16个地被菊品种的苗期抗寒性评价
2023-02-18韦淋馨何立飞张钰晗李庆卫
韦淋馨,何立飞,张钰晗,李庆卫
(1.北京林业大学 园林学院,北京 100083;2.北京市花木有限公司,北京 101309)
地被菊(Chrysanthemummorifolium)为菊科菊属多年生宿根草本花卉,是已故陈俊愉院士及其课题组利用原产我国的优良野生菊花资源反复杂交选育而诞生的具有园林应用价值的菊花新品种[1],适于各类绿地,是优良的地被植物。
地被菊具有较强的抗寒能力,至少可耐约-15℃的低温,一般品种可耐-20~-30℃的低温,少数品种可耐-35~-40℃的低温。地被菊在三北地区进行推广使用时,抗寒性是最为关键的因素。近年来,有学者在地被菊抗寒性方面进行了一定的研究,徐基艳等[2]在沈阳地区通过测定5个地被菊品系在自然低温条件下的各项生理指标,结果发现地被菊SOD酶活性随着温度的降低呈先升后降的趋势,可溶性蛋白含量、MDA含量和游离脯氨酸含量随着温度的降低呈总体增加的趋势;白永霞等[3]通过对‘紫勋章’在北京冬季自然低温条件下的观测,结果发现地被菊叶片和脚芽的SOD和CAT酶活性呈先升后降的趋势,POD活性呈先降后升的状态,叶片的可溶性糖随着温度的降低而显著增加,脯氨酸含量和可溶性蛋白总体呈下降趋势,可溶性糖含量显著增加,脚芽和根的膜透性则呈现先升后降的趋势;李晓伟等[4]对地被菊‘晚粉’脚芽在不同的冷藏温度和冷藏时间进行处理,结果表明冷藏温度为4℃时的脚芽生根率、平均生根数、平均最长根长最优,冷藏最长时间为4周。目前对于地被菊苗期抗寒性研究较少,本研究以16个地被菊良种为研究对象,于苗期对其进行低温胁迫后测定叶片相对电导率、超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量,计算苗期半致死温度,并对地被菊的抗寒指标进行综合评价,得到16个品种的抗寒性排序,从而给地被菊的推广应用提供品种选择的依据。
1 材料与方法
1.1 材料
供试材料于北京林业大学梅菊圃与大兴地被菊资源基地采集。选取16个生长良好、往年记录抗寒性优良的地被菊品种(表1)于2020年8月初扦插,进行日常田间管理。
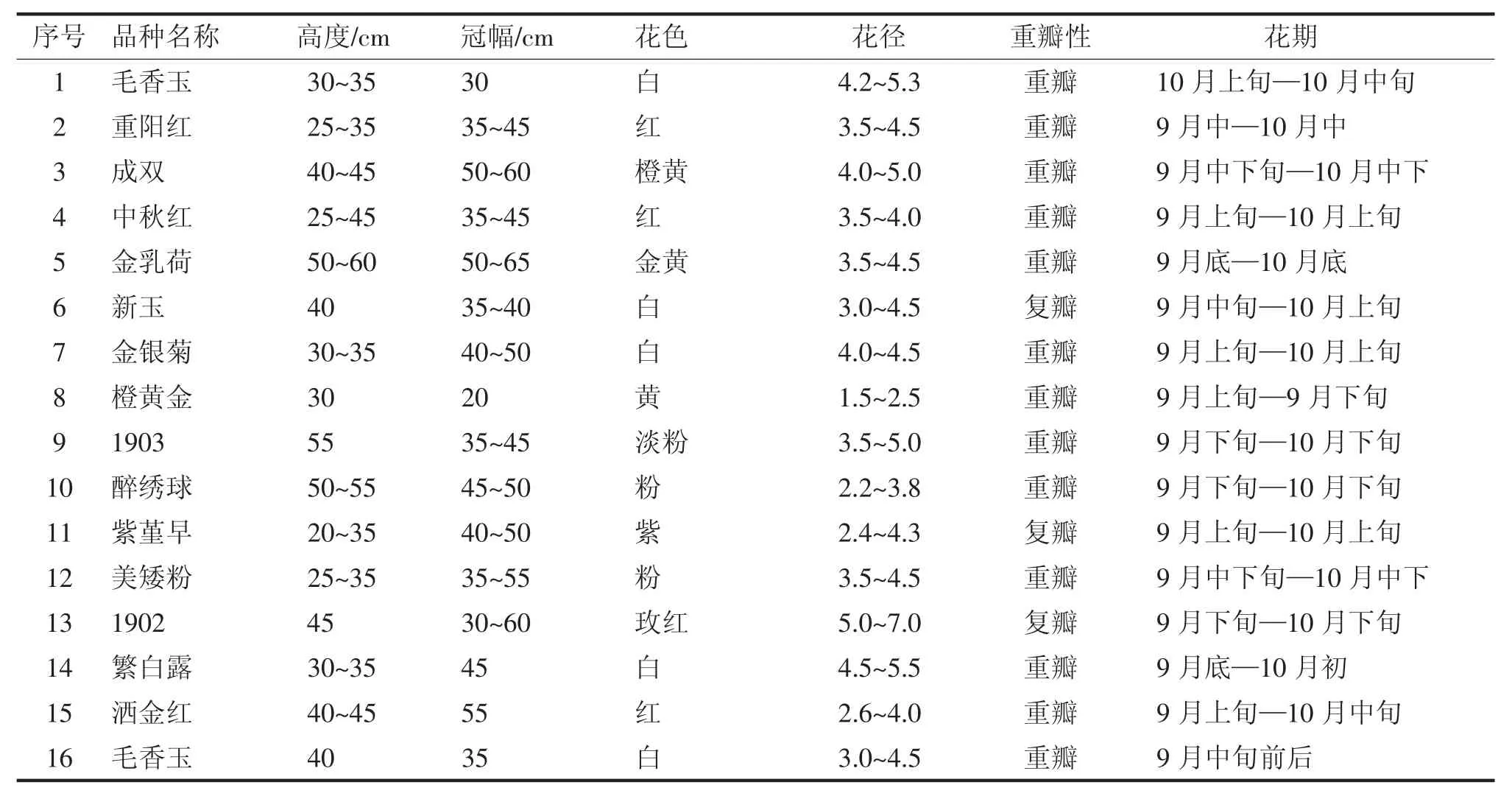
表1 参试地被菊品种田间性状观察统计
1.2 材料采集及处理
材料于11月13日采集进行低温胁迫梯度处理。参照陈煜[5]的方法,对叶片处理方式和提浸时间稍加修改。2020年11月13日采集参试的16个供试品种的植株中上部健康叶片约12 g,使用自来水冲洗干净,再用去离子水漂洗至少2次,在滤纸上吸干。将叶片分为6份,每份约2 g,为防止叶片失水,用湿纱布包好置于试管中,放置于4℃冰箱中过夜。次日在附有自动温控仪的卧式透明冰柜中进行低温处理。处理温度梯度设置为0、-4、-8、-12、-16、-20℃。供试材料在各温度下处理2 h,再匀速降温1 h,直到所有叶片处理完成。
1.3 生理指标测定
本试验采用电导仪测定叶片相对电导率;参照李合生[6]试验指导,采用氮蓝四唑(NBT)比色法测定SOD活性;使用硫代巴比妥酸(TBA)比色法测定MDA含量。
2 结果与分析
2.1 不同时间低温处理下叶片相对电导率的变化
根据16个地被菊品种叶片在不同温度处理下测得的相对电导率,使用Logistic方程进行拟合,求得各地被菊品种半致死温度(LT50)(表2)。对16个品种进行Logistic方程计算,并绘制4个品种的拟合曲线(图1)。试验结果表明,参试品种的相对电导率随温度的下降而上升,呈现明显的S型曲线,图中5个点对应的处理温度分别为0、-4、-8、-12、-16、-20℃。电解质渗出速率由慢到快,之后趋于稳定。S型曲线的拐点处理温度即为地被菊各品种的半致死温度。
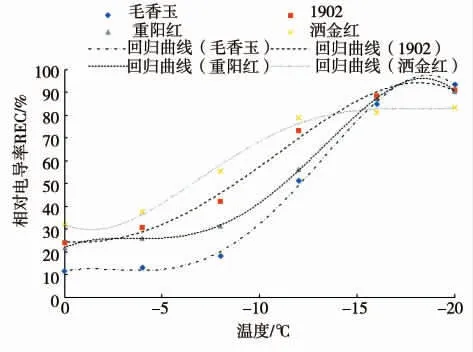
图1 4个地被菊地被菊品种相对电导率随温度变化的Logitic曲线

表2 16个地被菊品种相对电导率回归方程及半致死温度(LT50)
2.2 不同时间低温处理下叶片SOD活性的变化
不同低温处理后,各地被菊品种叶片SOD活性的影响见图2。大部分地被菊品种在0℃的条件下SOD含量相差不大,但是经过低温胁迫处理后,SOD活性出现了3种不同的变化情况。品种‘1902’、‘1903’、‘橙黄金’、‘毛香玉’、‘中秋白’、‘重阳红’和‘醉绣球’的SOD活性随着温度的降低呈降低状态;品种‘繁白露’、‘金乳荷’、‘金银菊’、‘美矮粉’、‘洒金红’和‘中秋红’的SOD活性随着温度的降低呈先升后降的趋势;品种‘成双’、‘新玉’和‘紫堇早’的SOD活性随着温度的降低呈现“升—降—升”的变化情况。
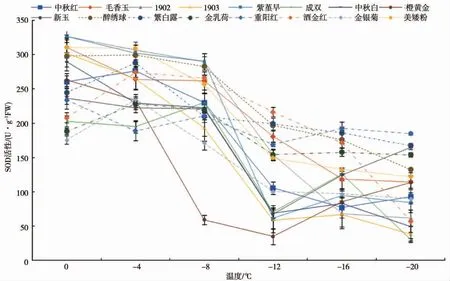
图2 16个地被菊地被菊品种叶片SOD活性随温度变化的曲线
将各温度梯度下测得的SOD活性变化率与抗寒性进行相关性分析,可得结果见表3。相关分析表明,6个温度处理下SOD活性的变化率与抗寒性指标的相关程度在-16~-20℃之间SOD活性相关系数较大,相关系数为0.209,但未达到显著水平,而-8~-12℃和-12~-16℃之间两者相关性呈显著水平。这说明地被菊SOD活性与抗寒性的关系在温度更低的情况下表现更为明显。抗寒性较好的品种在降温中期SOD活性下降较快,当温度继续降低时,抗寒性强的品种又展现出较强的抵抗能力。

表3 SOD活性变化率与抗寒性的相关分析
2.3 不同时间低温处理下叶片MDA含量的变化
不同低温胁迫处理后各品种叶片MDA含量的测定结果见图3。16个地被菊品种在低温胁迫下MDA含量的变化规律基本一致,呈现先降低后升高的趋势。8个品种在-12℃时MDA浓度最低,8个品种在-16℃时MDA浓度最低。

图3 16个地被菊地被菊品种叶片MDA活性随温度变化的曲线
将各温度梯度下测得的地被菊叶片MDA含量的变化率与抗寒性进行相关性分析,可得结果见表4。相关分析表明,在6个低温胁迫处理中,-16~-20℃之间的MDA变化率与抗寒性指标相关系数为最高,达到0.410。其余低温胁迫条件下测定的MDA含量变化率与抗寒性指标相关系数均比较低。由此初步推断,地被菊MDA含量与抗寒性的关系在温度更低的时候表现更为显著。原因可能是在-16~-20℃处理下已经对植物造成较大伤害,使得MDA含量上升。

表4 MDA含量变化率与抗寒性的相关分析
将16个品种的MDA含量变化率与抗寒指标进行聚类分析,当阈值取聚类距离2.418~6.369之间,可计算出各品种间的聚类距离(表5)。由表6可知,将16个品种分为4个抗寒类型:抗寒性极强,抗寒性较强,抗寒性较差和抗寒性差。‘毛香玉’为抗寒性极强的品种,‘重阳红’、‘金银菊’、‘成双’、‘金乳荷’、‘新玉’、‘1902’、‘1903’、‘紫堇早’和‘橙黄金’为抗寒性较强的品种,‘洒金红’、‘醉绣球’、‘中秋红’、‘美矮粉’和‘繁白露’为抗寒性较差的品种,‘中秋白’为抗寒性差的品种。
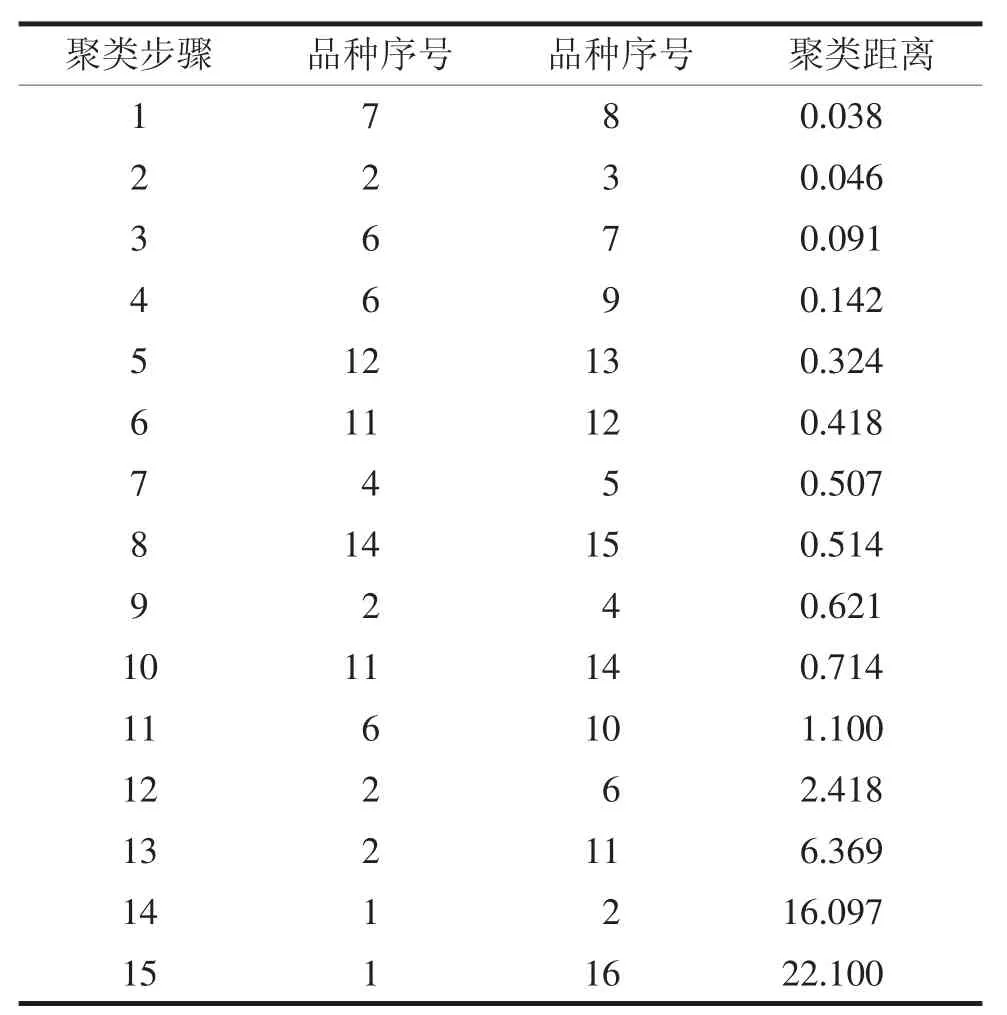
表5 地被菊叶片MDA含量变化率与抗寒性联合聚类分析
将聚类分析所得的抗寒类型与以-8~-12℃测定的MDA含量变化率的平均指标值进行比较,抗寒性强的品种在-8~-12℃的MDA含量降幅较小,抗寒性较差的品种在降温前期降幅较大,达到更低温度时,降幅已基本稳定。
2.4 地被菊苗期抗寒性评价
使用隶属函数分析法[7]对地被菊苗期抗寒性进行综合评价(表7)。通过隶属函数分析,结果表明,16个地被菊品种中,苗期抗寒性顺序为:‘毛香玉’>‘重阳红’>‘成双’>‘中秋红’>‘金乳荷’>‘新玉’>‘金银菊’>‘橙黄金’>‘1903’>‘醉绣球’>‘紫堇早’>‘美矮粉’>‘1902’>‘繁白露’>‘洒金红’>‘中秋白’。结合植物形态方面的观测,‘毛香玉’与其他品种相比叶片正反面均匀密布了一层柔毛,且叶片较厚;‘重阳红’和‘成双’叶片相比其他品种厚度较厚。植物低温下的指标与叶片的结构有着密切的联系,叶肉发达程度越高,栅栏组织与海绵组织的比值越高,品种的耐寒能力越高[8]。

表7 16个地被菊品种抗寒性综合评定指数与排序
3 讨论与结论
3.1 不同地被菊品种的苗期半致死温度
本研究在对地被菊叶片的相对电导率测定方法上进行Logistic方程拟合,通过求出S形曲线的拐点温度能够较为准确地估算出植物的半致死温度[9],初步评价了16个地被菊品种苗期的抗寒性,可初步推断为‘毛香玉’抗寒能力最强。
由于材料营养生长时间较短,且植株未经历低温锻炼,展现出的抗寒能力与前人对地被菊抗寒能力的研究存在差别。由于不同年龄阶段耐寒性差异不同,地被菊苗期抗寒性较弱,正常越冬的扦插苗与春季繁殖的扦插苗抗寒表现要强于夏季的扦插苗。因此,在寒冷地区进行地被菊栽培时需要选择合适的扦插时间,本研究中地被菊扦插时间较晚,但进入短日照环境后依然可以正常开花,开花并不整齐,所以扦插时间要早于8月份,以提高植株的抗寒性。在观测时,笔者发现夏季扦插的地被菊扦插苗根系幼嫩且数量少,由此可知地被菊夏季扦插苗抗逆性较差。经过与王晓倩[10]的研究的对比和大田记录的数据对比,笔者发现晚花品种抗寒性一般要强于早花品种,例如‘毛香玉’花期最晚,抗寒性最强。
3.2 不同时间低温处理对叶片SOD活性的影响
低温胁迫导致植物体内活性氧增加,分解消耗活性氧的能力下降,导致活性氧含量增高,当活性氧含量增多突破阈值时,膜系统将受到活性氧的攻击而产生的损害,SOD是第一种清除活性氧的酶,通过SOD的参与以减缓低温胁迫对生物膜的伤害。
本研究发现,随着胁迫温度的持续降低,SOD活性在不同品种间的变化差异较大,在0~8℃条件下,大部分品种SOD活性呈下降状态,但在温度继续降低时SOD活性的总体趋势为下降,但是抗寒性强的品种在低温胁迫后期SOD活性下降趋势要显著低于抗寒性差的品种,这与全英杰等[11]的研究结果一致。
3.3 不同时间低温处理对叶片MDA含量的影响
研究表明,植物在胁迫环境或衰老时,细胞内自由基代谢失衡,自由基数量增加,引发质膜过氧化作用产生MDA,进而破坏植物体内多种正常活动[12]。结果表明,地被菊叶MDA含量与低温胁迫呈显著正相关,但与品种间相关性不强,不宜单独用来评价地被菊叶的抗寒性,应结合其他指标进行综合评价。
3.4 地被菊苗期抗寒性评价
在大多数的植物抗寒研究当中,对于指标的记录不仅限于采用形态、生理生化、代谢等单一的指标来测定植物的抗寒能力,应当克服单一指标测定植株抗寒能力的局限性,利用多个指标综合评价植物的抗寒能力,测定更多指标如:叶宽、叶厚、叶面积、植株株高冠幅比、组织含水量、渗透调节物质和代谢活动等。在田间观测后记录表型形状,并采用主成分分析法、隶属函数法、分级比较法、回归分析和聚类分析等分析方法。结合前人制定的抗寒评价体系,可以更加全面地对菊花抗寒机理进行分析。
3.5 结论
本研究通过在不同低温胁迫下进行叶片的相对电导率测定,使用Logistic方程拟合计算地被菊苗期半致死温度LT50,得知‘毛香玉’半致死温度为最低,为-10.66℃,与前人对地被菊的抗寒性差距较大,由此可知地被菊苗期抗寒性弱,且地被菊不同年龄阶段的抗寒性差异较大。通过在不同低温胁迫下的测定叶片SOD酶活性和MDA含量,SOD活性在不同品种间的变化差异较大,大部分品种SOD活性总体趋势为下降,但是抗寒性强的地被菊品种在低温胁迫后期SOD活性下降趋势要明显低于抗寒性差的品种。地被菊叶片随着的MDA含量与低温胁迫基本呈正相关,MDA的含量与地被菊品种相关性不大,因而MDA含量不适合单独用于评价地被菊的抗寒性,应该与其他指标进行综合评价。综合前人的研究方法,本研究采用隶属函数法对地被菊的抗寒指标进行综合评价,对参试品种进行了抗寒性排序。
本研究所得结论可为之后地被菊抗寒试验提供一定的借鉴意义,尤其是苗期展现出较强抗寒能力的品种可以继续深入研究,为区域试验和市场化推广做出一定的基础贡献。
