北京地区酱香型白酒第六轮次堆积不同位置细菌差异比较研究
2023-02-16张成楠李秀婷王红安孙宝国
陈 曦,张成楠,李秀婷,王红安,王 坤,朱 华,孙宝国
(1.中国商业联合会酿酒微生物与酶分子工程重点实验室,北京 100048;2.北京工商大学食品营养与人类健康高精尖创新中心,北京 100048;3.北京工商大学食品与健康学院,北京 100048;4.北京华都酿酒食品有限责任公司,北京 102212)
酱香型白酒是中国传统的白酒香型之一,广受市场欢迎[1]。酱香型白酒的发酵周期为一年,共有八次的反复发酵,过程非常复杂[2],所产出酱香型白酒共有七个轮次,每轮酒之间都有差异。混合培养发酵、固态发酵、糖化发酵和同时发酵等方法使得酱香型白酒深受地理环境、气候变化的影响[3]。此外,发酵环境也为白酒提供了许多微生物菌种。研究表明,环境(室外地面、室内地面、工具和其他未知环境)对细菌群落的贡献率为62.61 %~90.90 %,对发酵真菌群落的贡献率为20.00 %~38.94 %[4]。发酵温度对微生物群落的生物多样性和微生物代谢也很重要,研究表明,较低的初始温度有利于微生物多样性的维持[5]。
堆积发酵和窖池发酵是每一轮酱香型白酒发酵的最基本步骤[6]。堆积发酵的过程是指将原料蒸熟后摊开冷却,在蒸熟的原料中加入一定比例的大曲,混合后堆积起来,在一定温度下培养4~5 d,在堆子顶部的温度达到45~50 ℃时,结束堆积发酵转而入窖发酵[7]。堆积发酵是典型的多菌种开放的发酵过程,可以充分网罗环境中的微生物[8]。在堆积发酵的中后期,为了维持堆子的稳定性,提升发酵质量,对堆子进行“倒堆”操作,是指在发酵生产中交换原料空间位置的过程,即里外颠倒,使表面层与中心层的酒醅相互交换,之后继续发酵[9]。目前对“倒堆”操作及其对微生物菌群变化的实际影响研究较少。
研究结果显示,在堆积过程中表面层和里芯层细菌的微生物多样性经过“倒堆”后显著下降。从细菌整体的属水平上看,表面层和里芯层共有大量的优势属,没有明显的差异。在“倒堆”过程前后,核心微生物属Lactobacillus、Lentibacillus和Bacillus的相对丰度含量发生了变化,无论是在表面层还是里芯层,Lactobacillus和Lentibacillus均呈负相关关系。目前对北京地区酱香型白酒的“倒堆”操作研究很少,其微生物群落变化的机制尚不明晰。
本研究旨在通过美吉平台提供的数据,研究“倒堆”这一生产操作对堆积过程中微生物细菌菌群结构演替的影响,以便更好地控制酱香型白酒的堆积发酵过程,为今后的实际生产提供理论参考和数据支撑。
1 材料与方法
1.1 材料、试剂及仪器
1.1.1 样品
酒醅样品采自北京华都酿酒食品有限公司。对第六轮次堆积发酵的酒醅进行采样,堆子成型开始记为0 h,堆积发酵总计持续84 h,在60~64 h 对堆积的酒醅进行“倒堆”操作。分别在堆积发酵不同时间点(0 h、24 h、48 h、66 h、72 h、78 h、84 h)的堆子表、里两层进行取样,每层的取样位置固定,基本信息如表1所示,取样后储存于-80 ℃待用。

表1 各取样点的详细编号信息
1.1.2 试剂及耗材
E.Z.N.A ® Soil DNA Kit,美国Omega Bio-Tek 公司;rTaq DNA 聚合酶试剂盒,北京全式金生物技术有限公司;DNA Marker,宝生物工程(大连)有限公司;引物合成,上海生物工程股份有限公司;Gengree染料,上海赛百盛有限公司。
1.1.3 仪器设备
Thermal Cycler 型梯度PCR 仪,美国Bio-rad 公司;Zealway G154DW 型高压蒸汽灭菌锅,致徽(厦门)仪器有限公司;台式高速冷冻离心机,德国Sigma 公司;DYY-8C 型电泳仪,北京六一仪器厂;JS-680C型凝胶成像仪,上海培清科技有限公司。
1.2 实验方法
1.2.1 样品总DNA提取
取酒醅样品10 g于50 mL离心管中,加入30 mL灭菌后的0.1 mol/L 磷酸缓冲盐溶液(PBS)悬浮,加入5~7 颗玻璃珠,充分振荡7 min,400 r/min 离心5 min,吸取上清液。将沉淀用PBS 洗涤,漩涡振荡4 min,400 r/min 离心5 min,收集上清液,重复用PBS 洗涤,直至得到40~50 mL 上清液。将所得液体进行12000 r/min 离心5 min,弃上清液,收集沉淀。总DNA 提取步骤参照E.Z.N.A®Soil DNA Kit操作说明。
1.2.2 PCR扩增
采用16S rRNA通用引物338F(5'-ACTCCTACGGGAGGCAGCAG-3')和806R(5'-GGACTA CHVGGGTWTCTAAT-3')扩增细菌16S rRNA 高变区V3-V4。
PCR 条件为:95 ℃预变性3 min;95 ℃变性30 s;55 ℃退火30 s;72 ℃延伸45 s;30 次循环;72 ℃终延伸10 min,之后降至4 ℃。
PCR 扩增体系包括:5×TransStart Fast Pfubuffer 4 μL,2.5 mmol/L dNTPs 2 μL,正向引物(5 μmol/L)0.8 μL,反向引物(5 μmol/L) 0.8 μL,Trans Start Fast Pfu DNA 聚合酶0.4 μL,模板DNA 10 ng,用ddH2O补至20 μL。
1.2.3 Illumina Miseq 测序
用质量分数为2 %的琼脂糖凝胶回收PCR 产物,利用AxyPrepDNA Gel Extraction Kit 进行纯化,Tris-HCl洗脱,再用质量分数为2%的琼脂糖凝胶进行电泳检测。利用QuantiFluor™-ST 进行检测定量。将纯化后的扩增片段在Illumina MiSeq平台上构建文库,并进行测序,分别对细菌V3-V4 高变区序列进行测序分析。
1.2.4 数据及图片处理
应用Fastp 软件对原始测序序列进行质控,使用Usearch 软件,在97 %的相似性阈值水平下,对基因序列进行OTU 聚类分析。利用RDP classifer进行分类学注释分析。Alpha 多样性指数、优势细菌属的相对丰度均采用单因素方差分析(one way analysis of variance,one way ANOVA)结合最小显著性差异法(Least-Significant Difference,LSD)分析不同堆积发酵时长样品间的差异显著性(P<0.05)。通过unweighted Unifrac 算法计算不同样品间的距离形成Beta 多样性计算矩阵,并利用主坐标分析(Principal Component Analysis,PCA)进行可视化展示。利用Origin 2019b 软件进行图形绘制。
2 结果与分析
2.1 Alpha多样性分析
为探究北京地区第六轮次堆积酒醅细菌群落的动态变化,对42 个酒醅样品中的原核微生物16S rRNA 基因的V3-V4 可变区进行序列分析,获得了3,491,384 个有效基因序列。OTU 聚类分析结果表明,堆积发酵样品的基因序列聚类成1634 个OTU。基于OTU 聚类分析结果,对不同堆积时间的样品进行Alpha 多样性分析,分别计算Chao 指数和Shannon 指数反映酒醅样品中细菌群落的丰富度和多样性[10]。
图1A 和图1B 表明,表面层和里芯层的所有酒醅样品基于Shannon 指数和Chao 指数的稀释曲线均逐渐趋向平缓,表明对酒醅中微生物多样性的分析已覆盖样品中的主要细菌种类[11],样本具有研究价值。Shannon指数反映酒醅样品中细菌群落的多样性,数值越大代表样本中的细菌群落丰度的均匀性越强;Chao 指数反映酒醅样品中细菌群落的丰富度,数值越大代表样本中的物种数量越多。堆积发酵过程中表面层和里芯层细菌的Shannon 指数和Chao 指数在倒堆前呈上升趋势(图1 C、图1D),在堆积刚刚开始时,受车间内各类环境因素例如温度、湿度和工具等影响,原料发酵产生细菌等微生物。“倒堆”前后,表面层与里芯层细菌的Shannon指数显著性下降,说明“倒堆”这个操作扰动了原有的微生物发酵体系,使细菌微生物的α多样性下降;表面层和里芯层的Chao 指数上升说明“倒堆”操作使酒醅充分富集了环境中的微生物,增加了细菌种类。“倒堆”后,表面层和里芯层细菌的微生物菌Shannon 指数下降,Chao 指数先下降再上升。总体上看,表面层和里芯层的Shannon 指数和Chao指数变化趋势一致,与姜明慧等[12]的研究一致。
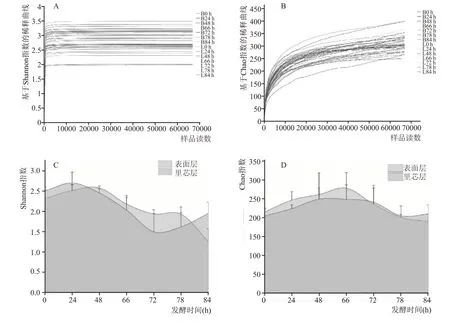
图1 北京地区酱香型白酒第六轮次堆积发酵过程中表面层和里芯层细菌群落的Alpha多样性分析
2.2 群落组成分析
为了探究第六轮次堆积发酵中细菌α多样性变化的原因,本研究进一步分析了第六轮次堆积发酵中表面层和里芯层不同发酵时长的酒醅样品在门水平上的群落结构差异,如图2所示。
第六轮次表面层和里芯层堆积发酵样品中所检出的细菌群落归属于29 个门。将相对丰度大于1%的细菌门定义为优势细菌门。从图2 可以清晰看出,表面层和里芯层的优势细菌门均为Firmicutes、Actinobacteria 和Proteobacteria。“倒堆”前后,表面层、里芯层的Firmicutes 丰度增加,Actinobacteria 丰度减少,Wang 等[13]研究发现习水地区7个发酵轮次中细菌优势门为Firmicutes、Actinobacteria、Proteobacteria、Bacteroidetes 和Fusobacteria,与本研究结果基本一致。戴奕杰等[14]研究了同样来源于贵州省产区的酱香型白酒,发现在第六轮次堆积发酵中优势细菌门包括Firmicutes、Actinobacteria、Proteobacteria 和Bacteroidetes 等,其中Proteobacteria 的相对丰度略高于Firmicutes。
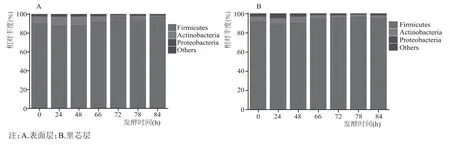
图2 北京地区酱香型白酒第六轮次堆积发酵过程中表面层和里芯层细菌在门水平上的群落组成
在门水平分析基础上,对42 个酒醅样品进行了科水平上的群落组成分析,发现第六轮次表面层和里芯层堆积发酵样品中所检出的细菌群落归属于285 个科。将相对丰度大于1 %的的细菌科定义为优势细菌科。在表面层酒醅中检测出Lactobacillaceae、Bacillaceae、Thermoactinomycetaceae、Staphylococcaceae、Pseudonocardiaceae、Leuconostocaceae、Sporolactobacillaceae、Actinopolysporaceae和Corynebacteriaceae 共9 个优势科。在里芯层酒醅中检测出Lactobacillaceae、Bacillaceae、Thermoactinomycetaceae、Staphylococcaceae、Pseudonocardiaceae、Sporolactobacillaceae、Leuconostocaceae、Enterobacteriaceae、Planococcaceae、Actinopolysporaceae 和Moraxellaceae 共11 个优势科(图3)。已有研究发现Actinopolysporaceae、Bacillaceae、Lactobacillaceae、Leuconostocaceae、Sporolactobacillaceae、Staphylococcaceae、Thermoactinomycetaceae、Moraxellaceae 和Pseudonocardiaceae 是酱香型大曲细菌的主体科组成[15]。
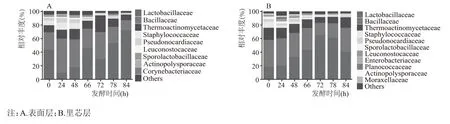
图3 北京地区酱香型白酒第六轮次堆积发酵过程中表面层和里芯层细菌在科水平上的群落组成
由图5 可知,表面层7 个取样时间点的酒醅样品中总共有124 个属;里芯层7 个取样点的酒醅样品中总共有116 个属。研究发现表面层经过“倒堆”后独有的菌属增加,里芯层则相反。将北京地区第六轮次表面层和里芯层堆积发酵样品中所检出的细菌群落归属于706 个属。将相对丰度大于1%的的细菌属定义为优势细菌属。由图4A 和图4B 可知,表面层细菌样品中的优势属共15 个:Lactobacillus、Lentibacillus、Kroppenstedtia、Oceanobacillus、Bacillus、Staphylococcus、Pediococcus、Saccharopolyspora、Thermoactinomyces、Weissella、Sporolactobacillaceae、Bacillaceae、norank _f__Actinopolysporaceae、Pseudogracilibacillus和Corynebacterium_1。里芯层细菌样品中的优势属为Lactobacillus、Lentibacillus、Kroppenstedtia、Oceanobacillus、Bacillus、Staphylococcus、Thermoactinomyces、Pedio-coccus、Saccharopolyspora、Sporolactobacillaceae、Weissella、Bacillaceae、Actinopolysporaceae、Kurthia、Acinetobacter和Enterobacter。与其他地区的优势细菌属类似[16]。
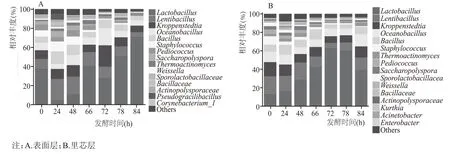
图4 北京地区酱香型白酒第六轮次堆积发酵过程中表面层和里芯层细菌在属水平上的群落组成
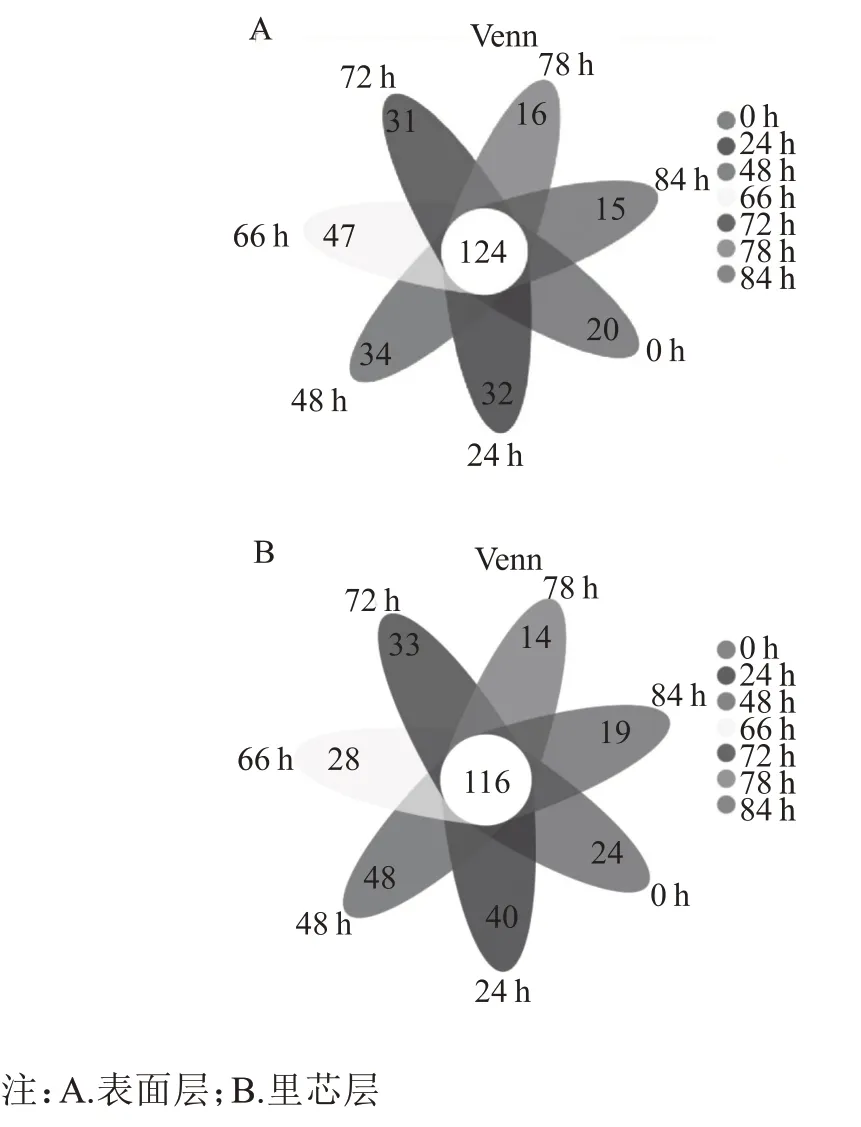
图5 北京地区酱香型白酒第六轮次堆积发酵过程Venn图
“倒堆”前,表面层Lactobacillus相对丰度含量下降,里芯层Lactobacillus相对丰度含量上升,可能是因为Lactobacillus为厌氧型细菌,表面层与外界环境接触距离近,氧含量高,里芯层与外界环境距离远,氧含量低。表面层的Lentibacillus和Bacillus丰度含量上升,里芯层的Lentibacillus和Bacillus丰度含量下降,有研究表明,Lentibacillus物种属于Bacillaceae,在白酒生产中的作用可能与Bacillus属相似[17]。“倒堆”前后,表面层和里芯层的Lactobacillus丰度含量上升,Lentibacillus丰度含量下降,Zhang 等[18]的研究发现,环境温度和堆积所产生的生物热导致堆积酒醅中温度的差异,可以影响Lactobacillus的生长代谢,进而影响整体的细菌群落结构及演替规律[19]。“倒堆”后,表面层的Lactobacillus相对丰度含量显著上升并达到70.8%(B84 h),里芯层的Lactobacillus相对丰度含量呈下降趋势。表面层的Bacillus丰度含量下降,在里芯层的丰度含量则上升。有研究报道芽孢杆菌属Bacillus属于嗜热菌属,不仅产生可以促进酒醅液化的α-淀粉酶,还产生中性或酸性蛋白酶,这些生物酶催化淀粉和蛋白质降解为葡萄糖、氨基酸和高级脂肪酸[20]。
2.3 相关性网络分析
为进一步探究北京地区酱香型白酒第六轮次堆积发酵过程中表面层与里芯层细菌群落结构的动态变化规律,聚焦堆积发酵过程中相对丰度较高的细菌属,分别计算两两细菌属间的Spearman 相关系数,绘制相关性网络图,见图6。由图6A 和图6B 中可知,第六轮次堆积发酵表面层的相关性网络图包含39 个细菌属间的正相关关系,10 个负相关关系;里芯层的相关性网络图包含44 个细菌属间的正相关关系,13个负相关关系。相关性分析表明大多数细菌属之间呈正相关调节机制。
相关性网络图中的高相关性关键节点是指与分类水平前20 名的物种与其他微生物均有相关性的微生物,又称关键性位点(hubs)[21],其数量和种类可直观的反映微环境中微生物群落结构的稳定性[21]。在图6A 中发现,表面层微生物中共有15 个hubs,图6B 中发现,里芯层微生物中共有18 个hubs。在图4A 和图4B 中相对丰度含量较高的Lactobacillus和Lentibacillus呈负相关,无论是表面层还是里芯层的Lactobacillus均与多个细菌菌属呈负相关关系,成为核心负相关性hubs 之一,与其他9 个细菌菌属存在负相关关系,表明在堆积发酵中Lactobacillus对部分细菌菌属有拮抗作用,Li 等[22]研究发现Lactobacillus的相对丰度在发酵谷物中也占据绝对优势。在堆积发酵过程中,Lactobacillus可以产生乳酸,乳酸是乳酸乙酯的前体,可以增强中国白酒的醇厚感[23];Lactobacillus还可以产生大量乙酸,乙酸的堆积可以显著降低酒醅的pH 值,从而抑制不耐酸微生物的生长[24]。从图6A 中同样可以看出Bacillus与多数细菌菌属呈正相关,成为主要的正相关hubs 之一。芽孢杆菌是一种重要的微生物,在白酒发酵过程中产生芳香物质[25]。地衣芽孢杆菌可以产生四甲基吡嗪、苯乙醇和高级脂肪酸酯,它们给白酒带来独特的风味和额外的健康益 处[26]。Saccharopolyspora和norank_f__Actinopolysporaceae也为核心正相关hubs,分别与10 个和8 个细菌菌属间存在正相关关系。从图6B 中可以得知,里芯层的Lentibacillus与10 个细菌菌属呈正相关,属于核心正相关hubs 之一,这可能与其含量在“倒堆”前较高有关(图4B)。“倒堆”影响了微生物的种类与含量,进而影响了表面层与里芯层微生物间的相互作用关系,最终影响了微生物发酵过程中的生物学调控机制,维持了整个堆积体系的自然稳态发展和动态平衡[27]。
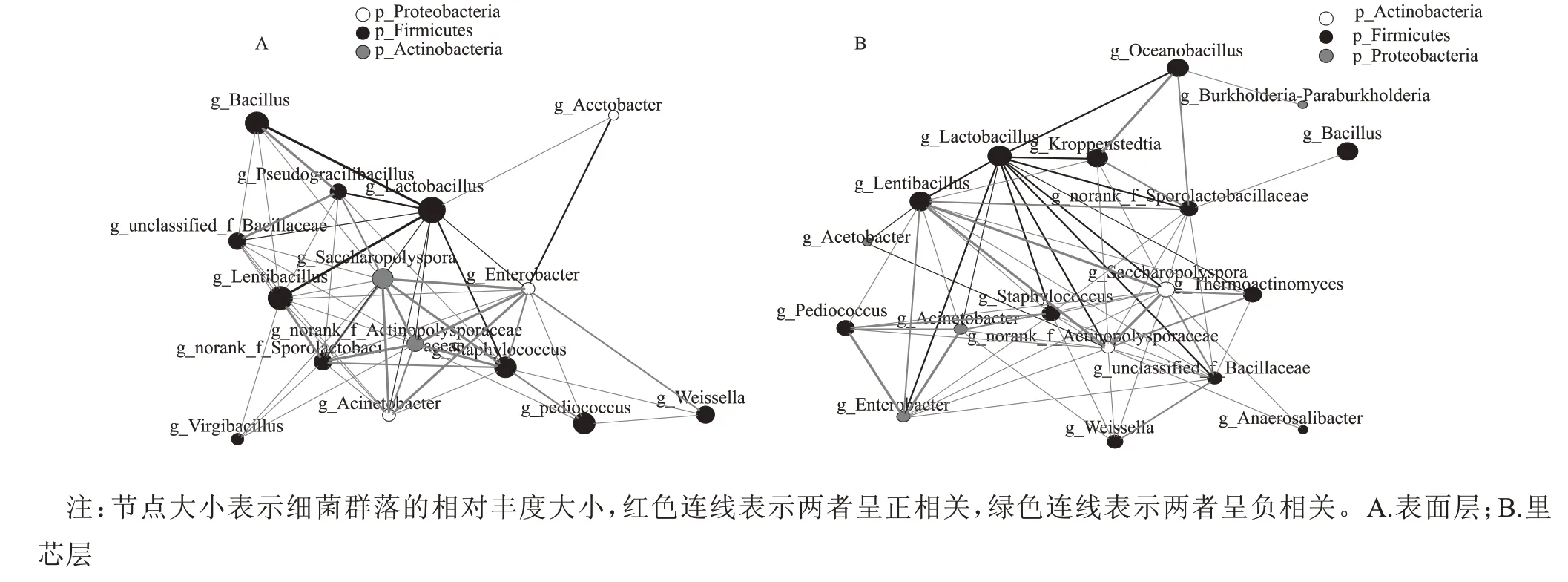
图6 北京地区酱香型白酒第六轮次堆积发酵过程中细菌群落相关性网络图
2.4 Beta多样性分析
从图7 中可以清晰看出,“倒堆”操作的发生扰乱了整个发酵体系,影响了不同堆积时长样本的物种组成。相较于表面层(图7 A),里芯层(图7 B)的细菌菌属群落在“倒堆”操作后变化更为明显。随着堆积发酵时长的延长,堆积发酵堆子的不同空间位置的理化指标(如酸度含量、还原糖含量、酒精度、含氧量与温度)均出现差异[28],“倒堆”操作可以更好的调控这些差异,促进优势菌群的生长[29]。
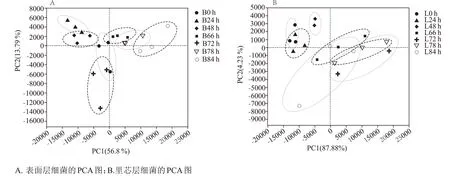
图7 北京地区酱香型白酒第六轮次堆积发酵过程中表面层和里芯层细菌群落的Beta多样性分析
3 结果与讨论
本研究利用高通量测序方法分析比较了北京地区第六轮次酱香型白酒堆积发酵中表面层和里芯层的细菌结构及动态变化。
在细菌α多样性分析上,发现无论是表面层还是里芯层的Shannon 指数在“倒堆”前后均明显下降。“倒堆”操作使表面层好氧微生物埋到里芯层隔绝空气,里芯层厌氧微生物与大量的空气接触,达到控制菌群种类与数量的目的,延长堆积发酵时间使原料各个位置组分发酵更为完全,为入窖发酵做好准备,同时提高最后轮次产酒的质量与风味。
在细菌群落组成分析上,表面层和里芯层的优势属共有12 个,为Lactobacillus、Lentibacillus、Kroppenstedtia、Oceanobacillus、Bacillus、Staphylococcus、Pediococcus、Thermoactinomyces、Saccharopolyspora、Sporolactobacillaceae、Weissella和Bacillaceae,总和占总菌属相对丰度95 %以上。说明“倒堆”操作并没有改变表面层和里芯层的菌属种类,只调控了微生物堆积发酵的过程,维持了整个堆积体系的自然稳态发展和动态平衡。
在细菌Beta多样性分析上,里芯层在“倒堆”操作后,细菌群落组成变化差异比表面层更为明显,这可能是因为除了氧气含量,“倒堆”前堆积发酵中产生的乙酸、乳酸与生物热等随着空间位置的原因积攒在里芯层,导致里芯层细菌受外界环境因素的影响程度强于表面层,群落变化差异明显。
本研究可为了解北京地区酱香型白酒酿造过程中细菌群落演替提供数据支撑,为探究不同产区白酒酿造的特征差异提供科学基础,解释“倒堆”操作对堆积发酵的影响,为生产实际提供基础参数,在未来可以加强对“倒堆”工艺的管理与控制。
