猪粪及其生物堆肥对小麦幼苗生长和土壤细菌群落结构的影响
2023-02-16邹文萱罗慧赵境怡岳中辉刘四义梁爱珍王新军束长龙韩丽丽沈菊培
邹文萱,罗慧,赵境怡,岳中辉,刘四义,梁爱珍,王新军,束长龙,韩丽丽,沈菊培
(1.中国科学院生态环境研究中心,北京 100085;2.中国科学院大学,北京 100049;3.哈尔滨师范大学生命科学与技术学院,哈尔滨 150525;4.中国科学院黑土区农业生态重点实验室,中国科学院东北地理与农业生态研究所,长春 130102;5.交通运输部科学研究院,北京 100029;6.植物病虫害生物学国家重点实验室,中国农业科学院植物保护研究所,北京 100193;7.福建师范大学地理科学学院,福州 350007)
随着我国畜牧业规模化和集约化的迅速发展,畜禽养殖带来的环境污染问题日益突出。据我国畜牧总站的调查显示,全国每年畜禽粪污的排放量约38亿t[1]。畜禽粪肥中含有丰富的有机质及植物生长所需的氮、磷、钾等营养元素[2]。将畜禽粪肥合理还田进行资源化利用不仅可以大大降低化肥施用量、增加土壤有机碳积累、提高土壤肥力,而且有助于消除畜禽养殖带来的污染问题[3]。然而,禽畜粪肥如果施用不当则可能通过径流或淋溶造成地表水体富营养化和地下水污染,进而影响环境和人体健康[4]。因此,开展畜禽粪肥的资源化利用研究对指导农业生产、降低环境污染、实现资源循环利用具有重要意义。
自20 世纪80 年代以来,我国采用的畜禽粪便资源化利用方式包括肥料化、能源化和饲料化[5-7],其中前两者是主要利用方向。肥料化是利用畜禽粪便制作有机肥的过程,包括物理、化学和生物等方式。生物堆肥具有能耗小、二次污染物少等优点,是实现畜禽粪便肥料化、降低环境污染的有效生物降解技术[7]。例如,利用蚯蚓吞食过腹消化和微生物的分解作用对畜禽废弃物进行降解是一种应用较广、符合可持续发展的生物处理法[8-9]。黑水虻由于繁殖迅速,饲养操作简单,在生产中易于管理,其作为一种资源性昆虫具有一定的养殖前景与应用价值[10]。白星花金龟(Protaetia brevitarsis)幼虫可取食秸秆、畜禽废弃物,并将其转化为腐植酸含量较高的有机肥[11],在畜禽粪便资源化利用方面具有较好的应用前景。金龟幼虫肠道是一个多变的系统,随着取食、消化和排泄等活动而不断变化,在脂肪和碳水化合物吸收利用过程中起着重要作用,被称为有机废弃物资源加工的“生物反应器”[12]。
畜禽粪便生物堆肥处理在农业生产中已展现较大的潜力,对土壤生态系统也产生了显著影响。如王先哲[13]在农田土壤中施用了经黑水虻堆肥的虻粪,结果发现土壤中细菌群落多样性和酸杆菌门的相对丰度降低,而细菌群落的均匀度增加。Vivas 等[14]通过新鲜橄榄废渣及羊粪的混合物进行蚯蚓堆肥,结果表明相比原粪肥,蚓粪显著增加了细菌的数量及多样性。目前关于畜禽粪肥施用对土壤-植物-微生物体系的影响研究已有较多,但白星花金龟幼虫生物堆肥对土壤环境和作物生长的影响及其微生物作用机制鲜有报道。因此,本研究通过小麦盆栽试验,探究猪粪及其生物堆肥得到的虫粪处理后小麦栽培60 d 的生理特性、氮肥利用率及土壤细菌群落结构的变化,并分析小麦生长的主要影响因子,探讨土壤理化性质、小麦幼苗的生长状况与土壤细菌群落的相关性,以期为畜禽粪便资源化的农业效应评估提供参考,同时为探究畜禽粪肥还田的施用量及微生物作用机制提供理论依据。
1 材料与方法
1.1 试验材料
供试土壤采自北京市延庆区(40°29′N,115°56′E)吴庄村旱柳地表层土壤(0~20 cm),土壤类型为潮土,容重为1.31 g·cm-3,过2 mm 筛后风干、混匀备用。供试的小麦种子为北京研禾玉科技发展有限公司的“轮选987”品种,生育期258 d。供试有机肥包括猪粪和虫粪。
供试有机肥:在湖南农业大学动物科学技术学院养殖基地选取1 头公猪,饲喂玉米籽粒(来自中国科学院东北地理与农业生态研究所)和其他配料制成的粉状饲料7 d,收集4~7 d的粪便,混匀风干后过筛,将其作为供试猪粪,其总碳和总氮含量分别为361.2 g·kg-1和36.9 g·kg-1。从中国农业科学院获取3 龄的白星花金龟(Protaetia brevitarsis)幼虫,饲喂上述猪粪,7 d 后收集新鲜的颗粒状虫粪,风干后过2 mm 筛,将其作为供试虫粪,其总碳和总氮含量分别为376.9 g·kg-1和31.3 g·kg-1。
1.2 试验设计
采用盆栽法进行试验。选取12 个内径为18.5 cm、高为7.5 cm 的花盆,每盆装入土壤1 400 g。试验共设置3 个处理,分别为未施肥处理(CK)、虫粪处理(LF)以及猪粪处理(SM),每个处理4 次重复。CK 处理中无氮肥输入,添加的磷肥和钾肥为过磷酸钙和氯化钾,施入量分别为247.2 mg·kg-1和158.5 mg·kg-1。LF和SM 处理按照含氮量200 mg·kg-1等氮量添加,添加的虫粪和猪粪量分别为639.1 mg·kg-1和542.1 mg·kg-1,混匀后将土壤含水量调节至田间持水量的50%。
选择籽粒饱满、大小均匀的小麦种子,经30%H2O2消毒10 min后用蒸馏水反复冲洗,滤纸吸干后备用。每盆播种上述小麦种子25 粒后用100 g 土覆盖,出苗后保留20 株,在25 ℃、12 h·d-1光照条件下种植60 d 后进行植株和根际土壤样品采集。采集时先将小麦幼苗地上部分剪切,然后测定鲜质量,放入烘箱内105 ℃杀青60 min,在75 ℃下烘至恒质量,研磨过0.15 mm 筛后备用。土壤挑去根系后分为三部分:第一部分存放于-80 ℃冰箱用于DNA的提取,第二部分于4 ℃保存用于土壤有效态氮含量的测定,第三部分风干过筛后进行土壤总碳(TC)、总氮(TN)的测定。
1.3 测定方法
1.3.1 土壤性质的测定
土壤TC 和TN 含量采用元素分析仪(Vario ELⅢ-Elenemtar,德国)测定;土壤铵态氮(-N)和硝态氮(-N)经1 mol·L-1的KCl 溶液浸提后用连续流动分析仪(AA3,SEAL,德国)测定;土壤可溶性有机碳(DOC)和可溶性有机氮(DON)用0.5 mol·L-1的K2SO4浸提,并用总有机碳分析仪(TOC-VCPH,Shimadzu,日本)测定。
1.3.2 小麦幼苗生理指标的测定
采用尺量法测定小麦五叶期、六叶期的株高和近根叶组的第5 片叶的叶面积;小麦植株的TC 和TN含量采用元素分析仪测定;五叶期和六叶期的小麦叶片叶绿素SPAD值用叶绿素仪(SPAD-502 Plus,Konica Minolta,日本)测定。作物氮肥利用率采用差减法计算。
1.3.3 细菌16S rRNA高通量测序
利用土壤DNA 试剂盒(DNeasy®Power Soil®Kit,QIAGEN,德国)提取0.35 g 冷冻土壤,用NanoDrop ND-1000 分光光度计(NanoDrop,Thermo ScientificTM,美国)测定提取DNA 的浓度和质量。使用细菌16S rRNA基因V4~V5区域的通用引物338F(ACTCCTACGGGAGGCAGCA)和806R(GGACTACHVGGGTWTCTAAT)进行PCR扩增,PCR反应体系(20 μL):5×Fast-Pfu 缓冲液4 μL,2.5 mmol·L-1dNTPs 2 μL,浓度5 μmol·L-1的前后引物各0.8 μL,FastPfu 聚合酶0.4 μL和DNA模板10 ng。在ABI GeneAmp®9700 PCR扩增仪上进行扩增,用凝胶电泳检测扩增产物。PCR产物用AxyPrep DNA 凝胶回收试剂盒(Axygen Biosciences公司)进行纯化,随后送上海美吉生物医药科技有限公司利用Illumina MiSeq 平台进行末端配对(2×250 bp)测序分析。
通过Quantitative Insight into Microbial Ecology(QIIME.V1.9.1)平台[15]对所测序列进行分析。在USEARCH[16]中将双端序列拼接成单条序列,并对该序列进行质控(最大期望错误率0.5)及去冗余分析。在USEARCH 中使用UNOISE2[17]挑选正确的生物序列(Zero-radius Operational Taxonomic Units,ZOTUs)。在QIIME 中使用BLAST 算法将这些代表性序列与SILVA 数据库进行比对,共获得638 981 条高质量序列,按100%相似性挑选得到6 030 个代表ZOTUs。最后通过QIIME计算细菌α多样性和β多样性。
1.4 数据处理
利用Excel 2013 对数据进行整理、汇总,利用SPSS 软件(SPSS Statistics 24)进行单因素方差分析(ANOVA)、显著性检验(Duncan,P<0.05)和多元回归分析,采用R(Version 3.4.3)软件[18]进行冗余分析、主成分分析和差异分析。
2 结果与分析
2.1 土壤性质和小麦生长状况
不同施肥处理下小麦栽培60 d 后土壤性质如表1 所示。与CK 相比,施肥处理LF 和SM 显著提升了土壤pH值(P<0.05)。LF和SM处理的TC、TN、-N、-N、DON 含量均显著高于CK 处理(P<0.05),而LF和SM 处理之间差异不显著(P>0.05)。LF和SM 处理的DOC 含量显著高于CK(P<0.05),且LF 处理显著高于SM处理(P<0.05)。

表1 不同施肥处理的土壤性质Table 1 Soil properties in different treatments
不同施肥处理下小麦生理指标变化如图1 所示。LF 和SM 处理苗干质量、叶绿素SPAD 值均显著高于CK 处理(P<0.05),而小麦苗碳氮比显著低于CK 处理(P<0.05),且LF 和SM 处理间差异不显著。SM 处理氮肥利用率显著高于LF处理。另外,仅SM 处理的株高显著高于CK 处理(P<0.05),LF 和SM 处理的叶面积在五叶和六叶期与CK 处理均无显著差异(P>0.05)。
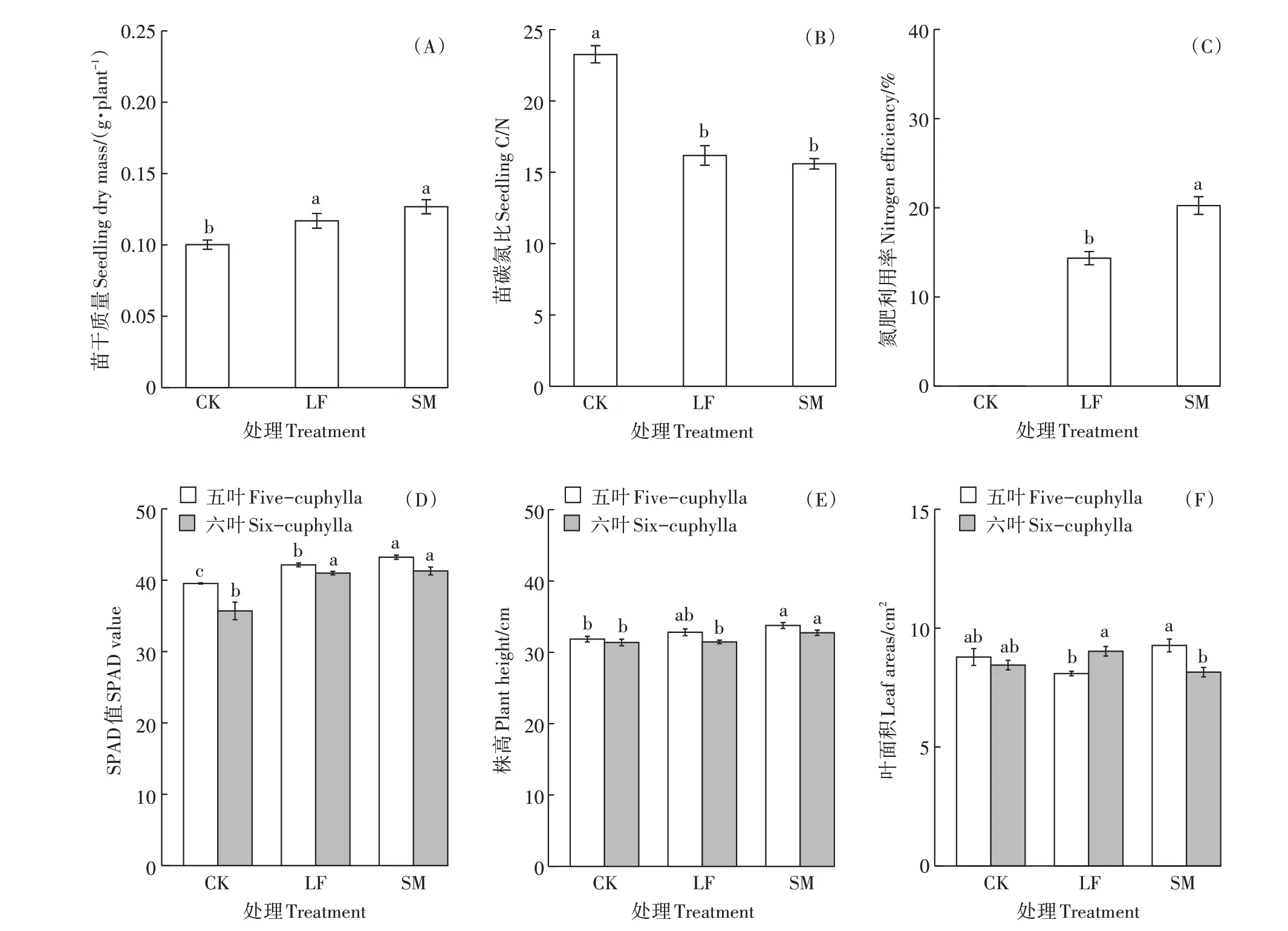
图1 不同施肥处理下小麦生理指标特征Figure 1 Wheat growth characteristics in different treatments
2.2 土壤细菌多样性
香农指数(Shannon index)可以指示土壤细菌的多样性。基于对土壤细菌测序ZOTU 数据的分析发现,不同处理间土壤细菌香农多样性指数无显著差异(P>0.05,图2A)。细菌群落组成结果表明,丰度前10位的土壤细菌类群主要有放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、拟杆菌门(Bacteroidetes)、芽单胞菌门(Gemmatimonadetes)、Patescibacteria 菌门、厚壁菌门(Firmicutes)、棒状杆菌门(Rokubacteria)、疣微菌门(Verrucomicrobia)等。其中,土壤中丰度最高的类群为放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)和绿弯菌门(Chloroflexi),分别占土壤细菌的32%~34%、24%~26%、14%~17%和12%~14%(图2B)。
细菌类群相对丰度差异性分析的结果表明,不同粪肥施用对细菌类群产生了显著影响。例如,LF 和SM 处理中放线菌门的相对丰度比CK 处理分别提高了2.85、1.11 个百分点,而酸杆菌门的相对丰度在LF处理中最低(平均14%),在CK 处理中最高(平均17%)。LF 和SM 处理中绿弯菌门的相对丰度比CK处理分别降低了1.63、1.58个百分点。基于Bray-Curtis 距离算法对细菌ZOTU 数据进行PCoA 主成分分析(图2C),其中第一、二排序轴分别解释总变异的12.2%和11.4%,SM 处理与其他处理相距较远,具有较低的细菌群落相似性,CK 和LF 处理细菌群落结构分布部分重合,说明具有较高的细菌群落相似性。

图2 不同处理土壤细菌多样性和群落结构特征Figure 2 Soil bacterial diversity and community structure in different treatments
研究采用随机森林模型平均基尼指数减少值(Mean Decrease Gini)评估不同微生物类群的重要性,即平均基尼指数减少值越大,代表该微生物类群越重要。如图2D 所示,在门水平上检测到3 种显著差异的土壤细菌门类,包括拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)和硝化螺菌门(Nitrospirae)。其中,SM 处理拟杆菌门的相对丰度为5.8%,显著高于CK(2.9%)和LF(3.7%)处理(P<0.05)。SM 处理厚壁菌门和硝化螺菌门相对丰度分别为1.0%和0.2%,LF 处理厚壁菌门和硝化螺菌门相对丰度分别为1.2%和0.3%,均显著高于CK处理(0.6%和0.2%)。
2.3 土壤性质、细菌群落与小麦生理指标的相关性
将土壤性质和基于ZOTU 信息的细菌群落结构进行冗余分析(图3),结果表明其中7 个土壤因子共解释了细菌群落总变异的34.5%,RDA 前两个排序轴分别解释了18.5%和16.0%的群落变异。由图3 可知,影响土壤细菌群落的环境因子主要为-N、DOC、DON、TC、-N 和TN。皮尔森相关性分析表明,小麦幼苗的生长指标与土壤养分含量显著相关(表2),其中-N、NO-3-N、DOC 和DON 与苗干质量、SPAD值和氮肥利用率均呈显著正相关,而与苗碳氮比呈显著负相关(P<0.01)。TC、-N、DON 与株高呈显著正相关(P<0.05)。

图3 土壤细菌群落结构与土壤性质的冗余分析Figure 3 Redundancy analysis(RDA)of soil bacterial community structure and soil properties

表2 小麦苗期生长指标和土壤性质的相关性分析Table 2 Pearson correlation analysis of wheat seedling growth characteristics and soil properties
基于随机森林模型筛选出3 种具有显著差异的土壤细菌门类群,将其与土壤因子进行皮尔森相关性分析(表3)。结果表明土壤TC、TN、-N 和DON 含量与拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)和硝化螺菌门(Nitrospirae)均呈显著正相关。多元回归结果(表4)显示,pH、TC、-N、-N、细菌群落及多样性等能够共同解释小麦苗干质量变异的88.9%(r=0.943,P=0.027),显著性检验结果表明-N含量对苗干质量的影响具有统计学意义(P<0.05)。

表3 显著差异细菌门类群相对丰度和土壤性质的相关性分析Table 3 Pearson correlation analysis among the relative abundance of most significant bacterial phyla and soil properties

表4 小麦幼苗干质量与土壤性质及细菌群落结构之间的多元回归分析Table 4 Multiple regression analysis of soil properties,bacterial community structure and wheat seedling dry mass
3 讨论
3.1 不同有机肥处理对小麦生长和氮肥利用率的影响
本研究发现施用猪粪和生物转化后虫粪对土壤养分和小麦幼苗的生长均具有一定的促进作用。土壤的养分含量(总碳、总氮、铵态氮、硝态氮及可溶性有机碳氮)和小麦生长指标(株高、叶面积、苗干质量及SPAD 值)均增加,且施用等氮量猪粪与虫粪的效果一致,表明畜禽粪肥经生物转化后对土壤和作物的生长同样具有重要作用,可作为有效转化畜禽废弃物资源的可能途径。这与田小燕等[19]的研究结果一致,经过生物堆肥后的虫粪对土壤肥力和植物生长具有促进作用。作物的株高、叶面积和苗干质量等生长指标能够反映作物的生长状况和养分吸收情况,SPAD值表征了作物叶片中叶绿素的相对含量,反映作物的光合能力和长势[20]。等氮量虫粪和猪粪处理增加了土壤碳氮养分,满足了小麦生长的养分需求,进而同等程度地增加了小麦地上部生物量的积累以及叶绿素含量,说明畜禽粪肥经生物堆肥处理后能够促进小麦生长[21-22]。
氮肥利用率能够表征施用的氮肥被作物吸收、在土壤中残留及环境损失的实际情况,是用于评价氮肥施用效果的重要指标[23]。本研究发现施用猪粪处理土壤中小麦氮肥利用率高于生物堆肥后虫粪处理的氮肥利用率。这可能是由于后者经白星花金龟幼虫处理后更多的氮源成为腐植酸的主要成分[24],在保证植物生长的同时,又为后续生长提供了稳定有效的养分资源库[25]。Li 等[24]研究发现以玉米秸秆和木屑饲喂白星花金龟幼虫,提取的虫粪无植物毒性,且腐植酸含量达到24.37%和14.46%。因此,今后还需通过观测整个小麦生长周期进一步认识生物堆肥氮素的有效性。
3.2 不同有机肥处理对土壤细菌群落的影响
土壤微生物群落结构和多样性是衡量土壤质量的重要指标[26]。细菌作为土壤中最为丰富的一类微生物,对维持土壤生态系统具有重要意义。本研究表明,与施用猪粪相比,施用虫粪对土壤细菌α 多样性和群落结构均无显著影响,说明虫粪携带的微生物没有改变土壤细菌群落,这与Wang 等[27]研究的家蝇幼虫生物堆肥的结果一致。有学者通过溯源分析的方法,发现家蝇幼虫生物堆肥第1 天仅有1.4%的细菌源于幼虫肠道,而在处理的第6 天,有近20%的细菌来源于幼虫肠道,说明堆肥处理过程中肠道微生物的组成显著影响了畜禽粪便微生物组成[27]。本研究也推测,白星花金龟幼虫生物降解过程中肠道微生物发挥了巨大作用。白星花金龟幼虫的肠道主要分为三部分:前肠、中肠和后肠[11]。中肠是一个pH 高达11的碱性环境,能消解大部分粪源性微生物;而后肠含有大量消化有机化合物的蛋白酶[12]。经肠道处理产生的虫粪微生物结构相对稳定,且大部分畜禽携带的致病菌被去除[28]。Zhao 等[29]也发现,猪粪和鸡粪经白星花金龟幼虫生物处理后,微生物之间的关系更加稳定,土壤抗生素抗性基因的传播风险降低。基于这一结论,笔者认为白星花金龟幼虫可以将畜禽有机废弃物有效转化,产生的虫粪具有较低的生态风险。
尽管生物堆肥对土壤细菌α 多样性和群落结构的影响不显著,但对部分细菌类群的相对丰度产生了显著影响,如拟杆菌门、厚壁菌门和硝化螺菌门,这与Wolters 等[30]的研究结果类似。施加粪肥增加了厚壁菌门的相对丰度,这可能是由于厚壁菌门更能适应富碳的环境[31],可以参与有机碳等复杂有机物的降解,能够为作物生长提供养分。硝化螺菌门是土壤中主要的硝化细菌,而小麦是喜硝作物[32],因此施加粪肥可提高土壤中硝化螺菌门,有利于促进土壤中-N向-N 的转化,进而为小麦提供有效养分。而施用虫粪的土壤中拟杆菌门的相对丰度显著低于施用猪粪处理。拟杆菌门主要来自畜禽肠道,往往携带一些致病菌等[29],对土壤环境造成潜在的生态风险。本研究发现生物堆肥处理显著降低拟杆菌门的相对丰度,这与Wang等[27]对家蝇幼虫的研究结果相似,但与Ao 等[33]对黑水虻的研究结果相反,原因可能是不同腐食性动物肠道微生物组成及其与外源畜禽微生物作用方式不同有关。这一结果也再次证明白星花金龟幼虫在畜禽有机废弃物转化中的潜在价值。
4 结论
(1)经白星花金龟幼虫转化后的虫粪和等氮量猪粪施用均显著提高了土壤养分(TC、TN、-N、-N、DOC、DON)含量,促进了小麦幼苗的生长,而施用虫粪处理的小麦氮肥利用率低于施用猪粪处理。
(3)与施用猪粪相比,施用其生物堆肥产物对土壤细菌群落结构和多样性的影响不显著,这对维持土壤生态功能具有重要的参考价值。
致谢:感谢湖南农业大学沈维军教授在样品制备过程中的帮助,同时感谢中国科学院生态环境研究中心张丽梅研究员对论文工作的指导。
