治沙措施通过植物群落的质量效应驱动土壤因子变化
——以玛曲高寒草甸沙化区为例
2023-02-03王新源兰芳芳马仲武王旭洋
王新源,兰芳芳,马仲武,陈 萍,唐 霞,连 杰,王旭洋,曲 浩,*
1 甘肃省生态资源监测中心,兰州 730020 2 甘肃省林业调查规划院,兰州 730020 3 中国科学院西北生态环境资源研究院,兰州 730000 4 中国科学院西北生态环境资源研究院奈曼沙漠化研究站,兰州 730000
青藏高原是我国“两屏三带”生态安全战略格局的重要组成部分,也是我国天然草地分布最为广泛的地理单元之一。其中高寒草甸约占青藏高原草地面积的50%,是反映青藏高原生态功能的主要植被类型[1-3]。伴随着气候变化和人类干扰加剧,高寒草甸退化日趋严重,不利于青藏高原生态功能的维持与稳定[4-5]。生态系统退化主要以植物群落和土壤退化为指征,同样,生态系统的改善亦通过植物群落和土壤的恢复来体现[6-7]。因此,土壤理化性质和植物群落特征成为草地退化与恢复研究关注的重要指标内容,也是定量分析生态系统动态变化和趋势预测的核心参数指征[8-10]。
玛曲县地处青藏高原东北缘,高寒草甸是其境内分布广泛的草地植被类型[11]。源于高海拔和大地貌单元塑造的特殊气候条件,玛曲县高寒草甸生态系统脆弱且敏感[12]。主要表现为在人类活动与自然因素影响下,玛曲高寒草甸生态系统出现湿地萎缩、径流减少、沙漠化等生态问题[13-15],其中草甸沙漠化问题最为普遍,也最为突出[11]。基于玛曲重要的生态地位与社会经济方面的现实需求,生态学者针对玛曲县高寒草甸沙漠化的成因[15]、过程和机理[16]展开了诸多研究。在此基础上,通过植物群落重建状况与土壤质量的改善程度来探索与衡量沙障固定[17]、种草植灌[18—19]等沙漠化防治措施的生态效益。
目前,包括玛曲县在内的青藏高原高寒草甸沙漠化严重区,有关治沙措施效果评估的研究多聚焦于两个方面:一是某一确定恢复措施在不同恢复年限对植物群落和土壤的影响[20—21],这类研究相对较多;二是不同治沙措施对给定时段内植被-土壤系统中某些生态因素的影响[19,22],这方面研究亦有所开展。但有关同一实施时段内,相同治沙措施在不同区域流动沙地治理效果的研究还较少。自2016年起,玛曲县在县城南部黄河岸边及河曲马场南部两个分布有斑块状流动沙丘的高寒草甸退化区开展沙障固定+种草植灌+围封相结合的综合性治沙措施。那么,在治理措施一致的情况下,以上两个不同区域的植物群落是否有效恢复?土壤质量是否改善?土壤质量变化的驱动力主要来自植物群落的物种多样性还是生物量?这种定量分析综合性治沙措施实施后,植物群落是如何优化高寒草甸严重沙化区土壤因子的研究在甘南地区尚属首次,将有助于精确评估植物群落驱动土壤质量改善的机制和探索人工积极干预下高寒草甸严重沙漠化区域生态恢复的模式,从而为治沙工程的完善及精准部署提供参考依据。
1 材料与方法
1.1 研究区概况
玛曲县地处甘肃省西南部,隶属于甘南藏族自治州,位于甘肃、四川和青海3省的交汇地带。全县土地面积为 1.019×104km2,海拔3300—4806 m,是典型的青藏高原高寒生态区。黄河由西部的青海省久治县入境,从南、东、北三个方向形成U形环流,造就黄河首曲第一湾。境内全年无夏,冬季漫长,植被生长季短暂;年均气温仅为1.2 ℃,极端最低气温-28 ℃,最热月(7月)均温11.3 ℃;年均降水量 615.5 mm,年均蒸发量为1347.3 mm,气候呈明显的冷湿特征。自然植被类型主要为高寒草甸,交替分布有高寒草原及沙化草甸等[23]。根据第六次全国荒漠化和沙化监测结果,玛曲县高寒草甸沙漠化严重的区域主要集中于黄河西南岸的河曲马场境内,县域中部曼日玛和采日玛乡交界区及欧拉镇黄河两岸亦有分布。沙丘高约5—30 m,顶部平缓,背部陡峭、呈集中连片或孤立分布,其中集中连片的沙丘面积较大,主要出现在平缓的高寒草甸区;孤立分布的沙丘常出现在小型山坡的迎风面,毛穗赖草(Leymuspaboanus)和青藏苔草(Carexmoorcroftii)是沙化区的主要优势植被[15,24]。
1.2 样地设置与数据采集
2016年开春,选取玛曲县城黄河南岸(下称县城南岸,33°56′—33°58′N,102°03′—102°06′E)和河曲马场东南部(下称河曲马场,33°44′—33°47′N,102°10′—102°13′ E)2个高寒草甸沙漠化严重(已退化为流动沙地)的部分区域,同时采取沙障固定+种草植灌+围封相结合的治沙措施进行修复。具体规格为:铺设1.2 m×1.2 m 麦草沙障,每个沙障方格中心扦插20—30 cm健康的2年生高山柳(Salixcupularis)枝条,同时混播垂穗披碱草(Elymusnutans)和燕麦(Avenasativa)草种;草种质量比2∶1,标准为垂穗披碱草18 kg/hm2,燕麦9 kg/hm2,以上措施实施完毕后立即进行围封处理。
2020年7月底至8月初,植物生长茂盛期,将县城南岸与河曲马场作为调查区,每个调查区的流动沙地和治沙措施区内分别设置6个小型样地作为重复,规模为10 m×10 m,2个调查区共计24个。样地布设在流动沙地和治沙措施区的丘间地上,以尽可能确保立地条件的相似性。单个样地内,沿对角线设置3个1 m×1 m的草本样方;其中治沙措施区的样方布置在高山柳冠层的边缘处,共可获得草本样方72个。确认草本样方中所有植株种类,量度物种丰富度、多度、盖度和株高等群落指标。采用收获法将样方内植物的地上部分收集,去除杂质清洗干净后,于65 ℃烘箱内烘干至恒重,通过称量法获得地上生物量。在样方对角线平均选取3个点,采用土钻法获取0—20 cm的表层土壤,混合均匀后在室内自然风干,剔除杂质后过筛与研磨,用于土壤理化性质测定。
1.3 土壤样品测定
土壤有机质采用重铬酸钾-硫酸容量法分析;全碳和全氮采用Costech ECS4010元素分析仪,按照燃烧法进行测定[25];全磷使用酸消解-钼锑抗比色法测定;速效磷采用碳酸氢钠浸提-钼锑抗比色法测定;速效钾采用乙酸铵浸提-火焰光度计法测定;土壤pH值利用电极法测定[26]。
1.4 数据处理
1.4.1重要值和多样性指数
重要值(IV)是计算多样性指数的基础指标。重要值的公式为:
IV= (RC+RA+RH) /3
式中:RC为相对盖度;RA为相对多度;RH为相对高度。
物种丰富度指数(R)由单个样方内草本植物物种数表示,Shannon多样性(H)和Simpson优势度指数(D)公式为[27]:
式中:Pi是物种i的相对重要值。
基于用一个综合性指标衡量群落物种多样性的考虑,应用主成分分析(PCA),将物种丰富度指数、Shannon多样性指数和Simpson优势度指数3个指标进行降维处理;鉴于仅第1主成分的特征值大于1(2.86),且方差贡献率达95.47%,基于Kaiser-Guttman准则,采用第1主成分载荷因子除以对应主成分特征值的平方根作为指标参数[28-29],可获取物种多样性指数(SD),表达式为:
SD= 0.568R+0.595H+0.574D
式中:R为物种丰富度;H为Shannon多样性指数;D为Simpson优势度指数。
理论上,植物群落主要通过“质量效应”和“多样性效应”2种机制来影响土壤养分[30]。“质量效应”是指植物生物量在改变土壤环境方面具有关键作用[31],“多样性效应”则代表物种多样性是驱动土壤因子变化的主导因素[32]。本研究以地上生物量来反映“质量效应”的影响,应用物种多样性指数SD来表征“多样性效应”。
1.4.2双因素方差分析与线性混合效应模型
应用SPSS 20.0进行双因素方差分析,通过研究调查区和治沙措施对植物地上生物量、物种丰富度、Shannon多样性指数、Simpson优势度指数以及各类土壤因子的影响,判断调查区和治沙措施的主效应与交互效应,交互效应显著(P<0.05)时,进行简单效应分析。
为了定量探讨不同调查区治沙措施实施后植物群落对土壤因子的影响,利用R4.1.2 “lme4”函数包构建线性混合效应模型。模型中,各土壤指标为响应变量,植物地上生物量和物种多样性指数为固定效应(预测变量),样地设置为随机效应。之后,应用“MuMIn”包[33]和“glmm.hp”包[34],定量阐述植物地上生物量与物种多样性对土壤因子变异的解释程度。
由于量纲和数据尺度间的差异,对计算所需的变量进行均值为0,标准差为1的标准化处理。应用Origin 2018和R4.1.2 “forestplot”包绘图。
2 结果与分析
2.1 优势物种的变化
不同调查区治沙措施实施前后物种的组成存在差异(表1)。县城南岸流动沙地的优势种为莎草科(Cyperaceae)的青藏苔草和高山嵩草(Kobresiapygmaea),两物种重要值总和达98.27。经过4 年恢复,高山嵩草和青藏苔草的重要值有所下降,依次为31.69和29.29,仍是群落中的主要优势种。禾本科(Gramineae)植物垂穗披碱草定植成功,重要值为16.36,成为群落中的重要物种。河曲马场流动沙地的优势种为青藏苔草及禾本科的毛穗赖草,重要值分别是60.23和32.80;实施治沙措施后,群落中主要优势种按重要值大小排序依次为青藏苔草、垂穗披碱草和毛穗赖草。可以发现,垂穗披碱草是玛曲高寒草甸沙漠化治理的关键物种,在不同区域均有良好的适应性。
2.2 植物群落数量特征与物种多样性的变化
图1反映了治沙措施和调查区差异对植物群落数量特征及物种多样性的影响。双因素方差分析表明,治沙措施和调查区对地上生物量与Simpson 优势度指数不存在交互效应。治沙措施实施后,县城南岸的植物地上生物量由原始流动沙地的1.51 g/m2显著增加到18.04 g/m2(P<0.05),Simpson 优势度指数则由流动沙地的0.123大幅增加到0.526(P<0.05);河曲马场的植物地上生物量与Simpson 优势度指数分别达到24.67 g/m2和0.692,显著高于未治理的流动沙地(P<0.05)。对于不同调查区,河曲马场流动沙地的Simpson 优势度指数显著高于县城南岸流动沙地,实施治沙措施后,河曲马场与县城南岸的Simpson 优势度指数依次为0.692和0.526,两者间差异同样显著(P<0.05);无论是否实施治沙措施,地上生物量在不同调查区间的差异不显著(P>0.05)。另一方面,治沙措施和调查区对植物盖度、物种丰富度、Shannon多样性指数及物种多样性指数存在极显著的交互效应(P<0.01),说明需进行独立水平的简单效应检验。县城南岸和河曲马场治沙措施区

表1 不同调查区植物物种组成Table 1 Plant species composition of investigation area in our study
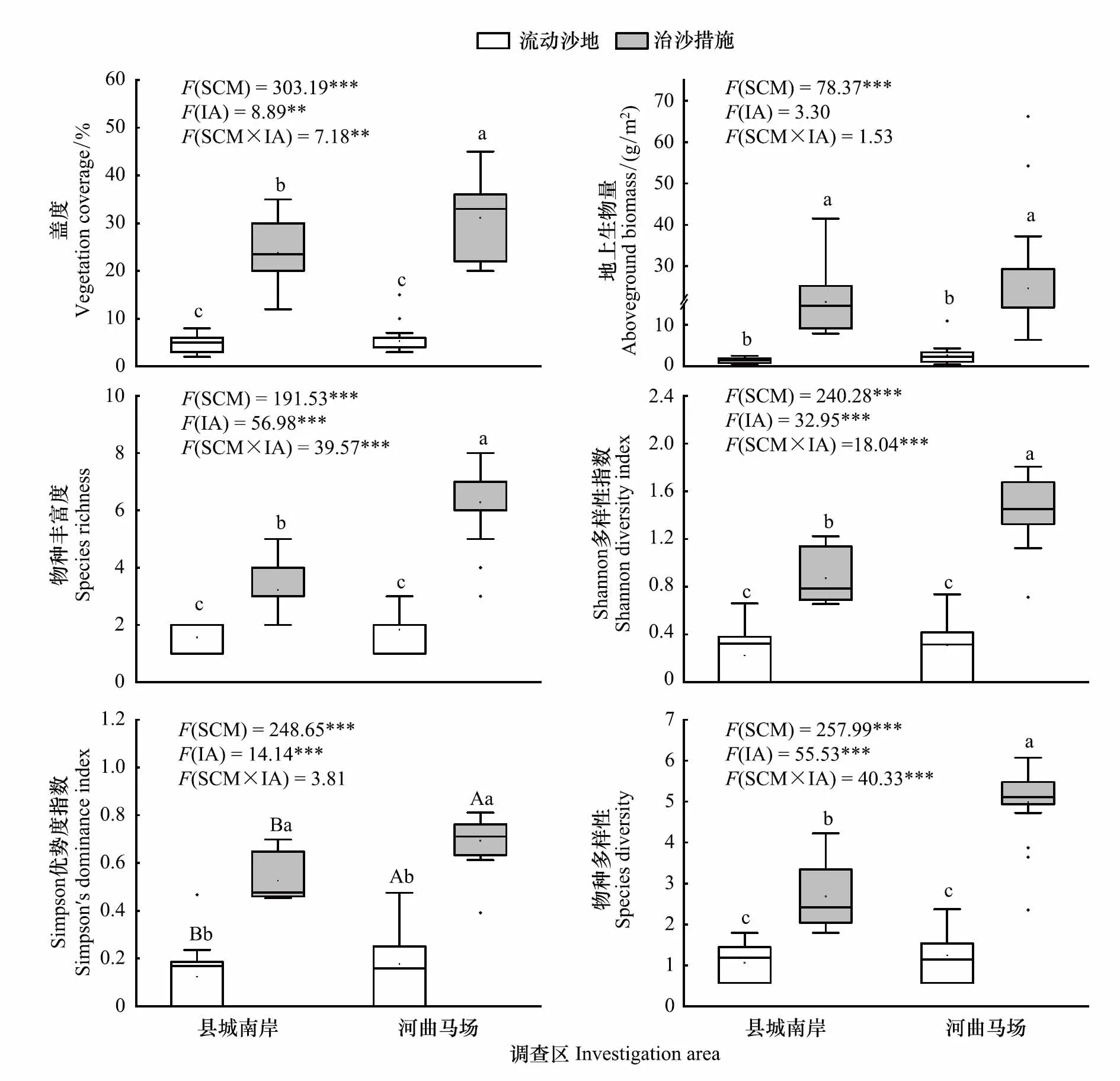
图1 治沙措施对不同调查区植物群落的影响Fig.1 Effects of sand control measures on plant community in different investigation areaSCM:治沙措施,Sand control measures;IA:调查区,Investigation area.*:P<0.05,**:P<0.01,***:P<0.001;不同小写字母代表治沙措施实施前后差异显著P<0.05;不同大写字母代表调查区间差异显著P<0.05
的盖度分别为23.83%和31.11%,较原始流动沙地依次显著提高了487.32%和589.20%(P<0.05)。治沙措施实施前,县城南岸和河曲马场流动沙地间盖度差异不显著(P>0.05);实施后,河曲马场盖度显著高于县城南岸(P<0.05)。进一步探索双因素方差分析结果可知(图1),物种丰富度、Shannon多样性指数与物种多样性指数在统计学上的变化规律与盖度一致。
2.3 土壤因子的变化
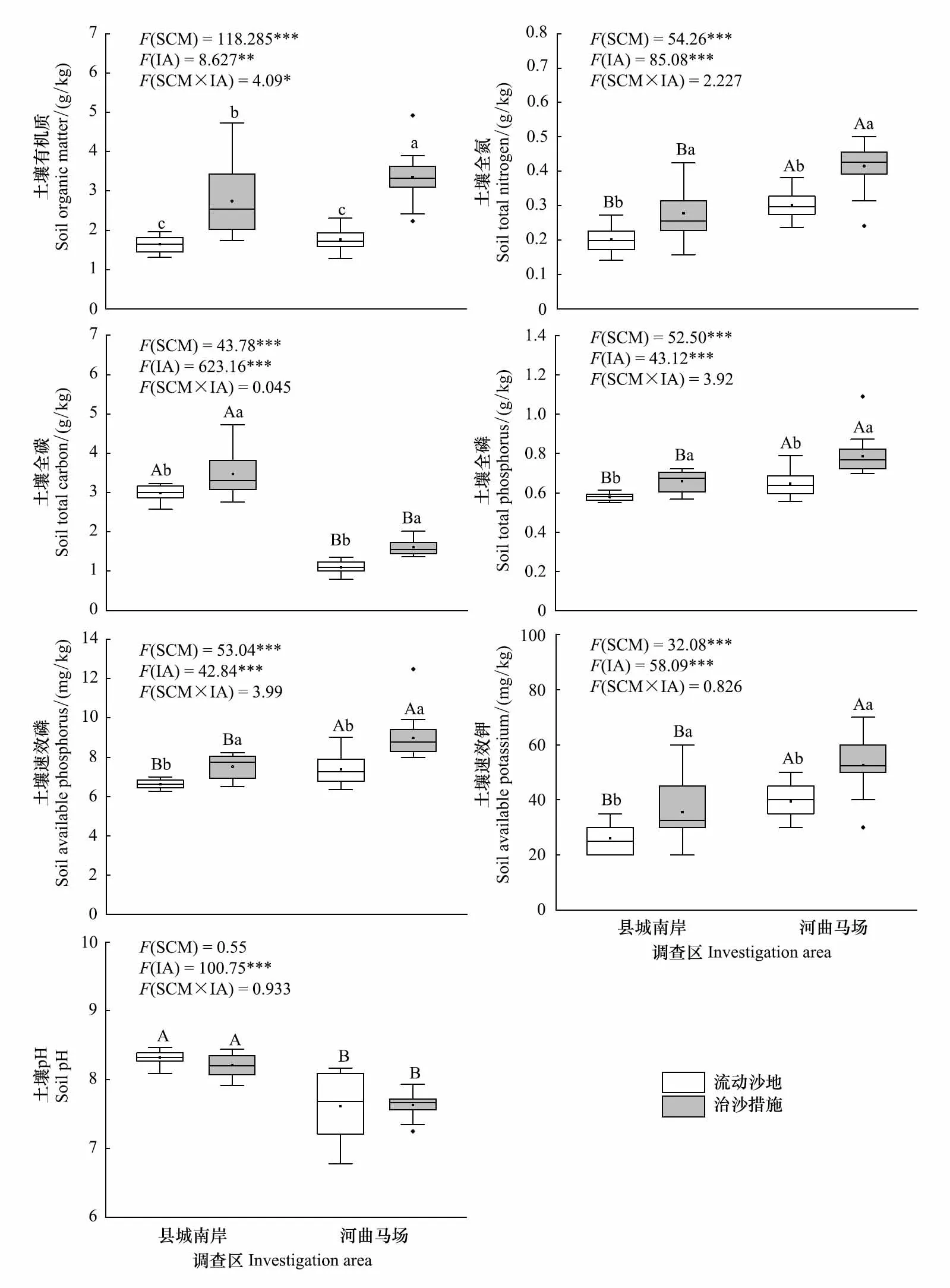
图2 治沙措施对不同调查区土壤因子的影响Fig.2 Effects of sand control measures on soil factors in different investigation area
治沙措施和调查区影响下土壤因子的变化规律如图2所示。土壤全氮、全碳、全磷、速效磷和速效钾受治沙措施和地区差异的影响均达到显著水平(P<0.05),两个因素间不存在交互效应。治沙措施实施后,县城南岸的土壤全氮、全碳、全磷、速效磷和速效钾均大幅提高,与原始流动沙地间的差异达到显著水平(P<0.05);河曲马场以上土壤因子的变化规律及显著性水平(P<0.05)与县城南岸一致。针对不同调查区,土壤全氮、全磷、速效磷和速效钾的差异基本一致,表现为无论是否采取治沙措施,河曲马场的这些土壤因子显著高于县城南岸(P<0.05);土壤全碳则呈相反规律,在县城南岸的含量高于河曲马场(P<0.05)。土壤pH值仅受调查区的影响,对治沙措施的响应不敏感(P>0.05)。另一方面,治沙措施和调查区显著影响土壤有机质含量,且两者间交互效应显著(P<0.05)。简单效应分析表明,县城南岸和河曲马场治沙措施区的土壤有机质分别为2.74 g/kg和3.35 g/kg,显著高于各自区域原始流动沙地的1.64 g/kg和1.76 g/kg (P<0.05)。但就不同调查区而言,县城南岸和河曲马场流动沙地间土壤有机质差异不显著(P>0.05),治沙措施实施后,河曲马场土壤有机质含量显著高于县城南岸(P<0.05)。
2.4 治沙措施实施后植物对土壤的影响
2.4.1治沙措施实施区植物群落对土壤因子的影响
图3反映了采取治沙措施后植物地上生物量与物种多样性对土壤因子的影响。在县城南岸,地上生物量能够显著改善土壤有机质、全氮和全碳(P<0.05),效应量分别为0.72、0.94和0.89。而对于土壤pH值,地上生物量产生显著的负效应(P<0.05),效应量为-0.84。针对其他土壤养分,地上生物量的影响不明显(P>0.05)。不同于地上生物量,物种多样性在县城南岸治沙区域对土壤因子的作用不显著(P>0.05)。在河曲马场,地上生物量对除土壤pH值外的土壤因子均产生正向效应,但仅能显著提高土壤有机质和全碳含量(P<0.05)。物种多样性与土壤全磷及速效磷间呈显著负相关(P<0.05),效应量分别是-0.71和-0.72。对于其他土壤因子,物种多样性的影响不显著(P>0.05)。
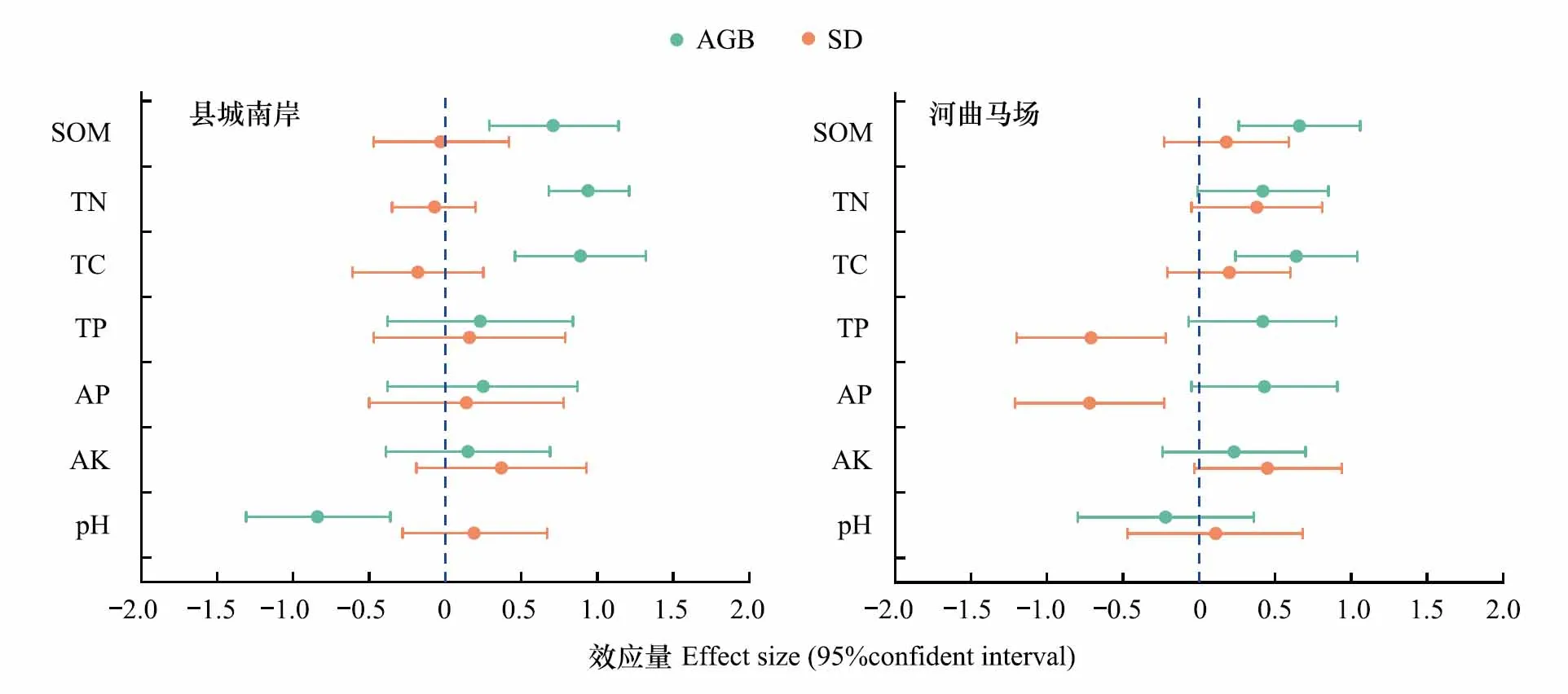
图3 治沙措施实施后植物群落对土壤因子的影响Fig.3 Effect of plant community on soil factors under sand control measures图中效应量是线性混合模型中固定因子的标准化回归系数,蓝色虚线代表斜率为0,水平线段表示效应量的 95%置信区间;当区间范围与0值线不重叠时,效应达到显著水平(P<0.05).AGB:地上生物量,Aboveground biomass;SD:物种多样性,Species diversity;SOM:土壤有机质,Soil organic Matter;TN:全氮,Total nitrogen;TC:全碳,Total carbon;TP:全磷,Total phosphorus;AP:速效磷,Available phosphorus;AK:速效钾,Available potassium;pH:土壤pH,Soil pH
2.4.2地上生物量和物种多样性对土壤因子变异的解释
由表2可得,在县城南岸治沙措施区,地上生物量是调控土壤有机质、全氮、全碳和土壤pH值的主导因素,分别能够解释以上4个土壤因子变异的34.91%、65.68%、46.96%和21.25%。物种多样性的作用相对较低,对所有土壤因子变异的解释率均未超过15.82%。在河曲马场治沙措施区,土壤有机质33.75%、全碳38.44%和土壤pH值14.79%的变异归因于地上生物量的变化,对于上述土壤因子的动态变化,地上生物量的贡献率明显高于物种多样性。不同于其他土壤养分,土壤全氮变异的24.47%来自于地上生物量,20.02%源于物种多样性,2个固定效应的贡献程度接近。另一方面,相比地上生物量,物种多样性对土壤全磷、速效磷和速效钾变异的解释能力更强,依次为27.47%、28.27%和22.52%。需要注意的是,无论在哪个调查区,样地(随机效应)和其他因素对土壤全磷和速效磷变异的解释率均处于较高水平。
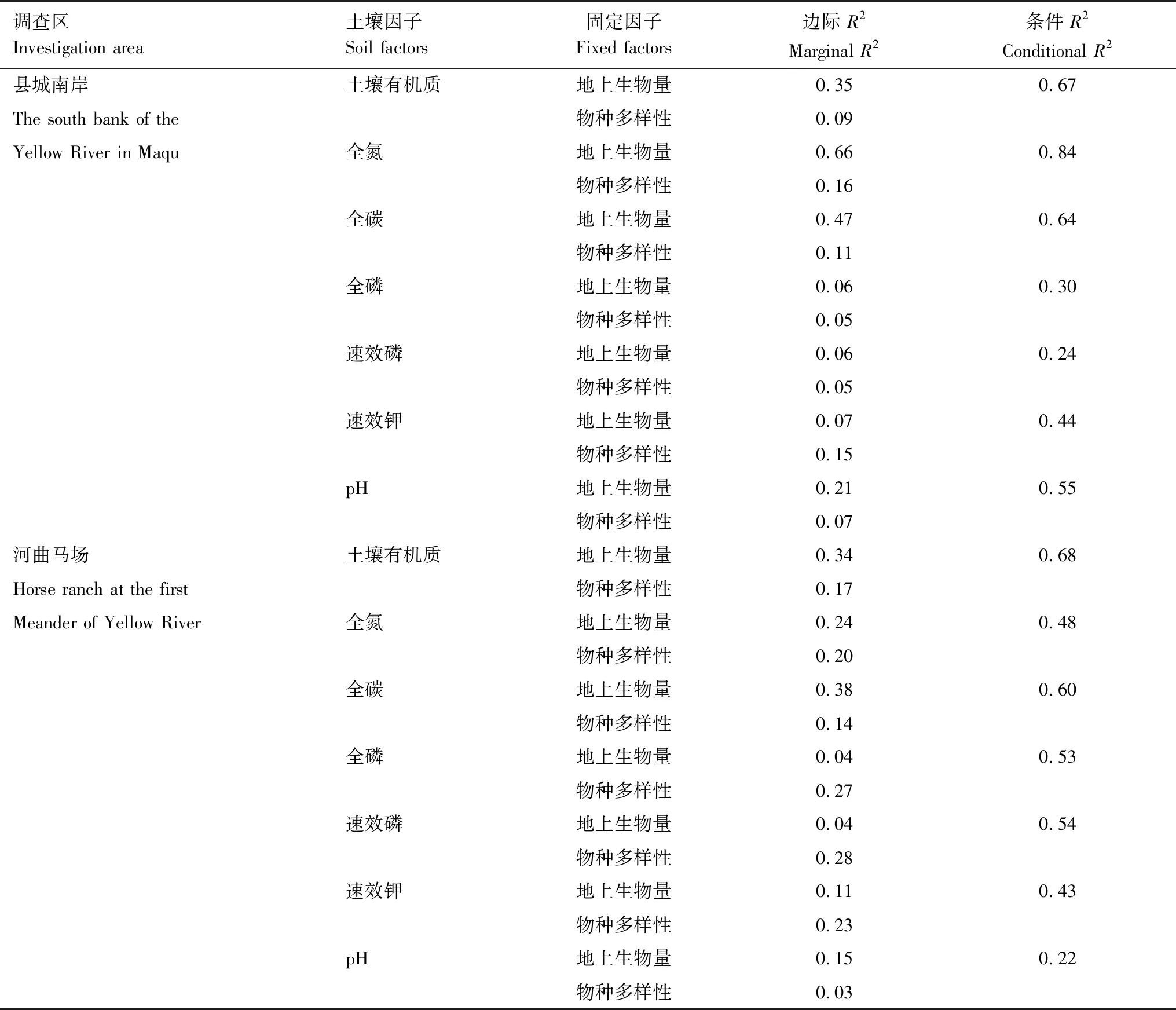
表2 治沙措施实施后植物群落对土壤因子影响的边际R2和条件R2Table 2 Marginal R2 and conditional R2 by the effect of plant community on soil factors under sand control measures
3 讨论
3.1 治沙措施对植物群落和土壤因子的影响
生态恢复的主要目标是重建与改善退化生态系统的服务与功能[35],植物群落和土壤作为生态系统中物质和能量的重要载体与中转站,能够直观反映生态系统的承载力及功能稳定性[36-37],是衡量生态恢复质量的关键指标。本研究中,经过4 年恢复治理,县城南岸与河曲马场高寒草甸严重沙化区(流动沙地)的物种产生了一定程度的替代性变化,由莎草科植物为主转变为莎草科与禾本科植物并重。具体表现为人工播种的垂穗披碱草成为群落中的优势种之一,青藏苔草的优势度下降,但仍是群落中重要值最高的物种;燕麦作为人工混播的另一草种,仅在群落中零星分布。植物群落中物种形成上述分布格局主要归因于优先效应和种间作用的共同影响[38-39]。优先效应体现在青藏苔草作为生境中占据生态位的先锋种,对之后入侵的物种产生抑制作用,而种间作用则更加强调垂穗披碱草良好的生态适应性与竞争力,相较其他草本植物,垂穗披碱草应对干旱和盐分胁迫的能力更强[40]。尽管由于地区因素的作用,2个调查区流动沙地的Simpson 优势度指数间存在差异,但这种差异未影响到治沙措施对该指标的积极效果。本研究发现,实施治沙措施后,植物群落的地上生物量和盖度大幅提高,预示着植被生产力得到有效恢复。另一方面,物种多样性水平亦增加显著,反映出植物群落结构趋于改善,群落弹性加强。植物群落的正向变化,与治沙措施的综合性(沙障固定+种草植灌+围封)密不可分。这其中,麦草沙障通过提高地表粗糙度,有效降低了流水和风力的侵蚀,为植物的萌发与定植提供了条件[41]。围封最大限度排除了牲畜对植物的啃食与踩踏,这有助于植被,尤其是适口性较佳的禾本科植物的生长与恢复[42]。种草植灌措施中,作为玛曲地区的乡土灌木,高山柳可通过防风固沙、优化土壤质地、聚集养分等生态功能,改善草本植物的生存环境[43];但在另一方面,基于高山柳巨大林冠结构与发达根系的影响,冠层下草本植物对光照与养分的利用可能会受到限制[44-45]。然而,相关研究发现,种植高山柳能够大幅提高垂穗披碱草的种子出苗率[40],表明就垂穗披碱草而言,高山柳的作用为协同而非权衡。此外,垂穗披碱草与燕麦的混播,人为形成禾本科植物的入侵效果,整体提高了群落的生产力与多样性。
本研究中,治沙措施能够显著改善除土壤pH值之外的土壤因子,这在很大程度上同样归因于治沙措施的综合性。其中,麦草沙障能够减缓富含土壤养分的细颗粒物质和团聚体的流失[37,41];围封减少了牲畜的践踏、保护了土壤表层结构;种草植灌中,垂穗披碱草属于地上生物量较大,养分含量较高的禾本科植物,可通过根系与凋落物返还营养物质,相较而言,燕麦对土壤有机质、全氮等主要养分的贡献率较低[46],这主要源于两方面原因,一是较低的凋落物输入量与地下生物量[47],二是燕麦属于一年生草本植物,地上植株留存时间较短,对土壤水分的保护能力不及多年生的垂穗披碱草[48],较低的土壤含水率抑制了土壤微生物活性,不利于土壤有机质的矿化和分解;作为高大灌木,高山柳能够形成“沃岛效应”[43,49],有助于土壤养分的存储与富集。不同于其他土壤因子,土壤pH值仅受地区差异影响,可能原因是不同调查区之间土壤母质不同。
综上可知,对于玛曲高寒草甸不同区域的流动沙地(县城南岸与河曲马场),综合性治沙措施(沙障固定+种草植灌+围封)能够显著改善植物群落数量特征、物种多样性水平以及与土壤质量密切相关的各类养分。说明该治沙措施在玛曲地区具有较好的生态效益和普适性。
3.2 治沙措施实施后植物群落驱动土壤因子变化的机制
植被和土壤是密不可分的生态体系。土壤通过提供定植场所、供给水分和养分资源来影响植被的生长和繁育,植被则主要通过根系生长与分泌、生成凋落物等途径来塑造与改善土壤。考虑到土壤相较植物群落具有更高的稳定性,在严重沙化的高寒草甸生态系统,土壤恢复的时间远长于草地植被[3]。因此,对于恢复初期的生态系统,尤其是在以快速重建和改善植物群落为主要治理手段的流动沙地,植物群落在土壤与植被互馈关系中居于主导地位[37]。“质量效应”和“多样性效应”是植物群落影响土壤因子的主要机制[30]。“质量效应”认为包括土壤因子在内的生态系统功能和服务主要决定于植物群落生物量主要贡献者的性状特征[6,31]。而“多样性效应”又称作“单一假说”,是指植物群落中的物种具有唯一性,即每个独立物种都对土壤属性和功能存在影响[50]。
本研究利用线性混合效应模型分析了治沙措施实施后,地上生物量和物种多样性对土壤因子的影响。不受地区因素的干涉,植物地上生物量能够显著或接近显著的提高土壤有机质、全氮和全碳,且可以很大程度上解释以上养分的变化规律。表明由综合性治沙措施塑造的植物群落主要通过“质量效应”来主导土壤有机质和全氮等土壤养分的变化。事实上,“质量效应”驱动土壤因子的核心是与植物群落地上生物量密切相关的凋落物与土壤生物[51]。伴随着群落地上生物量的增加,凋落物产量相应提高,这为土壤动物和微生物的生存与活动提供了物质与能量来源,土壤生物丰度与活性提高反过来加速了凋落物的分解,在这一系列生态过程中,凋落物的产量和分解速率均明显增加,来自植物的大量养分返还到土壤中。优势种中,治沙措施中引入的垂穗披碱草属于禾本科,是典型的高氮型草本植物,产生的凋落物更易被土壤生物分解[52],其地上生物量的增加有助于土壤中氮等养分的富集。地上生物量增加对土壤因子的积极效应还可能来自于其他因素的间接影响[53-54],如植被盖度增加能够降低大风和地表径流的侵蚀作用,利于土壤养分的留存。
不同于地上生物量,物种多样性对土壤有机质、全氮及全碳的影响不显著(图3),且对以上养分变异的解释率均处于较低水平(表2)。究其原因,很大程度上在于治沙措施实施期较短,群落恢复仍处于初级阶段,物种间生态位分化不明显,包括土壤在内的生态系统整体属性和功能主要依靠生物量较高的少数优势种来实现[31]。河曲马场植物多样性与土壤全磷及有效磷间呈现显著负相关,其可能原因在于增加的禾本科植物对于土壤磷的消耗。然而,由物种多样性产生的负面效应并未改变河曲马场治沙区土壤全磷和速效磷显著高于流动沙地的动态变化。鉴于植物物种多样性对于土壤全磷和有效磷变化的解释能力未超过30%(表2),这预示着除了样地引起的随机效应,可能存在其他因素主导着土壤全磷和速效磷的动态变化。

综合可见,通过人类积极干预,玛曲高寒草甸严重沙化区土壤有机质、全氮等关键养分的改善主要决定于植物群落的地上生物量或者是植物群落中贡献主要生物量的少数优势种,而不是物种的多样性水平。鉴于此,建议在实施种草植灌时,应优先考虑植物生物量的增加与稳定,在此基础上,兼顾物种多样性的提高。
4 结论
尽管植物群落和土壤因子的多数指标受调查区变化的影响,但无论在县城南岸还是河曲马场,沙障固定+种草植灌+围封相结合的综合性治沙措施均能显著改善植物群落数量特征、地上生物量与物种多样性。土壤因子中,得益于植物地上生物量的提高,有机质、全氮和全碳等主要养分增加明显。土壤pH值在地区间差异明显,但对治沙措施的响应不敏感。综合来看,综合性治沙措施在种草植灌方面仍存在改进空间,混播草种中燕麦的生态效益不明显,可替换为其他适应性更佳,生产力更高的乡土草种。
