辽东山区蒙古栎次生林天然更新特征及其影响因子分析
2023-02-20董莉莉汪成成赵济川刘红民王建军高英旭李兴忠
董莉莉,汪成成,赵济川,刘红民,王建军,高英旭,李兴忠
(1 辽宁省林业科学研究院,辽宁 沈阳 110032;2 本溪满族自治县国营草河掌林场,辽宁 本溪 117108)
森林天然更新是以木本植物为主体的生物种群在时间、空间上的不断延续、发展和演替,是森林生态系统自我繁衍和恢复的重要手段[1],对未来森林群落的变化、功能的实现和生物多样性的丰富具有重要影响[2]。成功的森林天然更新必须具备充足且有生命力的种子,以及适合种子萌发、支持幼苗成活和幼树正常生长的环境条件[3]。森林天然更新主要受更新树种生物生态学特性、林分因子、地形条件以及外界干扰等方面的影响。欧芷阳等[4]研究表明,桂西南蚬木种群天然更新受林分垂直结构和林分密度影响较大。王瑞红等[5]研究认为,急尖长苞冷杉(Abiesgeorgeivar.smithii)更新幼苗数量与苔藓厚度呈极显著正相关。Dyderski等[6]研究发现,适度的光照和较低的土壤pH更有利于挪威云杉幼苗密度的增加。可见,森林类型不同、立地条件不同,森林天然更新影响因素也不同。因此开展森林群落更新特征及更新机制研究,对于了解群落组成和演替过程、实现退化森林植被的恢复具有重要意义。
蒙古栎(Quercusmongolica)为中国北方地带性森林植被的重要组成树种[7],以蒙古栎为建群种的天然次生林广泛分布于中国东北地区。辽宁省蒙古栎林面积181.68万hm2,蓄积量11 622.12万m3,分别占辽宁省天然林面积的71%、蓄积量的63%[8]。蒙古栎林在涵养水源、保持水土以及维持生态系统稳定等方面发挥了重要作用,具有较高的生态和经济价值。近几十年来,由于过度砍伐和不合理经营,蒙古栎林普遍存在更新能力差、幼苗成活率低、幼苗难以长成幼树等现象,迫切需要找出制约蒙古栎林天然更新的关键因子,为制定科学合理的经营措施提供依据。本研究以辽东山区典型蒙古栎天然次生林为研究对象,对其林下更新树种组成和更新特征进行分析,并采用Pearson相关分析法和多元线性逐步回归分析法探讨各环境因子对更新幼苗、幼树的影响,以期为辽东山区蒙古栎天然次生林的恢复和可持续经营管理提供理论依据。
1 研究区概况
研究区位于辽宁省新宾县猴石国家森林公园,地理位置124°26′~124°20′E,41°40′~41°35′N。该区域为长白山系龙岗余脉的延伸部分,平均海拔520 m,最高峰海拔1 131 m,年均气温7 ℃,≥10 ℃的年有效积温2 300~2 500 ℃,年均降水量750~850 mm,相对湿度75%,无霜期150 d,属北温带大陆季风气候区。 土壤类型以暗棕壤和棕壤为主。主要森林类型为蒙古栎林、落叶阔叶混交林、针阔混交林、红松人工林和落叶松人工林。研究区内的主要乔木树种有蒙古栎、水曲柳(Fraxinusmandshurica)、花曲柳(Fraxinusrhynchophylla)、核桃楸(Juglansmandshurica)、色木槭(Acermono)、紫椴(Tiliaamurensis)、黄波罗(Phellodendronamurense)、白桦(Betulaplatyphylla)和刺楸(Kalopanaxseptemlobus)等,灌木树种主要有卫矛(Evonymusalatus)、忍冬(Lonicerajaponica)、接骨木(Sambucuswilliamsii)和东北山梅花(Philadelphusschrenkii)等,草本植物主要有荨麻叶龙头草(Meehaniaurticifolia)和球果堇菜(Violacollina)等。
2 研究方法
2.1 样地设置
在全面踏查的基础上,按照林分因子、林地因子和人为干扰等因子的差异性,选择具有代表性的蒙古栎次生林,设置面积为25 m×25 m的样地,共计24块。对样地内胸径≥5 cm的乔木进行每木检尺,记录树种名称、位置、数量、胸径、树高、冠幅等信息,同时在每块样地内随机设置3个5 m×5 m的更新苗样方、3个2 m×2 m草本样方和3个0.5 m×0.5 m枯落物样方。更新苗样方记录胸径<5 cm的乔木幼苗(株高H≤1 m)和幼树(H>1 m,胸径<5 cm)[9-10]的种类、数量、高度、基径及更新方式等,其中更新方式分为实生和萌生,实生苗是指由种子萌发产生的幼苗,萌生苗是指从侧芽(包括不定芽)或根系产生的幼苗[11];草本样方记录草本种类、平均高度、盖度等;枯落物样方记录四边和中心点枯落物的厚度及盖度等信息。利用手持GPS测定样地的海拔、坡向、坡位、坡度等信息,样地基本情况见表1。
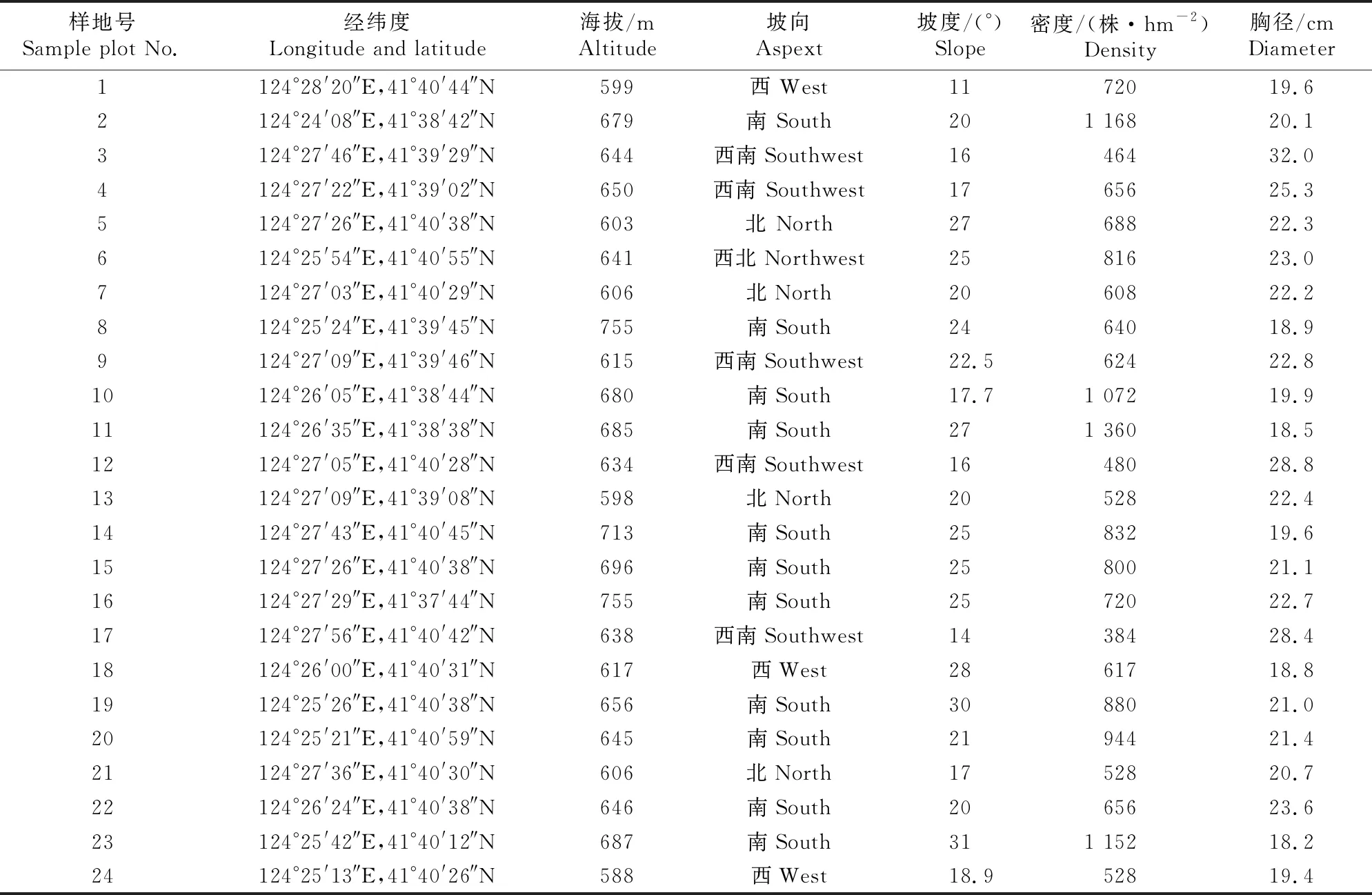
表1 辽东山区蒙古栎次生林样地基本概况
2.2 树种重要值计算
根据野外调查数据,统计蒙古栎林乔木更新幼苗(树)的物种组成,计算每个物种的重要值。其计算公式如下:
重要值=(相对多度+相对盖度+相对频度)/3;
相对多度=(某种幼苗多度/所有幼苗多度和)×100%;
相对盖度=(某种幼苗盖度/所有幼苗盖度和)×100%;
相对频度=(某种幼苗频度/所有幼苗频度和)×100%。
2.3 更新幼苗(树)等级的划分
以更新苗高度(H)为标准,每30 cm划分1个等级,共划分为12个等级:Ⅰ.H<30 cm;Ⅱ.30 cm≤H<60 cm;Ⅲ.60 cm≤H<90 cm;Ⅳ.90 cm≤H<120 cm;Ⅴ.120 cm≤H<150 cm;Ⅵ.150 cm≤H<180 cm;Ⅶ.180 cm≤H<210 cm;Ⅷ.210 cm≤H<240 cm;Ⅸ.240 cm≤H<270 cm;Ⅹ.270 cm≤H<300 cm;Ⅺ.300 cm≤H<330 cm;Ⅻ.H≥330 cm[12]。
2.4 林下光环境测定
采用冠层分析仪(WinScanopy2010a,Canada),在每个更新苗样方中心点距离地面1.5 m处随机拍摄3张半球面林冠照片,通过Win-scanopy和XLS canopy分析软件获取林冠开阔度、叶面积指数和林下总光合辐射等参数。
2.5 土壤样品的采集与测定
沿样地对角线和中心点采用剖面法收集0~20 cm土层的土壤样品,共计120份,用于土壤pH值和养分含量的测定。土壤pH值采用pH计测定,土壤全氮含量采用凯氏定氮法测定,碱解氮含量采用碱解扩散法测定,全磷含量采用硫酸-高氯酸消煮-钼锑抗比色法测定,有效磷含量采用盐酸-氟化铵浸提-钼锑抗比色法测定,全钾含量采用氢氧化钠碱熔-火焰光度法测定,速效钾含量采用乙酸铵浸提-火焰光度法测定,有机质含量采用重铬酸钾容量法-外加热法测定,土壤含水量采用手持土壤水分测定仪(TDR-HH2)测定。
2.6 数据处理
采用SPSS 21.0对幼苗、幼树密度与环境因子进行Pearson相关性分析,采用DPS 18.1软件对幼苗、幼树密度与环境因子进行多元线性逐步回归分析。
3 结果与分析
3.1 辽东山区蒙古栎林乔木树种的更新特征
3.1.1 更新树种的组成特征 表2显示了蒙古栎林更新苗的物种组成及物种特征值。由表2可知,辽东山区蒙古栎林下更新乔木共有30种,隶属于14科21属,其中蒙古栎重要值最大,为12.71%,元宝槭(A.truncatum)、花曲柳、假色槭(A.pseudosieboldianum)和紫椴次之,重要值均大于8%,5个树种的重要值之和占所有更新苗重要值之和的近57%,为乔木更新苗中的优势种;其次为千金榆(Carpinuscordata)、怀槐(Maackiaamurensis)、暴马丁香(Syringareticulatasubsp.amurensis)、鼠李(Rhamnusdavurica)和稠李(Padusavium)等,重要值均大于3%。总体而言,蒙古栎次生林乔木更新物种组成丰富,能够为森林群落结构的自然优化提供一定的保障。
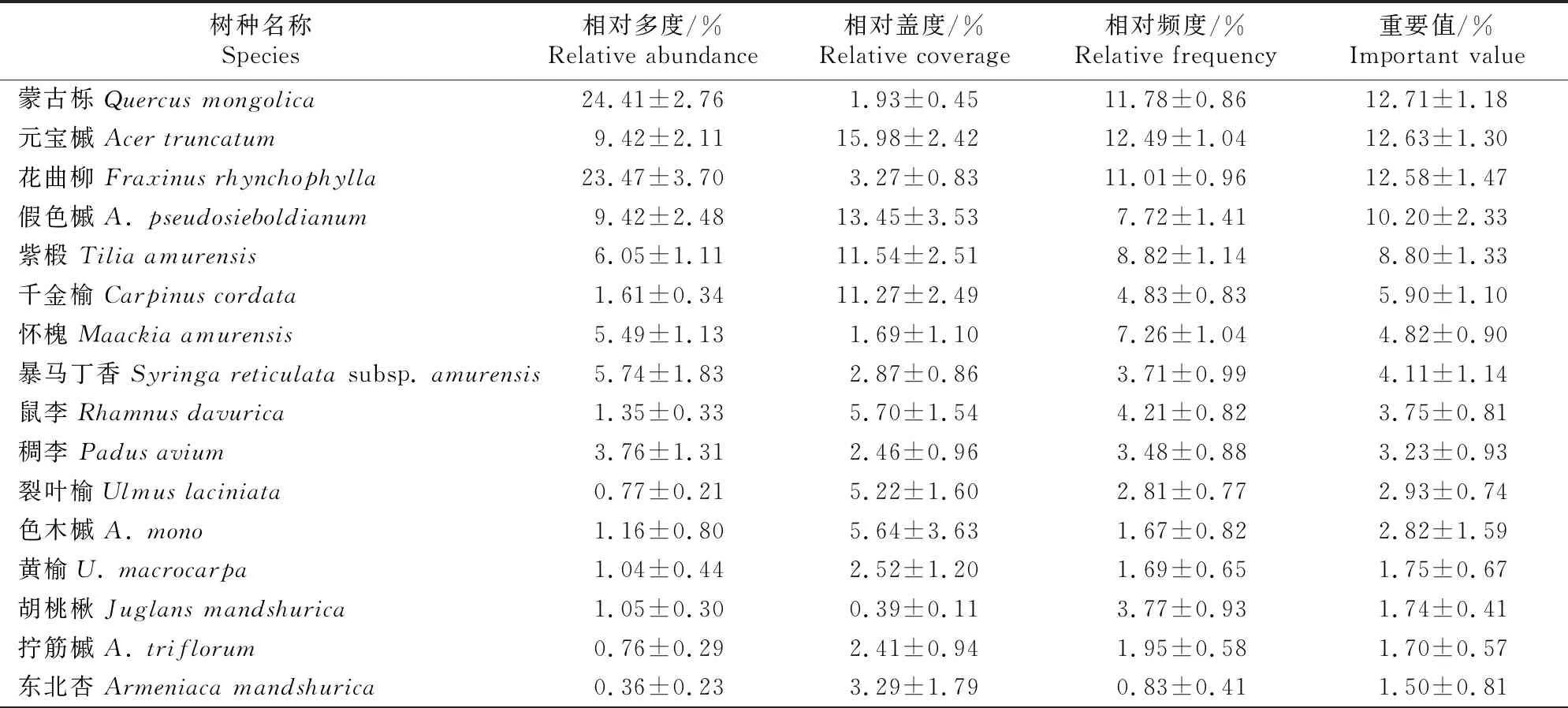
表2 辽东山区蒙古栎林乔木幼苗(树)物种组成及特征值
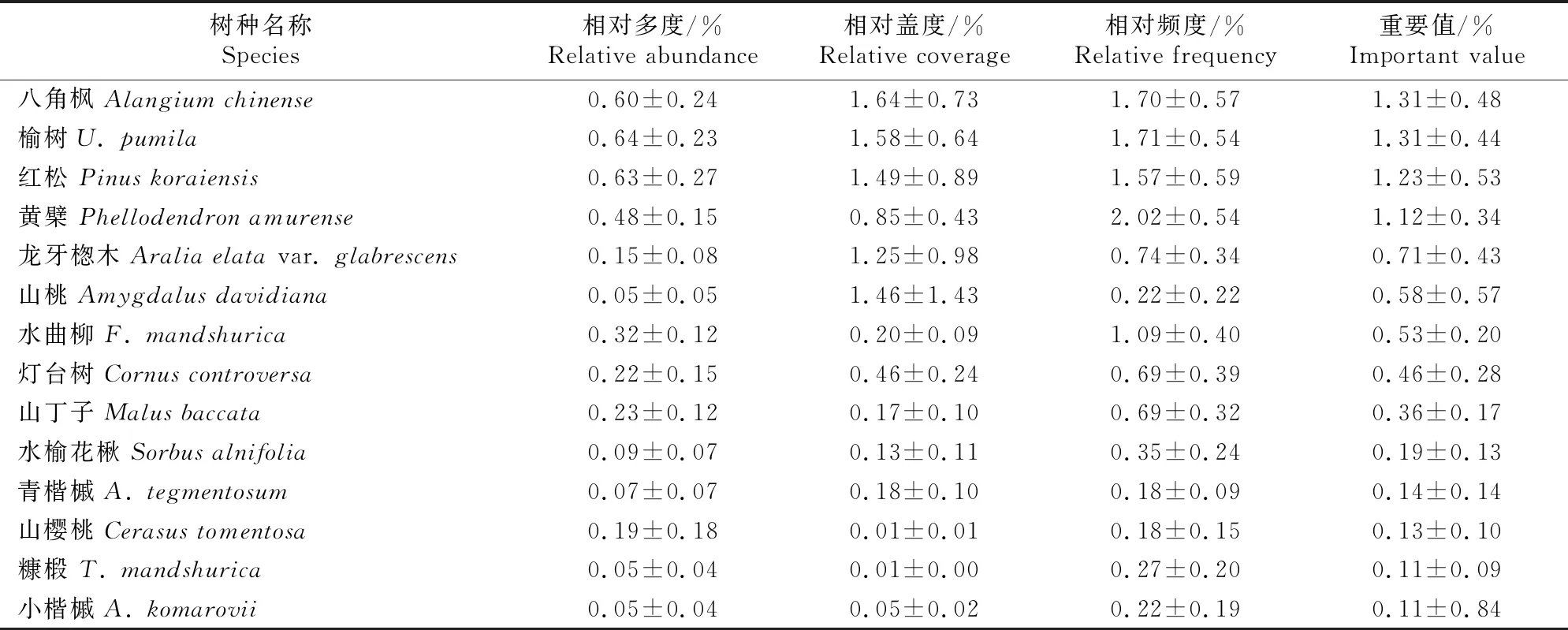
表2(续) Continued table 2
3.1.2 更新幼苗(树)的高度级分布 由图1可知,蒙古栎林乔木更新幼苗(树)高度级分布曲线呈倒“J”型,随着高度级的增大,幼苗(树)密度呈降低趋势。Ⅰ、Ⅱ级幼苗数量最多,占更新苗总数的60.20%;Ⅲ级幼苗数量锐减,仅占Ⅱ级幼苗的35.00%;而Ⅹ级幼树密度仅为Ⅰ级幼苗的1.91%。可见,蒙古栎林群落中有丰富的幼苗库,为森林群落的天然更新奠定了一定的基础,但从幼苗到幼树的发育过程中,更新苗死亡率较高,造成幼树个体数量较少,群落天然更新困难。
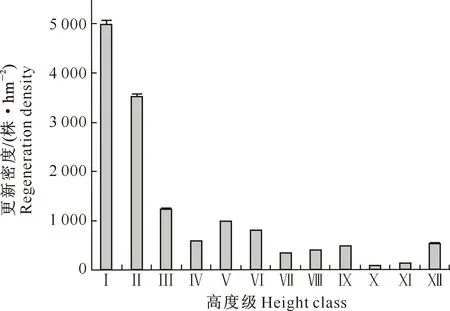
图1 辽东山区蒙古栎林乔木树种在不同高度级的更新密度
进一步分析幼苗(树)中重要值最大的5个树种的高度级分布曲线(图2)可知,蒙古栎幼苗的死亡率较高,从Ⅰ级到Ⅱ级,幼苗密度急剧减少,由2 948株/hm2锐减到290株/hm2,说明此阶段蒙古栎幼苗大量死亡。花曲柳表现为Ⅰ、Ⅱ级幼苗密度相对较大,Ⅲ级幼苗数量锐减,其他阶段趋于平稳。紫椴Ⅲ级幼苗密度最大,Ⅳ级数量锐减仅占Ⅲ级幼苗密度的13.31%。元宝槭和假色槭虽然更新幼苗的数量相对较少,但在每个高度级均有一定数量的幼苗,能够完成种群的自我更新。

图2 辽东山区蒙古栎林5个主要乔木树种在不同高度级的更新密度
3.1.3 更新方式 由图3可知,在蒙古栎次生林中,实生苗和萌生苗密度分别为12 695和290株/hm2,实生苗数量为萌生苗数量的38倍,表明更新方式主要以实生更新为主。说明在蒙古栎次生林乔木更新过程中,母树分布和结实率对林分更新具有重要的影响,萌生更新仅为实生更新的一种补偿。
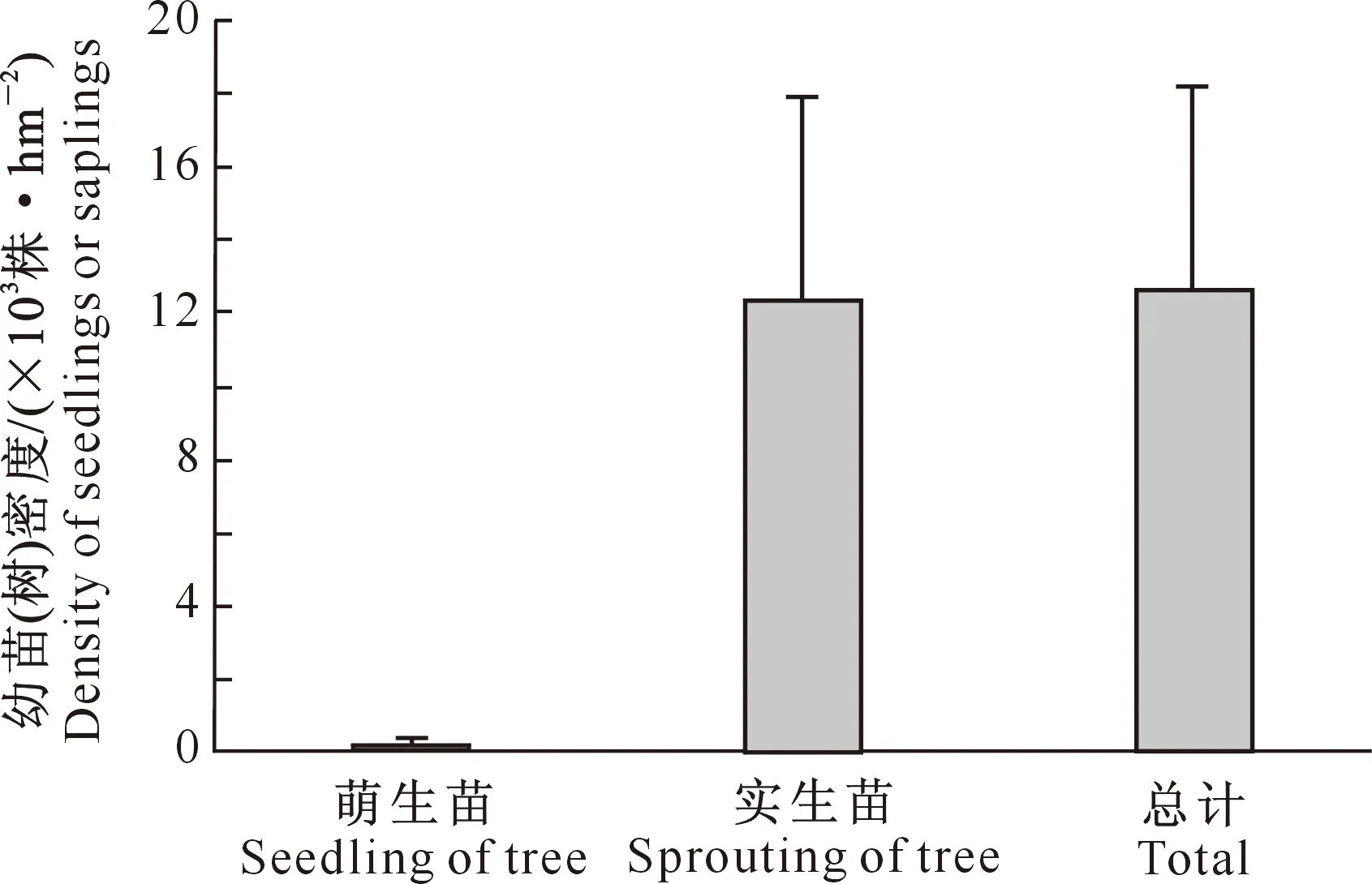
图3 辽东山区蒙古栎次生林在不同更新方式下的幼苗(树)密度
3.2 环境因子对蒙古栎次生林乔木更新的影响
分别以林下光环境、地被物和土壤理化性质等指标为自变量,以乔木幼苗、幼树密度为因变量进行Pearson相关分析,结果(表3)表明,幼苗、幼树密度对环境因子的响应不同,幼苗密度与林冠开阔度、林下总光合辐射、土壤全磷和有效磷含量呈显著正相关(P<0.05),与叶面积指数、枯落物厚度、土壤pH值、土壤全氮和碱解氮含量、土壤有机质含量呈极显著或显著负相关(P<0.01或P<0.05)。幼树密度与林冠开阔度、土壤全磷和土壤速效钾含量呈显著负相关(P<0.05),与叶面积指数、土壤含水量呈显著正相关(P<0.05),与其他指标相关性不大。

表3 辽东山区蒙古栎次生林更新乔木幼苗和幼树密度与环境因子的Pearson相关分析
为进一步明确影响蒙古栎次生林乔木幼苗、幼树密度的关键环境因子,分别以幼苗和幼树密度为因变量(y)、各环境因子为自变量(x)进行多元线性逐步回归分析,结果(表4)显示,影响幼苗、幼树密度的关键环境因子不同,幼苗密度受林冠开阔度、土壤全磷含量和土壤pH值的影响较大,模型判定系数R2=0.56;幼树密度主要受土壤全磷含量和土壤含水量的影响较大,模型判定系数为R2=0.42。

表4 辽东山区蒙古栎次生林更新乔木幼苗和幼树密度与环境因子的多元逐步回归模型
4 结论与讨论
4.1 蒙古栎次生林乔木树种更新特征
本研究表明,辽东山区蒙古栎次生林更新树种组成丰富,共有乔木树种30种,其中蒙古栎、元宝槭、花曲柳、假色槭、紫椴为更新层的优势树种,其他更新树种重要值较低,未形成明显的竞争关系。幼苗(树)高度级分布曲线呈倒“J”型,随着高度级的增大,幼苗(树)密度呈降低趋势,Ⅰ、Ⅱ级幼苗数量最多,占更新苗总数的60.20%,Ⅲ级幼苗数量锐减,仅占Ⅱ级幼苗的35.00%,而Ⅹ级幼树密度仅为Ⅰ级幼苗的1.91%。可见从幼苗到幼树的发育过程中,幼苗死亡率较高,成为更新苗进入主林层的制约因素。从更新方式来看,辽东山区蒙古栎次生林主要以实生更新为主,萌生更新为实生更新的一种补偿,这样更有利于林下更新层物种遗传多样性的维持[13]。
4.2 蒙古栎次生林乔木树种更新影响因子分析
林冠是森林与外界环境相互作用最直接和最活跃的界面[14],林冠通过影响林下光照、温度、水分等微生境,进而对更新幼苗的建立和生长产生影响。多数研究表明,林冠开阔度越大,林内透光条件越好,越有利于更新苗的建成[15-18]。本研究中,幼苗和幼树对冠层结构的适应能力不同,幼苗密度表现为随林冠开阔度的增加而增大,随叶面积指数增加而减小,幼树则表现为相反的趋势,这可能是由于蒙古栎次生林为阔叶混交林,叶片密集、透光度较差,光照相对充足的地块更有利于幼苗的建立和生长[19];然而幼苗发育到幼树的过程中,大量阳性树种死亡(幼树中一半以上为阴性或中性树种),相对于光资源,幼树对土壤资源的竞争更敏感[20],而与幼树进行资源竞争的主要为灌木,林冠开阔度的增加,促进了灌木的建立和生长,从而间接地抑制了幼树的存活和生长。
林下灌草层也是影响幼苗能否在林下成功定居的重要因素之一,大量研究表明,灌草层与更新幼苗共同竞争林下生存空间和养分资源,从而对更新苗产生一定的抑制作用[21-22]。本研究区域内由于灌木分布较少,因此仅就草本对更新幼苗的影响进行分析,结果表明草本盖度对幼苗密度影响并不显著,可能是由于调查样地中草本盖度主要集中在15%~30%,差异较小所致。本研究发现,枯落物厚度与幼苗密度呈显著负相关,这一方面可能是由于蒙古栎次生林枯落物储量高、湿度大,散落在枯落物层的种子易霉变失活,进而影响发芽;另一方面,即使种子在枯落物中萌发形成幼苗,但由于其根系不能伸入土壤,无法获得充足的养分,导致更新幼苗无法存活[2,23]。
有研究表明,土壤pH值和营养元素含量可对植物更新、生长产生直接影响[24]。Dyderski等[6]研究发现,低pH值(3.5~4.5)更有利于挪威云杉幼苗的更新;刘帅等[25]对长白山次生针阔混交林更新的研究表明,幼苗存活率与土壤pH值呈负相关。本研究也得出相似的结论,研究样地土壤pH值为4.8~6.1,相对偏酸性的土壤更有利于幼苗的更新。土壤养分含量对幼苗、幼树的影响不尽相同;幼苗阶段,土壤全氮和有机质含量对幼苗密度有极显著负影响(P<0.01),土壤全磷含量对幼苗密度有显著正影响(P<0.05);而幼树阶段,土壤全磷含量对幼树密度有显著负影响(P<0.05),这可能与植物在不同发育阶段对营养元素选择和吸收利用的差异性有关[26-27]。
多元线性逐步回归分析结果表明,影响幼苗密度的关键环境因子为林冠开阔度、土壤全磷含量和土壤pH值,影响幼树密度的关键环境因子为土壤全磷含量和土壤含水量。然而2个多元线性回归模型的决定系数均不高,说明其他因子如生物邻体等对蒙古栎次生林的更新也起到至关重要的作用。因此,在以后的研究中,应综合考虑生物因素与非生物因素对林分更新的影响,以期更深入地探寻影响辽东山区蒙古栎次生林天然更新的关键因子。
综上所述,人工促进蒙古栎林天然更新时,可采取枯落物清理、灌木割除和适度间伐相结合的经营措施,一方面为幼苗的定居和生长创造条件,另一方面可减少灌木对资源的竞争。
