干旱沙区生物土壤结皮对红砂种子定居的影响及作用机制
2023-02-03何芳兰赵赫然王忠文汪媛艳李雪娇金红喜2
何芳兰,赵赫然,王忠文,汪媛艳,李雪娇,金红喜2,,*
1 甘肃河西走廊森林生态系统国家定位研究站, 武威 733000 2 甘肃临泽荒漠生态系统国家定位观测研究站, 临泽 734200 3 甘肃省治沙研究所, 兰州 730070
在干旱沙漠区,沙丘由流动或半固定向固定演替过程中,地表逐渐形成生物土壤结皮,并随时间延长逐渐由物理-藻类阶段逐渐演替到地衣阶段,部分微环境好的区域甚至出现碎片状的苔藓结皮[1—2]。生物土壤结皮有效固定沙面、减少地表风沙活动以及土壤风蚀[3—4];同时不断为土壤提供氮源和能源,促进土壤异养微生物的生长[5—9]。但是,生物土壤结皮形成及发育一方面阻碍降雨及时入渗,导致土壤深层含水量下降[10—15];另一方面减少了沙物质对种子的掩埋,阻碍种子进入深层土壤,使得微区域种子空间分布浅表化[16—18]。因此,沙地生物土壤结皮对部分种子萌发及植株生长产生了一定的限制作用[19—20]。
红砂(Reamuriasongarica)为柽柳科红砂属的一种超旱生小灌木,具有很强的抗旱、耐盐、固沙、集沙和保持水土能力,是保护干旱荒漠化土地的重要生态“屏障”[21—22]。在干旱沙漠区,地表一旦有生物土壤结皮形成,原有固沙灌木不仅单株形态和生理特性表现出明显衰退现象,且林下很难发现实生或根蘖苗[23]。然而,红砂种群不仅可以自然侵入,并能实现种群繁殖与扩散[24]。红砂种子小、轻并附有密集的长种毛,在自然环境中主要通过风媒传播。那么,红砂种子被风媒“送”到固定沙地上后,生物土壤结皮是通过什么生态机制驱使其实现永久定居的?不同发育阶段结皮对红砂种子定居生态机制是否相同?但有关干旱沙漠区地表生物土壤结皮发育对易漂移性植物种子永久定居方面的研究报仍比较少。
为此,本文试图以巴丹吉林沙漠南缘已发育生物土壤结皮的沙包为研究场所,通过对红砂在不同发育阶段结皮上的动态变化、宿存微区特征及宿存种子数量与微区面积间关系等研究,探讨生物土壤结皮发育对易漂移型种子定居影响及作用机制,为干旱沙区植被恢复与重建提供理论基础。
1 研究方法
1.1 研究区自然概况
研究区位于巴丹吉林沙漠南缘 (102°59′05″E,38°34′28″N),该区域属于典型温带大陆性荒漠气候。冬季寒冷,夏季酷热,昼夜温差大,年平均气温7.6 ℃,极端低温-30.8 ℃,极端高温40.0 ℃,无霜期175 d;降雨量小,蒸发量大,气候干燥,年均降雨量113.2 mm,年均蒸发量2604.3 mm,干燥度5.1,最高达18.7,相对湿度47%;光热充足,年均日照时数2799.4 h,≥10 ℃的活动积温3036.4 ℃;冬季盛行西北风,全年风沙日可达83 d,并多集中在2—5月,年均风速2.5 m/s,最大风速为23.0 m/s;地下水位深23 m左右。
1.2 样方布设
在2019年4月上旬,在巴丹吉林沙漠南缘绿洲-荒漠过渡带依据单位面积内流沙以及不同类型生物结皮面积百分比大小,选择结皮前期(NCS)、藻类结皮阶段(ACS)、地衣结皮阶段(LCS)、藓类结皮阶段(MCS)的样地,每个阶段5个重复(详见表1)。分别在2019、2020和2021年4月布设样方,具体如下:
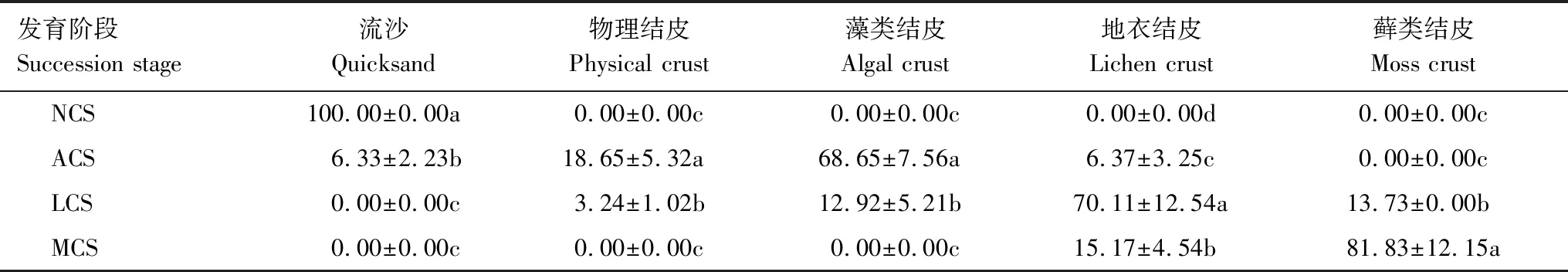
表1 不同发育阶段生物土壤结皮面积占比组成/%Table 1 Proportion compositions of biological soil crusts at different development stage
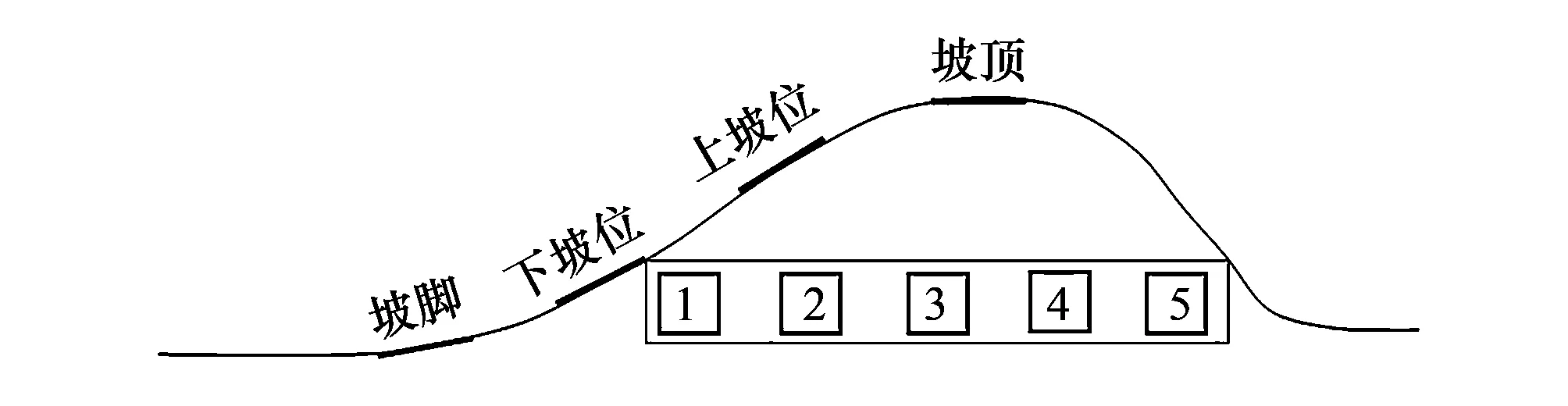
图1 A组样方布设位置示意图Fig.1 Location diagram of group A quadrats 图中1、2、3、4和5为样方编号
A组样方:2019年4月25—26日,在样地区选择互不相邻的大型沙包,并在沙包主风向迎风面坡脚、下坡位、上坡位或坡顶水平方向布设5个50 cm×50 cm样方组(样方编号为1—5,1、3、5为试验样方,2、4为监测样方;样方水平间距大于1 m)(图1),试验样方共300个(坡底、下坡位、上坡位均50个,坡顶100个)。
B组样方:2020年4月8—9日,在ACS、LCS、MCS样地区沙包迎风面的下坡位选择坡度相似的区域,分别布设125、50、50个50 cm×50 cm样方(样方间距大于5 m,避开A组样方),共225个。
C组样方:2021年4月15—16日,分别依据微区内枯落物、微坑(深、口径≥5 mm)、龟裂缝(宽、深≥3 mm)、长藓(≥3 mm)面积变化,在样地区沙包迎风面的下坡位选坡度相似区域,布设20 cm×20 cm(避开A和B组样方)样方(样方间距大于5 m),每种类型40个样方,共160个。
1.3 种子源提供

图2 红砂种子大小及外形Fig.2 Size and shape of R. soongorica seed
2019年4月28—30日,在无风或风力较小的时间段,用尖嘴镊子将饱满种子 “点”于A组1、3、5号样方内(每个样方100粒);2020年4月10—13日,以同样的方法给B组所有样方提供种子(每个样方100粒);2021年4月16—17日,给C组样方提供种子(每个样方40粒),方法同上。试验采用的红砂种子于2018年12月底就地采摘,千粒重约为1.133 g(图2)。
1.4 种子定居动态监测及微生境因子调查
A组样方:监测2、4号样方及其与1、3、5号间隔区域种子动态,并及时清除流入种子;在第1、3、7、15、30、60、90、120天分别统计1、3、5号样方内种子数,同时调查样方内不同微区(如枯落物、微坑、短藓、长藓等)中定居种子数;在样方间隔区域,以30 cm×30 cm标尺框及20 mm针状标尺钎为背景,用佳能EOS 70D单反相机对地表拍照,重复3张。
B组样方:定期清除样方外种子;第120天,调查样方内种子定居数及不同微区上定居种子数;以50 cm×50 cm标尺框为背景,对样方拍照。
C组样方:定期清除样方外种子;第120天,借助20 cm×20 cm标尺框对样方拍照,并分别调查样方内枯落物、微坑、龟裂缝、长藓中的种子数。
以上样方内,种子滞留或定居率=滞留或定居种子数/添加种子数×100%;微区内定居种子密度=定居种子数/微区面积
1.5 数据分析
采用Photoscan、AutoCAD 2019软件对相关照片进行处理,计算出样地区及样方内不同类型结皮、微区的面积。应用Excel和SPSS 22.0软件完成数据处理和统计分析。利用单因素方差分析和邓肯(Duncan)多重比较法对生物结皮发育4个阶段地表微区特征、种子定居能力进行显著性分析 (α=0.05),并对枯草丛、微坑、龟裂缝及长藓面积与其上定居的种子数进行相关性分析(α=0.01)。
2 结果与分析
2.1 生物土壤结皮发育过程中地表微区变化
沙包表面生物土壤结皮由结皮前期演替到藓类结皮过程中,地表出现了一定量的枯落物、微坑微、龟裂缝、藓株,单位面积内其组成及大小均处于变化状态(表2)。由表2可以看出,结皮前期演变成藻类结皮后,枯落物面积增值最大,但其面积百分比仍小于1%;藻类阶段演变到地衣阶段,微坑和龟裂缝增值较为明显,两者面积百分比增值均超过17%;地衣阶段演替到藓类阶段时,苔藓面积急剧增大,特别是长苔藓,面积增值超过50%。同时,随着生物土壤结皮不断演替,地表枯落物、微坑、龟裂缝面积百分比均呈现出先增大后减小的变化趋势,枯落物的峰值出现在藻类-地衣阶段,微坑和龟裂缝峰值均出现在地衣阶段,4个阶段的枯落物、微坑、龟裂缝面积百分比两两之间均存在显著差异性(藻类与地衣阶段的枯落物除外);生物土壤结皮演替到地衣阶段时藓类逐渐出现,并随着演替进行急剧增大,除结皮前期与藻类阶段外,其余各阶段两两之间差异显著。
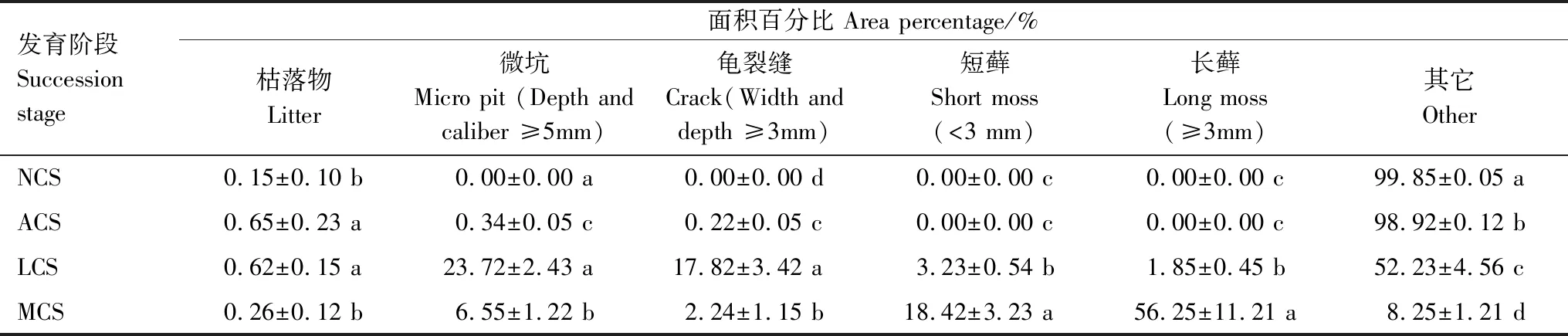
表2 生物土壤结皮发育过程中几种微区占比变化Table 2 Proportion changes of several typical microsites in the process of biological soil crust development
2.2 红砂种子在不同发育阶段结皮上的定居过程
自然条件下,红砂种子在不同发育阶段生物结皮上定居率存在显著差异(图3)。由图2可以看出,种子“落入”不同发育阶段生物土壤结皮上的第1天,地衣、藓类滞留的种子最多(约为75粒),两者之间差异不显著,其次为藻类,结皮前期最小(小于15粒);4个阶段滞留种子数两两之间差异显著,大小依次为:地衣>藓类>藻类>结皮前期。第7天时,4个阶段坡脚-坡顶区域滞留种子数均急剧减小,特别是结皮前期及坡顶的藻类结皮(无滞留种子);同一部位滞留的种子量大小与第1天一致,4者两两之间差异显著(坡顶的结皮前期和藻类除外)。第90天,种子在藻类、地衣、藓类结皮上滞留的种子实现成功定居,3者的定居率大小排序仍与第1天一致。
同样地,红砂种子在沙包不同部位的同阶段结皮上定居过程也存明显的差异。其中,结皮前期第1天的种子滞留数以下坡位最大,上坡位次之,坡顶、坡脚均显著小于前两者,7 d后所有区域的种子滞留数均为0;藻类阶段第1天坡顶种子滞留数显著小于其他3个部位,但其他3部位两两之间差异不显著;地衣阶段,不论是第1天还是更长时间,均以顶部滞留(定居)的种子量最小,短时间内(1—15 d)其他部分种子滞留数两两之间差异不显著,较长间后(90 d)下坡位的种子定居量显著大于坡脚或上坡位(坡脚与上坡位间差异不显著);藓类阶段第1天和第60天后的种子滞留数均以坡顶最小,并显著小于其他3个部位,其他时间4个部位种子滞留量两两之间差异不显著。
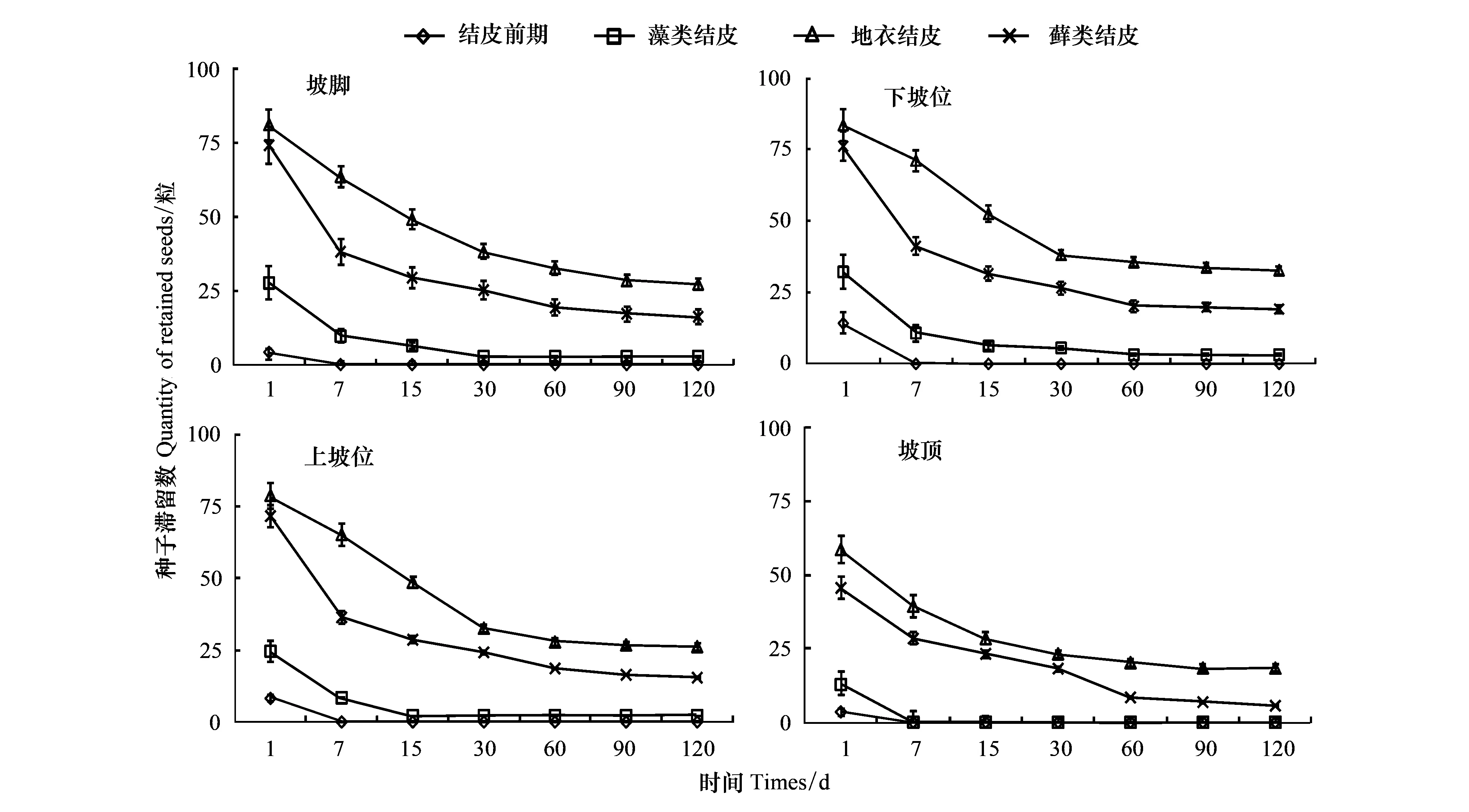
图3 红砂种子在不同发育阶段生物结皮上的定居过程Fig.3 Settlement process of R. soongorica seeds on biological soil crust at different developmental stages
2.3 定居种子在生物土壤结皮上的定居位置
在沙包迎风面坡脚、下坡位、上坡位、坡顶的不同发育阶段生物土壤结皮上,成功定居的种子着落微区特征见图4。可以看出,藻类结皮上的枯草丛是种子定居的唯一场所(图4)。在地衣结皮,种子主要定居于龟裂缝,特别是坡顶位置(高达95%),其次为微坑,枯草丛中定居数最小(坡顶为0);在同一位置(如坡脚、下坡位、上坡位或坡顶),3种微区内的种子定居数两两之间差异显著;随坡位上移,微坑和龟裂缝中定居种子数均先增大后减小(峰值均出现在下坡位),而枯落物中定居种子数持续减小。藓类结皮上定居的种子仅分布于龟裂缝和长藓株中,坡脚位置2者的定居种子差异不显著;随坡位逐渐升高,长藓上定居的种子数不断减小,但龟裂缝中定居的种子数呈现出先增大后减小,并显著大于相应位置长藓上的种子数。进一步对不同微区定居种子密度进行分析,发现等面积内龟裂缝中定居的种子数最多,分别是枯落物、微坑、长藓的2.5、12.5和18.0倍,4种微区种子密度两两之间存在显著差异性(微坑与长藓除外)(图5)。
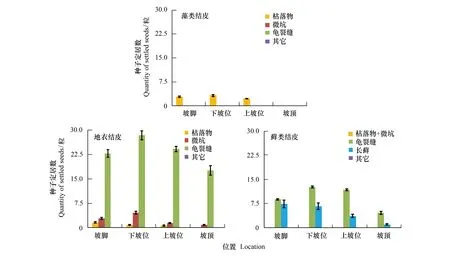
图4 红砂种子在生物结皮上宿存的位置Fig.4 Settlement location of R. soongorica seeds on the biological soil crusts
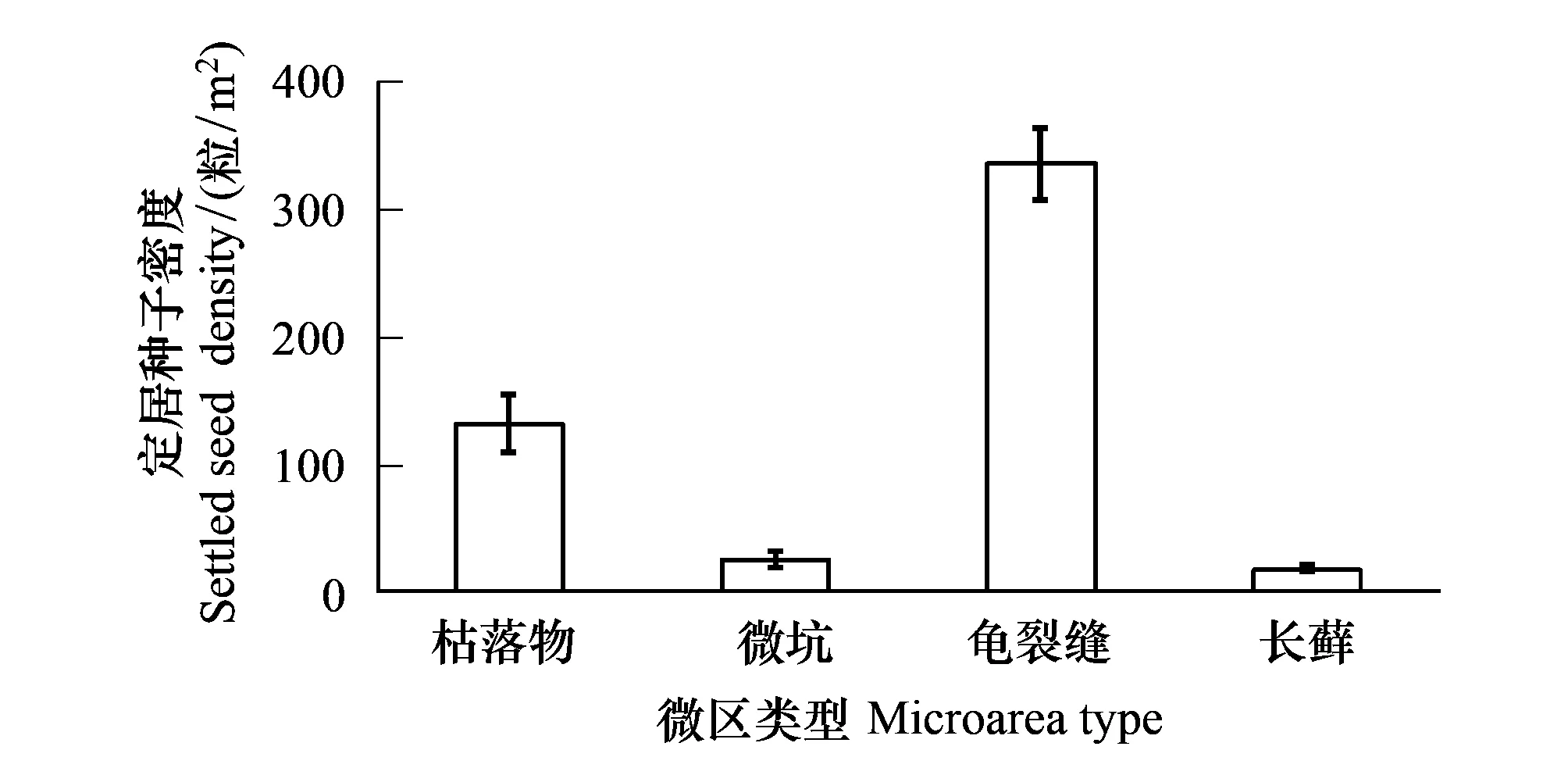
图5 四种微区内定居种子的密度Fig.5 Seed densities of settlement in the four micro areas
2.4 地表微区特征与种子定居的相关性
为了解生物土壤结皮上不用类型微区面积与种子定居的关系,对迎风坡下坡位样方中枯落物、微坑、龟裂缝、长藓上定居的种子数与其面积的关系进行了分析(图6)。由图6可以看出,生物土壤结皮上不同类型微区内定居种子数与其面积之间存在极显著的正相关关系,相关系数大小依次为:龟裂缝>枯落物>微坑>长藓;同时,4种微区内种子定居数与其面积间的关系均可由二次项表示(y=ax2+bx+c,a≠0),龟裂缝的拟合系数最大(0.962),其次依次为枯落物(0.945)、微坑(0.907)、长藓(0.762)。
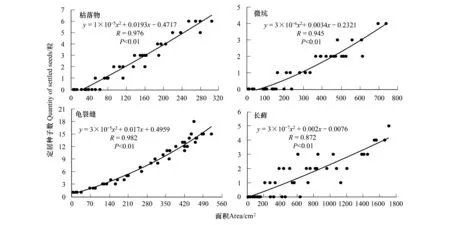
图6 不同类型微区面积与定居种子数的关系Fig.6 Relationships between the area and seed quantity in litter, micro pit, crack and long moss
3 讨论
种子传播是种子植物种群扩散的基础,其传播过程受多因素影响[25],因此传播机制存在多样化[26]。在干旱沙漠区,瘦小、轻质并附长毛/翅的种子借助风媒实现空中长距离传播及落地后的多次迁移[27—28]。本研究显示,红砂在不同发育阶段生物土壤结皮上均发生了一定程度的迁移,其迁移强度两两之间均存在差异性(图3)。比如,结皮前期85%以上的种子在第1天内发生迁移,第7天时样方内无滞留的种子;生物结皮上的种子在0—90 d内也发生了不同程度迁移,但迁移强度均显著小于结皮前期,尤其是地衣结皮和藓类结皮,120 d后其样方内仍有一定量的种子宿存。这说明生物结皮,特别是地衣和藓类结皮,不仅能有效减低风媒对红砂种子的迁移强度,并具有捕获红砂种子的能力,因此有利于红砂种子定居。前人在类似区域内对一些借助风媒传播的种子进行了研究,发现生物结皮对黄花补血草的捕捉率明显大于物理结皮[29],对霸王、沙拐枣的滞留率大于流沙[28],生物土壤结皮对红砂种子迁移的影响与这些研究相一致。
研究表明,易被风媒传播的植物种子着地后再次迁移过程与沙面的稳定性及地表微区特征密切相关,且地面越粗糙种子更容易定居[30—31]。在生物土壤结皮发育过程中,沙面逐渐固定并出现一定量枯落物、龟裂缝、微坑(结皮龟裂边缘翻卷、地衣发育、小型动物活动等形成)、苔藓等(表2),因此推测:不同发育阶段土壤结皮上红砂种子迁移过程的差异性与地表稳定性增强及以上几类微区出现有关。本文对不同发育阶段结皮上种子定居位置调查发现,红砂种子落地120 d时,藻类结皮上仅枯落物中有极少量红砂种子宿存(定居),地衣结皮上种子主要宿存于龟裂缝中(枯落物和微坑中也有少量),藓类结皮上的种子主要宿存于长藓株和龟裂缝中(图4),这进一步确定以上4种微区是影响不同生物土壤结皮上红砂定居过程及定居率的关键因子。此外,在沙包坡面不同位置上(坡脚、下坡位、上坡位或坡顶),风媒对红砂种子的迁移能力也存在明显的差异性,这一方面可能是不同位置坡度及风速不同引起的,比如,坡顶样方地势高、四周空旷、地表平坦,这使得风媒对种子干扰频度及强度均高于其它区域,所以种子更容易流失;另一方面是近地面植被盖度及其空间分布不同导致近地面风媒活动轨迹或强度不同,有待于进一步研究。
同一区位(即沙包迎风面下坡位)单位面积内枯落物、微坑、龟裂缝、长藓面积与其上定居种子数之间均存在极显著的正相关关系(图6),说明红砂种子在生物土壤结皮上定居能力由单位面积内不同微区面积组成决定。在试验中还发现,等面积枯落物、微坑、龟裂缝、长藓上定居种子数两两之间存现显著差异性,并以龟裂缝中宿存量最大,分别为枯落物、微坑、长藓的2.5、12.5和18.0倍(图5),这意味着龟裂缝更有利于红砂种子定居,其次依次为枯落物、微坑、长藓。尽管在结皮前期有枯落物存在,但无红砂种子定居,这主要是沙面未固定、枯落物面积太小(<0.2%),无法形成种子定居的理想场所。在藻类结皮向地衣、藓类结皮演替过程中,单位面积内4种微区总面积百分比持续增大,即:约1.2%、44.0%、65.3%,但红砂种子定居数呈现出先增大后减小的变化趋势(峰值出现在地衣阶段),导致两者变化不一致的主要原因为:地衣阶段单位面积内形成了大量龟裂缝(17.8%),所以捕获种子的能力大大提高;在藓类阶段龟裂缝面积百分急剧下降(仅为2.2%),长藓面积急剧增大(高达56.3%),而长藓对红砂种子捕获能力远远小于龟裂缝,因此单位面积内定居的种子远远少于地衣阶段。
基于以上研究与分析,本研究认为生物土壤结皮对红砂种子定居影响的作用机制主要为:①生物土壤结皮固定了沙面→提高枯落物宿存/滞留量→枯落物间的空隙为红砂种定居提供了适宜场所;②结皮层发生龟裂→裂缝边缘结皮层向上翻卷形成微坑并增大裂缝的宽度和深度,再加上地衣发育增大结皮表面的粗糙度→增强种子迁移难度并提供大量的宿存空间;③生物结皮发育到藓类阶段,大量龟裂缝及微坑被藓株“填平”,但长藓株之间的缝隙有利于红砂种子宿存。
4 结论
通过对干旱沙区地表藻类、地衣、藓类结皮上红砂种子的定居过程进行了研究,得出以下结论:
(1)藻类、地衣、藓类结皮均有效促进了红砂种子定居,并以沙包中下部的效果最佳,分别3.2%、32.8%和19.2%;定居种子主要宿存于地表枯落物、龟裂缝、微坑、长藓4类微区内,微区与定居种子数间存在显著的正相关关系,其对种子定居的正向作用大小依次为:龟裂缝>藓类>微坑>枯落物;等面积的以上4种微区,龟裂缝对种子捕获能力最大,约为枯落物、微坑、长藓的2.5,12.5和18.0倍。
(2)藻类结皮通过增强地表稳定性来降低了枯落物的流失量,进而促进种子定居宿存;地衣结皮层龟裂并翻卷导致地表出现大量宽而深的裂缝(≥3mm)及微坑(深、口径≥5mm),进而通过阻碍种子移动或直接捕获种子来促进种子定居;苔藓生长发育后,部分藓株间形成了大量空隙,一定大小的空隙(≥3mm)降低了种子迁移速度并促进其宿存。
