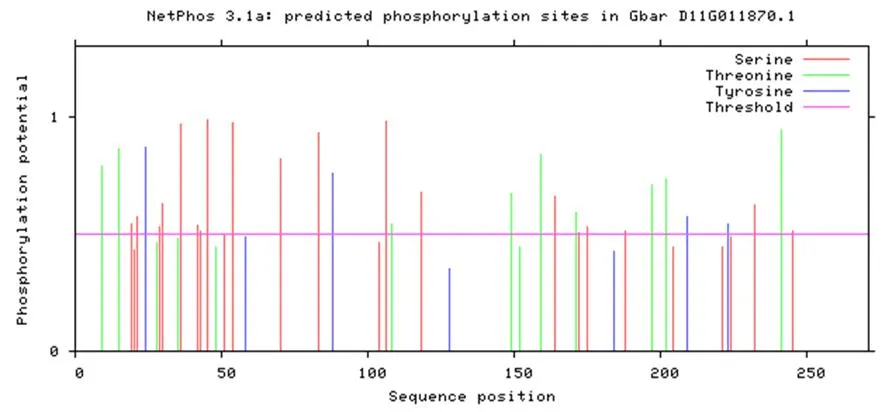
图8 海岛棉GbD27-6的磷酸化位点Fig.8 Phosphorylation site of GbD27-6 in Gossypium barbadense

图9 GbD27-6与其他植物保守基序预测分析Fig.9 Predictive analysis of GbD27-6 and conservated motif in other plants

图10 GbD27-6与其他植物氨基酸序列比对Fig.10 Analysis of amino acid sequence alignment between GbD27-6 and other plants
海岛棉GbD27-6蛋白除与巴西海岛棉(KAB2003250.1)、陆地棉(XP 016695906.1)等棉属的蛋白距离较近外,还与可可属可可树(XP 007051131.2)、榴莲属榴莲(XP 022757850.1)等的蛋白距离较近;与梨属白梨(XP 009349930.2)、猕猴桃属猕猴桃(PSS14736.1)的距离较远。海岛棉GbD27-6基因可能与可可树D27具有相似功能(图中海岛棉(GbD27-6);巴西海岛棉(KAB2003250.1;PPD89448.1;PPS11066.1);陆地棉(XP 016695906.1;XP 016710237.1;KAG4119971.1;XP 016695907.1;XP 040936992.1);戴维逊棉(MBA0617518.1);松散棉(MBA0715122.1);旱地棉(MBA0686082.1);雷蒙德氏棉(XP 012490306.1;XP 012490307.1;KJB41793.1);拟似棉(MBA0741465.1);司笃克氏棉(KAH1081179.1);黄褐棉(TYJ09133.1);树棉(XP 017631093.1;KHG05677.1)斯温迪芒氏棉(MBA0861344.1);哈克西尼棉(MBA0802487.1);毛棉(TYI00282.1);可可树(XP 007051131.2;EOX95288.1);榴莲(XP 022757850.1);木槿(XP 039007481.1);长蒴黄麻(OMO82110.1);板栗(KAF3964893.1);芒果(XP 044501650.1;XP 044501649.1;XP 044501652.1);美洲山核桃(KAG7967178.1;KAG2693308.1;XP 042991374.1);欧洲栓皮栎(XP 023904362.1);核桃(XP 018822128.1);番木瓜(XP 021904458.1);加利佛尼亚白栎(XP 030948830.1);文冠果(KAH7578239.1);灰籽南瓜(KAG7025594.1))。图11
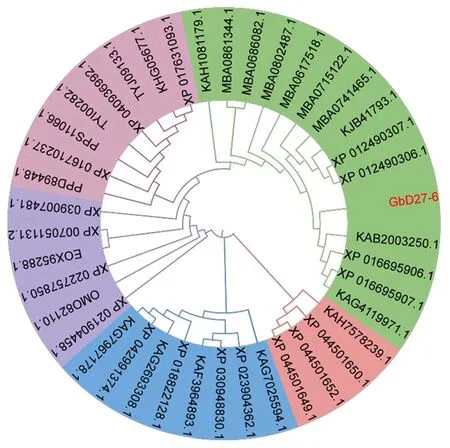
图11 海岛棉GbD27-6蛋白系统进化树Fig.11 Phylogenetic tree analysis of GbD27-6 protein in sea island cotto
2.6 海岛棉GbD27-6基因在不同组织中的表达特性
2.6.1GbD27-6基因在海岛棉不用组织中的的表达
研究表明,GbD27-6基因在花瓣、花药、苞叶中均有表达,但其表达量在不同器官中存在差异。GbD27-6基因在苞叶中表达量最高,显著高于其他两个器官(P<0.05);在花瓣、花药中表达量较低,两者中表达量差异不显著,花药表达量略高于花瓣。苞叶中的相对表达量约为花药的34倍,约为花瓣的48倍。GbD27-6基因在花瓣、花药、苞叶中表达量的比例约为1∶1.4∶48.2。GbD27-6基因或许在花期起着不可替代的作用。图12
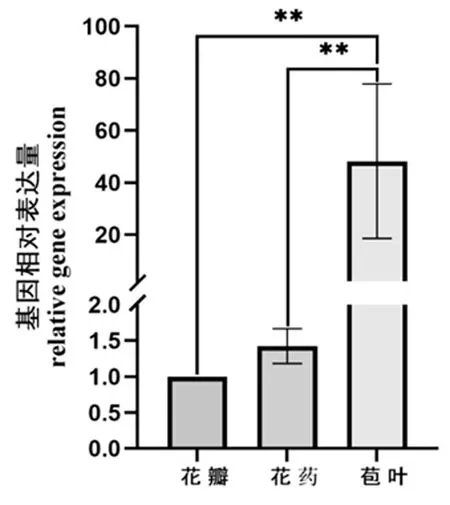
注:*、**和***表示同一基因不同处理间在0.05、0.01、0.001水平差异显著,下同
2.6.2GbD27-6基因在海岛棉不同纤维发育时期的表达特性
研究表明,GbD27-6基因在纤维发育20 d时表达量最高,显著高于其它时期(P<0.05);在纤维发育30 d时次之;在纤维发育5、25和35 d时再次之;在纤维发育0和10 d中表达量最低。GbD27-6基因可能在纤维的伸长期有着重要的作用。图13
3 讨 论
3.1D27基因在其中途径起到了特定的催化作用,是整个途径的重要组成部分。沈月等[10]研究发现,独脚金内酯与植物激素互作对植株根系产生影响。在番茄幼苗培养中,独脚金内酯的处理可以增加野生型、max1和max4的分生区皮层细胞数目而促进主根伸长,作用机制为抑制生长素转运,增加分生区和过渡区大小,促进主根伸长;与细胞分裂素互作调控主根的发育;与乙烯协同促进根毛伸长。张艳培[14]等通过试验发现,f2-132突变体是水稻中编码CCD7酶的D17/HTD1基因发生点突变,导致独脚金内酯无法合成,从而表现出半矮化多分蘖的表型,利用独脚金内酯人工合成类似物GR24进行处理,分蘖表型又恢复正常,证明了f2-132的矮化多分蘖表型是由于独脚金内酯合成受阻造成的。D27基因属于DUF4033超家族成员,含有特有结构域的蛋白质类似于拟南芥叶绿体β-胡萝卜素异构酶D27,该基因通过催化全反式β-胡萝卜素中C9-C10双键的异构化导致9-顺式β-胡萝卜素为CCD7提供底物,参与链内酯的生物合成。研究从海岛棉品种PimaS-7纤维中克隆获得GbD27-6基因,生信分析显示与其他植物D27基因具有较高的相似性,证明了所克隆基因的准确性。对其氨基酸序列分析发现,该基因编码271个氨基酸,编码的蛋白不稳定,属于疏水蛋白且不存在跨膜结构。对GbD27-6蛋白的二级结构预测结果显示其二级结构由无规则卷曲、α-螺旋、延伸链和β-转角构成,以无规则卷曲和α-螺旋为主,具有D27蛋白的典型特征。三级结构预测表明蛋白属于异质二聚体蛋白,推测与其他蛋白互作形成二聚体发挥功能。根据系统进化树分析推测海岛棉GbD27-6的蛋白序列与陆地棉,可可、榴莲有较近的进化关系,与白梨,猕猴桃进化关系较远。
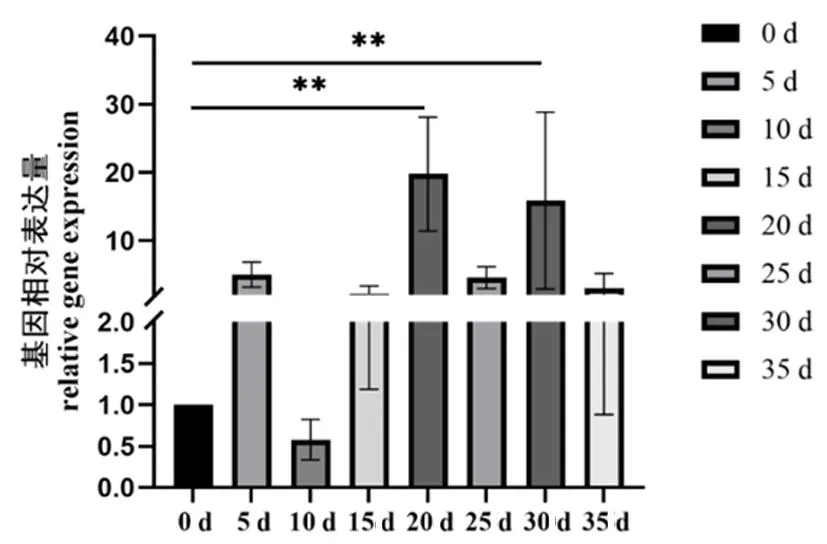
图13 GbD27-6基因在海岛棉不同 纤维发育时期的基因表达量
3.2植物中GbD27-6基因在不同组织、不同发育时期的表达存在差异,通过基因表达的组织差异性可以预测基因对海岛棉纤维发育可能存在调控作用。根据海岛棉三代全长转录组数据库筛选到GbD27-6基因序列,利用在线软件设计特异性引物,以海岛棉PimaS-7的cDNA为模板扩增GbD27-6基因[17,18]。海岛棉GbD27-6基因在苞叶、花药、花瓣中均有表达,但存在较大差异。其在苞叶中表达量最高,显著高于花药、花瓣的表达量。在苞叶中的表达量约为花药的35倍、花瓣的45倍。孙洪影[1]研究结果表明,草莓FveD27基因表达量在新叶与茎尖部位较高,其次是叶柄和老叶。该研究的组织器官虽有所不同,但同样证明了D27基因在苞叶中表达量最高。不同发育时期的基因表达量也差异较大:20~30 d时表达量达到顶峰,纤维发育5、25和35 d时次之;纤维发育0和10 d中表达量最低。在20 d的基因表达量约为30 d时的1.7倍,5 d时的5.5倍,25 d时的6.05倍,35 d的9.2倍,15 d的12.3倍,0 d的27倍,10 d的48倍。由于开花当天作为棉纤维发育0 d,当棉花纤维发育10 d时,棉花正处于盛花期,单株开花日增长量最多,生殖生长开始强于营养生长,营养分配以供应花铃为主,叶面积指数达到最大值,导致10 d时基因表达量最小。
推测该基因在20 d至30 d时控制纤维发育最强,可在此时期深入研究GbD27基因的调控机理。海岛棉纤维品质优异,使得其优于其他棉属,基于此现状,将进一步深入研究D27基因的生物学功能,为今后开展海岛棉纤维品质、株型等发育研究以及新品种的选育提供理论支持。
4 结 论
从海岛棉Pimas-7中克隆得到了GbD27-6基因,该基因编码区全长816 bp,编码271个氨基酸,蛋白分子式为C1335H2134N350O389S24,蛋白质分子质量大约为30.081 18 kDa,理论等电点(pI)为8.40,属不稳定疏水蛋白,不存在跨膜结构,其二级结构由无规则卷曲、α-螺旋、延伸链和β-转角构成,三级结构属异质二聚体结构蛋白,需要与其他蛋白结合发挥作用。亚细胞定位预测显示该蛋白主要存在于细胞膜。GbD27-6基因在纤维发育20 d至30 d表达量达到顶峰且在10 d时表达量最低。
