天敌昆虫蠋蝽线粒体基因组蛋白编码基因分析
2023-02-16丁晓飞魏久锋高晓云刘迎香
丁晓飞,魏久锋,高晓云,刘迎香,陈 超,赵 清
(山西农业大学 植物保护学院,山西 太谷 030801)
蠋蝽(Arma custosFabricius)又名蠋敌,隶属于半翅目(Hemiptera)蝽科(Pentatomidae)益蝽亚科(Asopinae)蠋蝽属,在我国各地区广泛分布,其适应能力较强[1]。蠋蝽作为农林业领域上的一种重要的捕食性天敌昆虫,以鞘翅目和鳞翅目等多种害虫的幼虫为食[2]。研究表明,蠋蝽对草地贪夜蛾(Spodoptera frugiperda)、斜纹夜蛾(Spodoptera litura)以及小菜蛾(Plutella xylostella)等鳞翅目害虫的幼虫具有较好的防治效果[3]。
昆虫线粒体基因组是一个长15~20 kb的双链共价闭合的环状DNA分子,共37个基因,包括13个蛋白编码基因(PCGs)、22个转运RNA基因(tRNAs)、2个核糖体RNA基因(rRNAs)和1个控制区[4-5]。其中,13个蛋白编码基因(ATP6、ATP8、COX1、COX2、COX3、ND1、ND2、ND3、ND4、ND4L、ND5、ND6和Cytb)的蛋白质产物对线粒体氧化磷酸化过程至关重要[6]。由于线粒体基因组具有大量的遗传以及各种系统进化发育信息,被广泛地应用于物种鉴定、系统进化发育、谱系生物地理学和基因组水平分子进化等领域的研究[7-9]。已有研究很多是基于单个基因或部分基因对物种进行系统发育分析,尤其是COX1基因由于分子量最大、功能结构域最保守,被广泛应用于不同分类阶元层次上系统进化分析[10-11];COX2、ND2和ND5基因进化速率较快,经常应用于亲缘关系较近的进化分析,可解决传统分类技术所不能解决的问题[12]。
近年对昆虫的密码子偏好性研究主要集中于目级阶元及以上(半翅目、膜翅目、鳞翅目和双翅目等),利用线粒体基因组中的某些基因或联合一些基因进行分析[13-14]。张玉波等[15]对半翅目线粒体的COX1基因进行密码子偏好性聚类分析,表明同一单系群里进化越快的昆虫其COX1基因的密码子偏好性越强。BEHURA等[16]通过比较分析双翅目与膜翅目22种昆虫基因组序列的密码子使用模式,发现双翅目偏好使用G/C结尾,而膜翅目偏好A/T结尾。GUAN等[17]对蜻蜓目线粒体基因组中密码子使用综合分析,发现豆娘的密码子使用偏好比蜻蜓弱,可能是由于蜻蜓比豆娘具有更高效的线粒体基因表达水平。但是针对半翅目益蝽亚科这类天敌昆虫的详细研究还未见报道,仅有MU等[18]对蠋蝽和益蝽进行测序及构建异翅亚目系统发育树与姜培[19]基于线粒体基因组构建异翅亚目系统发育树,但二者对于密码子的偏好性研究只基于同义密码子相对使用度来研究,研究方法相对单一。
本研究通过分析蠋蝽线粒体基因组13个蛋白编码基因,并利用生物信息学方法探讨该物种以及益蝽亚科的密码子偏好的使用模式,旨在为蝽科提供一些基本的生物学信息,增加蠋蝽的一些相关基因信息,探明该物种的进化机制及其进化关系;同时,蠋蝽作为重要的天敌昆虫,对其进行基因水平的研究将为该种的生物防治应用提供一定的理论基础。
1 材料和方法
1.1 序列信息
蠋蝽及其他蝽科物种的登录号如表1所示。

表1 蠋蝽及其他蝽科物种的登录号Tab.1 The accession numbers of Arma custos and other Pentatomidae species
从国家生物技术信息中心(NCBI)公开的GenBank数据库中检索蠋蝽及其他蝽科的线粒体基因组序列(https://www.ncbi.nlm.nih.gov/)(表1),蠋蝽线粒体基因组序列的GenBank登录号为NC_051562,使用Geneious 8.0.4[20]软件提取其线粒体基因组的13条蛋白编码序列。
1.2 试验方法
1.2.2 密码子偏好性分析 在软件MEGA 7.0 中计算益蝽亚科8个物种(蠋蝽、蓝蝽、峰疣蝽、黑益蝽、益蝽、黑曙厉蝽、叉角曙厉蝽以及奥喙蝽)的13个蛋白编码基因的同义密码子相对使用度(Relative synonymous codon usage,RSCU),然后利用软件Heml 1.0.3.7[22]根据RSCU值绘制热图。
1.2.3 蛋白质性质分析 蛋白质的亲水性、芳香性以及蛋白质长度等[23]被认为是影响线粒体基因组蛋白编码基因密码子偏好性的因素。因此,利用软件Codon W1.4.2[24]来计算出蠋蝽线粒体基因组蛋白编码基因的蛋白质性质,并通过SPSS 26.0[25]软件对ENC值和蛋白质性质之间的相关性分析。
1.2.4 有效密码子数(Effective number of codons,ENC或Nc) 使用软件Codon W1.4.2计算益蝽亚科8个物种的13个蛋白编码基因的ENC值,其值的范围在20~61,用以反映其蛋白编码基因中的密码子使用情况。一般来说,当ENC值为35时,被认为是判断密码子偏好性强弱的界限;当ENC<35时,表示该基因具有较强的偏好性;而当ENC>35时,表示该基因具有较弱的偏好性[26]。ENC-plot 绘图分析是以GC3含量为横坐标、有效密码子数(ENC)为纵坐标构建一个散点图,并在图中添加一条ENC期望曲线(除突变压力的影响外,不考虑其他因素)。
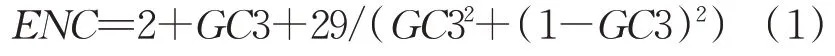
其中,位于曲线上方的点表示密码子偏好是由突变引起的,而位于曲线下方的点表示其受到选择或其他因素的影响[26]。
1.2.5 奇偶图分析 奇偶图是以A3/(A3+T3)为纵坐标、G3/(G3+C3)为横坐标构建的一个散点图。该图的中心点(A=T、G=C)表示其密码子偏好性完全受到突变的影响,数据点根据中心点来分析该物种的蛋白编码基因的偏性方向和程度。
1.2.6 中性图分析 中性图是以GC3为横坐标、密码子第1、2位GC含量平均值(GC12)为纵坐标来构建。如果GC12和GC3间显著相关,则表明突变压力主导密码子偏好性;若GC12和GC3间相关性不显著,则表明自然选择在密码子偏好性形成中影响较大。
1.2.7 系统发育分析 选取45个物种的线粒体基因组蛋白编码基因序列构建系统发育树,其中选择角盾蝽和紫丽盾蝽作为外类群,用PartitionFinder v.1.1.1[27]进行模型分区后,用软件MrBayes v.3.2.5[28]在具有4个独立马尔可夫链(MCMC)的GTR+G+I替代模型下运行10 000 000代,并且当平均标准偏差值低于0.01时停止,前25%的树作为老化被丢弃,其余的树用于构建50%多数共识树。
2 结果与分析
2.1 核苷酸组成分析
蠋蝽线粒体基因组蛋白编码基因结构如表2所示。
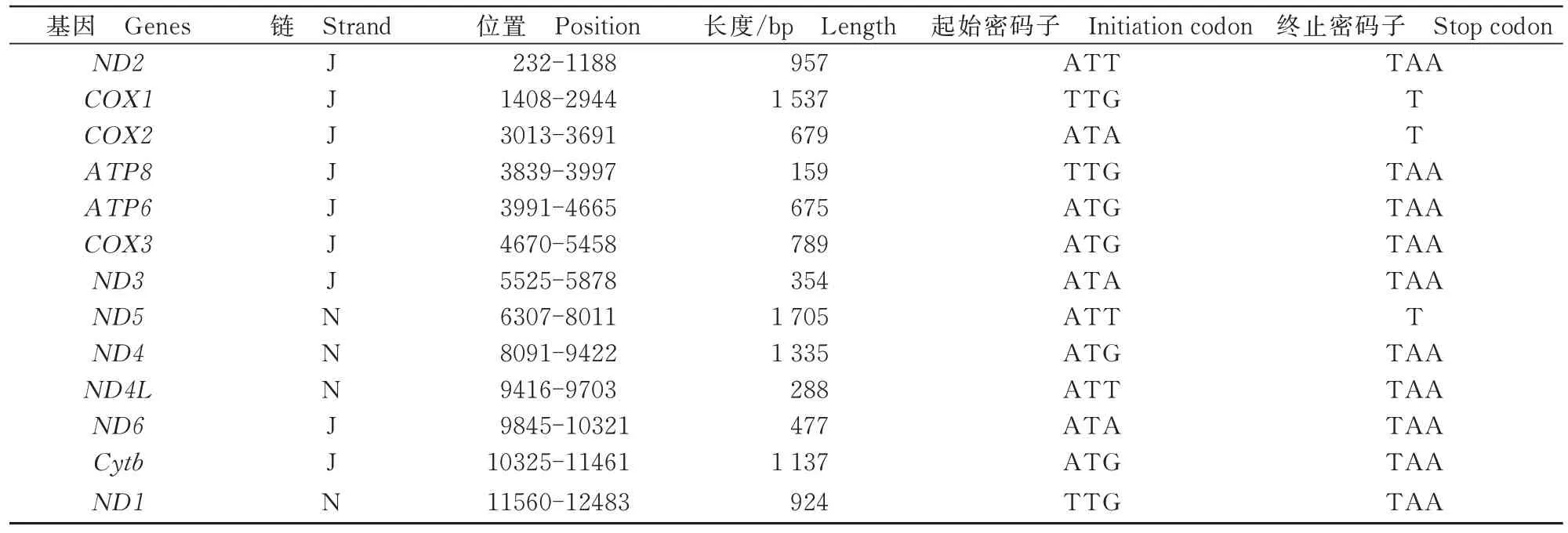
表2 蠋蝽线粒体基因组蛋白编码基因结构Tab.2 Protein coding gene structure in mitochondrial genome of Arma custos
由表2可知,蠋蝽线粒体基因组的13个蛋白编码 基 因 是 以 ATN(ND2、COX2、ATP6、COX3、ND3、ND5、ND4、ND4L、ND6和Cytb)和 TTG(COX1、ATP8和ND1)作为其起始密码子,该物种大多数的蛋白编码基因是以TAA作为其终止密码子的,只有COX1、COX2和ND5基因是以T结尾的。
1.培养良好的用餐行为。一日三餐要有规律,不规律的饮食会影响胃部的消化,导致胃黏膜损伤形成胃炎,出现胃痛,也会导致机体能量不足,出现营养不良、低血糖、低血压等现象。而且“一日之计在于晨”,早餐在一天中尤为重要,人体经过一夜的睡眠,体内的营养已消耗殆尽,血糖浓度处于偏低状态,不吃或少吃早餐,不能及时充分补充血糖浓度,上午就会出现头昏心慌、四肢无力、精神不振等症状,甚至出现低血糖休克,影响正常工作与学习。同时,晚餐也不宜太晚,且不宜过分油腻。
在不受到自然选择的情况下,突变会影响到其核苷酸组成,由于密码子的前2位基本保持不变,所以这种影响会反映到密码子的第3位上。蠋蝽的线粒体总基因组为15 629 bp,核苷酸组成分析结果显示,A(42.1%)是含量最高的碱基,其次分别是T(33.7%)、C(13.7%)和G(10.5%)。核苷酸使用频率的趋势是A>T>C>G(图1)。通过计算蠋蝽的线粒体基因组总碱基含量及第3位密码子的碱基含量,得到总GC含量为24.2%,与其他益蝽亚科(蓝蝽22.9%,峰疣蝽23.0%,奥喙蝽24.9%,黑益蝽28.3%,益蝽24.69%,黑曙厉蝽24.58%,叉角曙厉蝽24.47%)相差不大。同样,第3密码子位置的碱基含量表现出同样的趋势,即A3(41.1%)最高,其次依次是T3(34.4%)、C3(13.8%)和G3(10.8%),而这些组成变化也都表明了密码子的使用偏好。
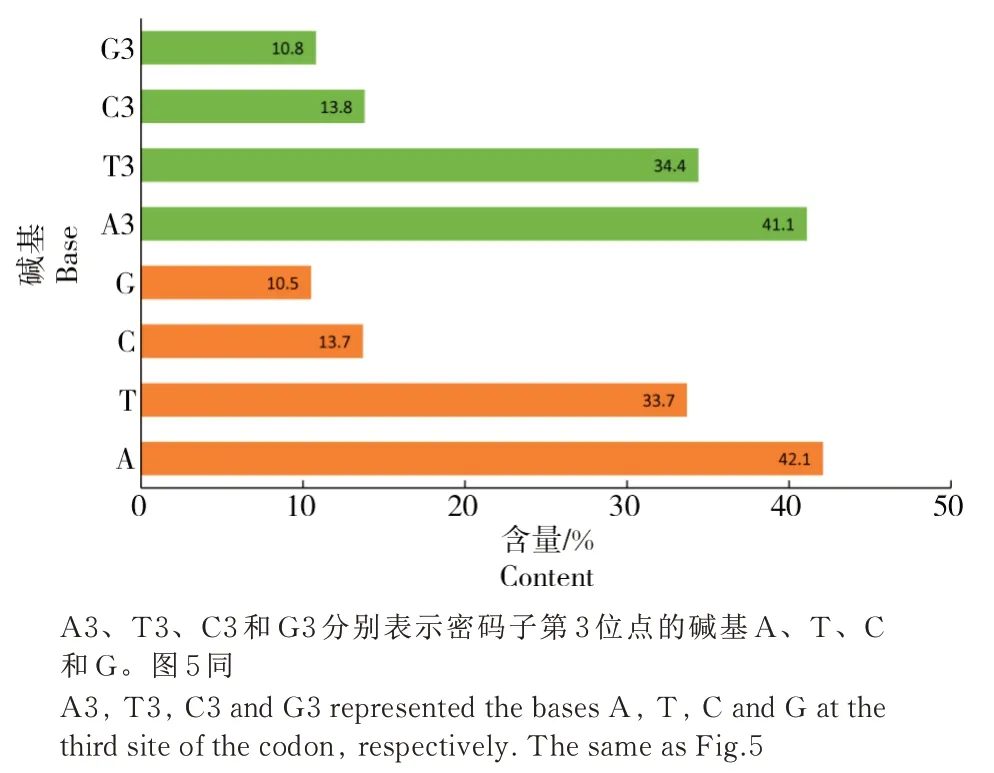
图1 蠋蝽线粒体基因组核苷酸组成及其第3密码子核苷酸组成Fig.1 Nucleotide composition of the mitochondrial genome of Arma custos and the third codon nucleotide composition
2.2 编码基因的RSCU效应
在益蝽亚科蛋白编码基因的62个同义密码子中(去除2个终止密码子),不同颜色显示出在不同蛋白编码基因之间的显著差异,表明在不同物种之间不同的蛋白编码基因间密码子的使用模式是不同的(图2)。
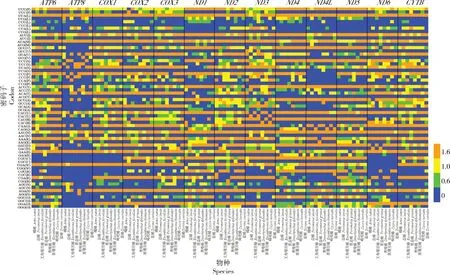
图2 益蝽亚科线粒体基因组13个蛋白编码基因的62个密码子RSCU值热图Fig.2 Heat map of the RSCU values of 62 codons of 13 protein coding genes in the mitochondrial genome of Asopinae
在益蝽亚科8个物种的13个蛋白编码基因中,RSCU≥1的密码子有TTA、ATT和ATA,这些密码子表现了较高的使用频率;RSCU<1的密码子有 CTC、CTG、ATC、ATG、TCG、CCG、ACG 与AGC,表现出了较低的使用频率。这些结果反映了益蝽亚科线粒体蛋白编码基因的密码子使用更偏向于以A/T结尾的密码子。
2.3 蛋白质性质分析
通过对蠋蝽13个蛋白编码基因的蛋白质疏水性和芳香性进行分析,结果如表3所示。
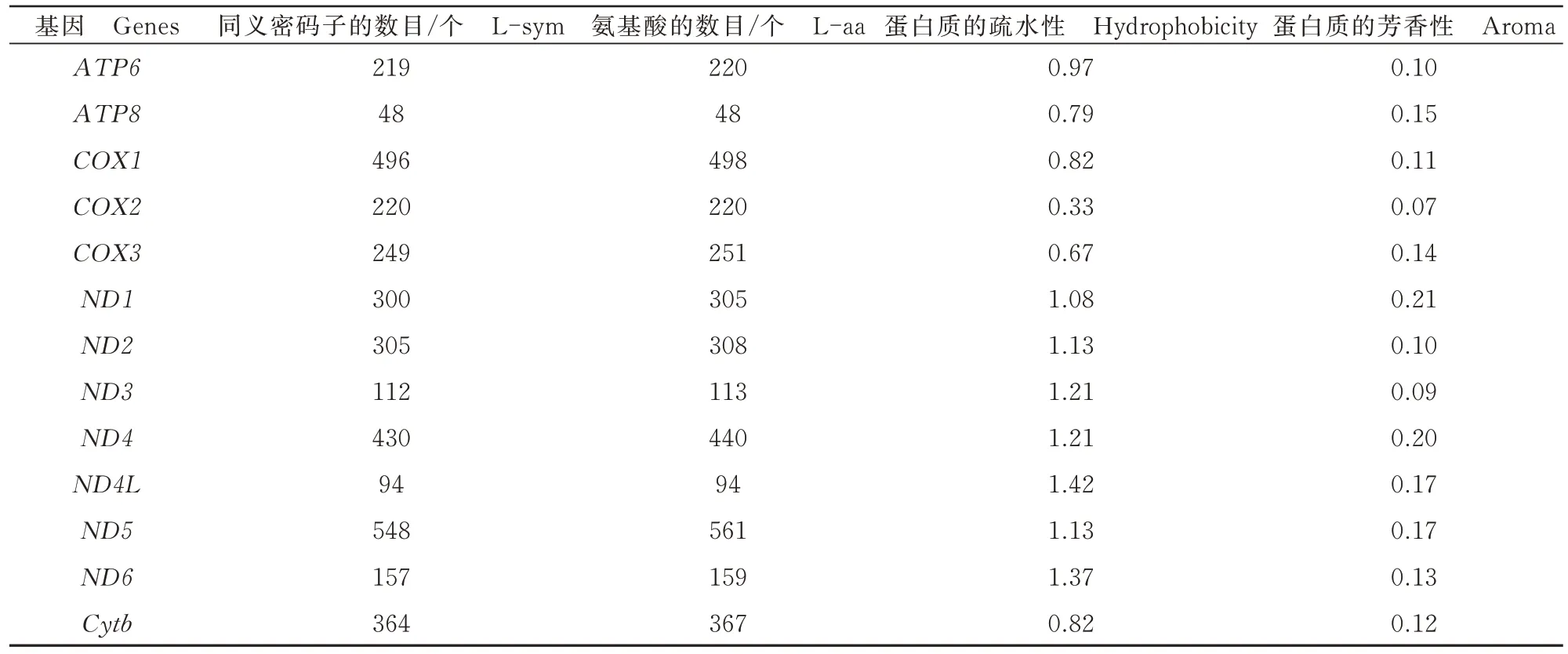
表3 蠋蝽线粒体基因组13个蛋白编码基因的蛋白质性质Tab.3 Protein properties of 13 protein coding genes in mitochondrial genome of Arma custos
从表3可以看出,蠋蝽的ND4L相较于其他的基因,蛋白质的疏水性最高为1.42,则其亲水性最低;而COX2的疏水性最低为0.33,则其亲水性最高。同时,蠋蝽COX2的芳香性相较于其他基因最低。通过分析ENC与Gravy(蛋白质的疏水性)之间的相关性,发现二者之间存在较低的相关性(r=0.421),表明蛋白质的性质会影响蠋蝽线粒体基因组蛋白编码基因密码子偏好性,但是影响比较小。
2.4 ENC绘图分析结果
蠋蝽蛋白编码基因的ENC值的范围在29.51~37.45,其平均值为33.60,表明其具有较强的密码子偏好性;而黑益蝽的蛋白编码基因ENC值的范围在34.43~47.45,其平均值为40.03,普遍高于其他的益蝽亚科,表明其具有较弱的密码子偏好性;以及蓝蝽与叉角曙厉蝽ATP8基因的ENC值为61,表明其无密码子偏好性,这可能与其进化速率快、序列较短有关(图3)。
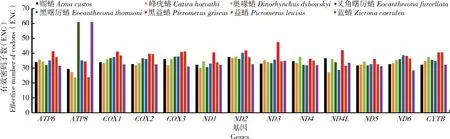
图3 益蝽亚科13个蛋白编码基因的ENC值Fig.3 ENC values of 13 protein-coding genes of Asopinae
根据标准曲线能够确定影响密码子偏好性的主要因素,若点位于标准曲线上方,则表示该点所对应蛋白编码基因的密码子偏好性不受自然选择的影响,结果显示,蠋蝽的ND4L基因、黑益蝽和蓝蝽的ND3基因以及峰疣蝽的ND2、Cytb与ND5基因位于标准曲线上方,其他大多数的蛋白编码基因位于曲线的下方(图4),结果表明,蠋蝽和其他7个物种具有相似的密码子使用方式,其线粒体基因组密码子偏好性主要受自然选择的影响。
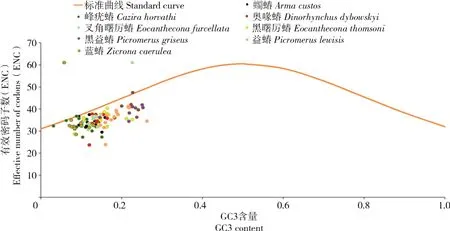
图4 ENC绘图分析Fig.4 Plotting analysis of the ENC
2.5 奇偶图分析
评估A/T和G/C对偏好的方向可以提供更多关于偏离中性突变力量的详细信息,通常使用奇偶图来测量昆虫蛋白编码基因中的这些偏差,结果显示,蠋蝽的8个蛋白编码基因集中于第1象限(A3>T3,C3>G3),3个蛋白编码基因(ND1、ND4和ND5)位于第 4象限(T3>A3,G3>C3),还有 2个基因ATP8和ND4L近似处于同一条直线上(图5-A),并且蠋蝽的线粒体基因组密码子使用模式与其他7种益蝽亚科昆虫的密码子使用模式高度一致(集中位于第1象限与第四象限)(图5-B)。表明益蝽亚科蛋白编码基因密码子的第3位使用具有偏好性,A/T和G/C之间的平衡在益蝽亚科中受到了干扰,说明其密码子的使用模式不仅受到突变的影响,还受到自然选择压力的影响。
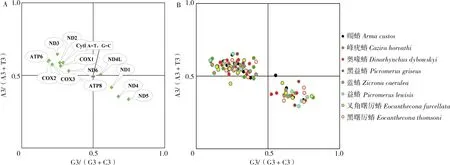
图5 奇偶图绘图分析Fig.5 Plotting analysis of the parity plot
2.6 中性图分析
研究蠋蝽、峰疣蝽、奥喙蝽、黑益蝽、蓝蝽、益蝽、叉角曙厉蝽和黑曙厉蝽线粒体基因组13个蛋白编码基因的GC12与GC3含量,除蓝蝽、叉角曙厉蝽和黑曙厉蝽外,其他物种的GC12与GC3含量均呈正相关,但以上8个物种均呈现出较低的显著性(R2<0.25)(图6)。所有的点呈现不均匀分布,表明自然选择和突变压力同时作用于蛋白编码基因,而密码子偏好的形成主要是受到自然选择作用的影响。
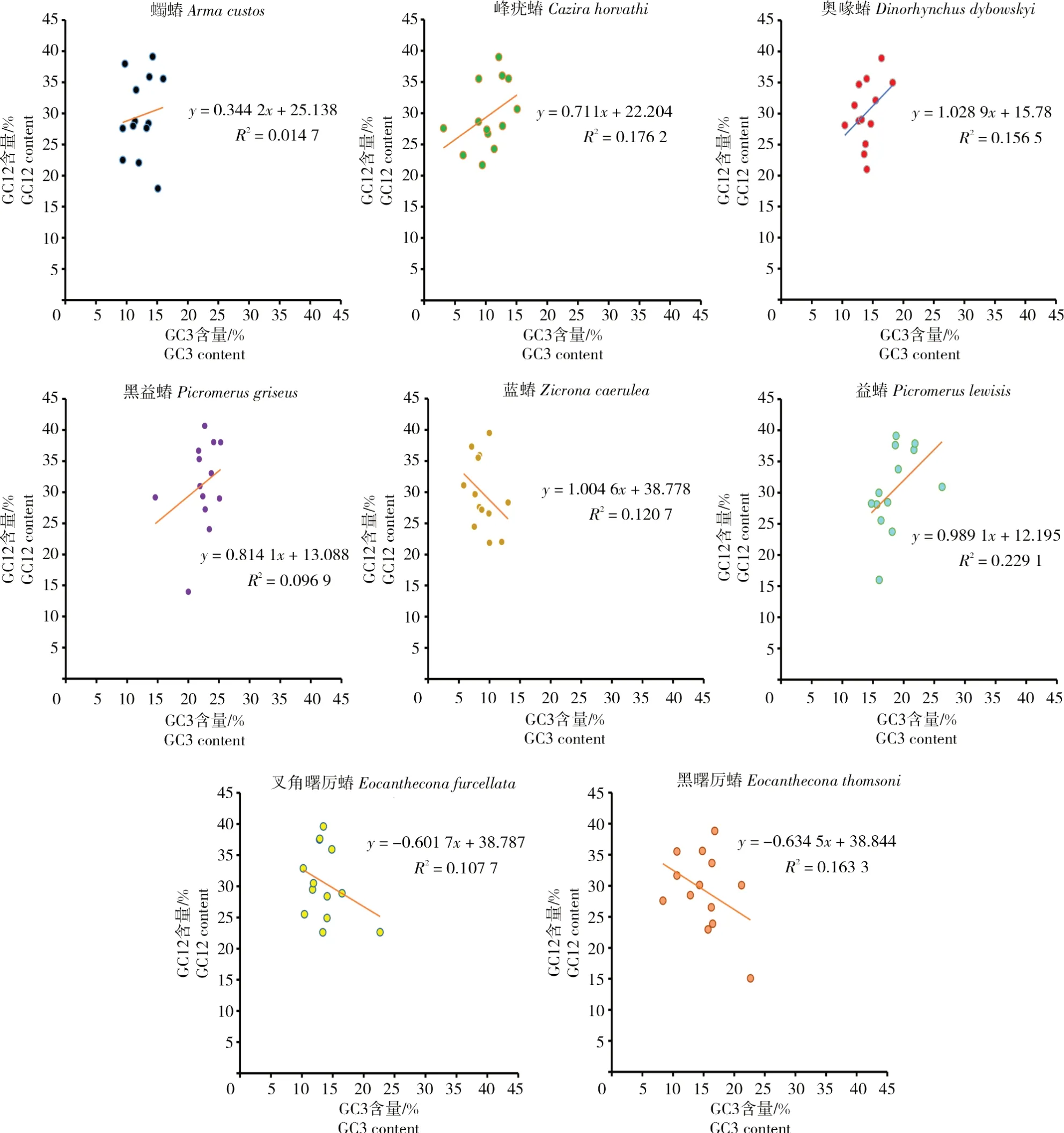
图6 中性图绘图分析Fig.6 Analysis of the neutrality plot
2.7 系统发育分析
比较线粒体基因组序列可以揭示物种之间的系统进化关系,同时阐明昆虫的进化模式。为深入了解蠋蝽在蝽科的进化规律和关系,本研究选取了8种天敌昆虫与蝽亚科35种植食性昆虫作为内群,选取角盾蝽和紫丽盾蝽作为外群,利用线粒体基因组13个蛋白编码基因序列来共同构建系统进化树。基于贝叶斯方法的系统发育树具有较高的自举值,说明其结果较为可靠。结果显示(图7),基于13个蛋白编码基因构建的系统进化树中蝽亚科与益蝽亚科的单系性都能得到很好的聚类。在系统发育树中,蠋蝽与蓝蝽关系较近,二者互为姐妹群。另外,基于蛋白编码基因构建的蝽科系统发育树的拓扑结构和基于形态研究的结果类似,也可证明蛋白编码基因在物种的系统进化关系中具有和形态特征同样的重要性。

图7 蝽科的线粒体基因组进化树分析Fig.7 Phylogenetic tree analysis of mitochondrial genome of Pentatomidae
3 结论与讨论
蛋白编码基因在生物体中具有比较重要的功能,本研究从微观的角度来探究蠋蝽蛋白编码基因的变化,从而为其分子水平的进化提供一定的理论依据。蠋蝽线粒体基因组核苷酸组成呈现出很强的AT偏向性,这与桑丽琴[29]研究的昆虫线粒体基因组和赵婉清等[30]研究的菜蝽结果一致。另一方面,RSCU值显示了益蝽亚科在不同蛋白编码基因之间的显著差异,并表明蛋白编码基因更偏好以A/T结尾的密码子。有效密码子数(ENC)表明,益蝽亚科(除黑益蝽外)具有较强的密码子偏好性。分析密码子偏好性的演化指数(ENC绘图、奇偶图和中性图)得到自然选择是影响密码子偏好性的主要作用。本研究利用线粒体基因组蛋白质编码基因构建蝽科系统发育树的拓扑结构与以往形态学[31]和分子分类[32]的研究结果一致,与蠋蝽亲缘关系较近的是蓝蝽,亲缘关系较远的是黑益蝽。先前的研究表明,密码子的使用偏好性较强可以促进转录和翻译过程的效率[33-35]。蠋蝽比黑益蝽具有较强的密码子偏好性,推测其可能比黑益蝽经历了更多的适应过程和线粒体基因表达的快速变化,从而增加了线粒体代谢过程的能力,有助于它们捕食能力的提高。然而,还有另一种解释(基于不同累积突变量)也是合理的。正如,前面所提到的GUAN等[17]对蜻蜓目线粒体基因组的研究得到,行动力较强的蜻蜓可以迅速去除有害的非同义替换,保留同义替换,定向突变导致某些类型密码子的使用更加频繁。因此,需要对益蝽亚科昆虫的转录和翻译机制进一步研究,这将阐明密码子使用和它们的各种生物学效应之间的关系。
密码子使用偏性是物种在进化过程中适应生长环境而逐渐形成的,通过对密码子的使用情况进行分析有助于研究不同物种的进化和环境适应性。然而,一个基因的密码子使用的偏好性取决于进化压力。构建密码子进化压力在于突变压力和自然选择压力[36]。不同物种或者同一物种的不同基因之间的密码子使用情况可能不同,双翅目偏好使用G/C结尾,而膜翅目偏好A/T结尾。以上结果表明,这些基因组以及基因进化过程中承受的压力不同。本研究首次探讨了蠋蝽及其他益蝽亚科物种的密码子使用模式,并验证了自然选择是影响蠋蝽密码子偏好性的主要作用。同时证明了亲缘关系越近的物种其密码子的使用模式相似度越高,这与张文娟[37]的研究结果一致。然而,由于蛋白编码基因ATP8的进化速率快且序列短的缘故,或许并不适用于密码子偏好性的分析。密码子优化是实现蛋白异源高效表达的重要策略,根据其最优密码子进行密码子优化,可提高蛋白表达水平[38]。但在昆虫线粒体基因组中最适密码子优化尚未见报道,受此限制,密码子偏好模式分析会受到影响,无法探讨其最优密码子。
