海上风电场建设与运行对渔业资源群落结构及空间分布的影响研究
2023-01-17汪润芝原峰李崇淑龙焰许清燕奚冰柔冼嘉俊叶锦韶
汪润芝原峰李崇淑龙焰许清燕奚冰柔冼嘉俊叶锦韶
(1.广东省海洋发展规划研究中心 广州 510220;2.暨南大学 广州 510632)
0 引言
发展海上风电与全球能源转型大势契合,与我国大力发展海洋经济、建设海洋强国和实现“双碳”目标的战略一致。广东省发展改革委《关于下达广东省2022年重点建设项目计划的通知》(粤发改重点〔2022〕157号)中强调2022年广东省重点建设的海上风电项目达14个,《广东省海洋经济发展“十四五”规划》(粤府办〔2021〕33号)中计划至2025年装机容量将达到1 800万k W。目前大规模发展海上风电对周边海域环境的影响尚不明确,海上风电场的建设与运行对周边海域渔业资源的影响是当前研究热点。
海上风电场建设阶段对渔业资源的主要影响因素包括打桩噪声和底质扰动[1]。水下冲击打桩噪声频域的能量分布与大黄鱼的听觉敏感频段相重叠,对大黄鱼听觉的影响较大[2];打桩噪声会轻微改变海域本土鱼类的运动行为,使位于打桩活动附近的大西洋鳕鱼(Atlantic cod)向风电桩基础附近的海床游动数米从而远离噪声源[3];打桩噪声对欧洲鲈鱼(Dicentrarchus labrax)具有声学胁迫,会直接改变鲈鱼的皮质醇含量水平[4];打桩时的声压级较高,会对周边海域栖息地的鱼类产生一定的干扰,使鱼群游得更浅[5]。海底电缆的电磁场和风机运行的叶片噪声是海上风电场运行阶段的主要环境影响因子。电磁场影响对电磁敏感的物种,可能会干扰敏感鱼类的迁徙与繁殖[6];室内模拟实验表明,电磁场会直接改变海洋生物的行为和存活率[7],如改变网箱内蓝鳍金枪鱼(Bluefin tuna)的游泳速度和倾斜方向[8],影响鱼类声音交流的方式和范围;风速和风力直接改变风机叶片的转速,当风速高于13 m/s时,运行中的风机叶片会直接吓跑鱼群[9]。海上风电场的建设和运行对海洋生物量、生物群落结构和群落多样性均可能产生影响[10],但同时有一定的积极效应,如风电桩基作为禁捕区可成为良好的庇护场所,桩基周围形成的新生境可能吸引其他适应性物种或种群[11-13],从而通过调节能量流动和食物营养结构而改变生物群落结构[14-16]。然而上述数据多数基于控制性实验研究,缺乏现场调查数据的支撑,且目前国内外关于海上风电场建设与运行对渔业资源群落结构和空间分布的影响研究仍较少。
珠海桂山风电场是广东首个海上风电示范项目,总装机容量为198 MW,分2期建成。项目Ⅰ期安装37台风机,已于2018年5月建成投产;项目Ⅱ期安装12台风机,已于2021年12月建成投产。珠海桂山岛海域拥有中华白海豚自然保护区,该风电场海域位于珠江河口的伶仃洋,而珠江河口是重要的鱼类繁育场[17]。因此,研究珠海桂山风电场的建设和运行对渔业资源的影响具有较强的代表性和重要的生态保护意义。
本研究于2021年7-12月在珠海桂山风电场海域开展现场调查,采用单船底拖网方式调查风电场建设与运行期间渔业资源的群落结构、优势种、群落多样性与空间分布特征,采用刺网调查结果验证风电桩基的人工鱼礁效应;结合项目论证期的历史数据,评估风电场建设与运行对渔业资源群落结构和空间分布的影响。研究结果有助于了解风电场建设期和运行期的渔业资源变化特征,为长期影响研究提供数据对比。
1 材料与方法
1.1 数据来源
游泳动物数据来源于7-9月和12月的单船底拖网调查,网具的网口目尺寸为50 mm,网囊目尺寸为20 mm,船速约为3 kn。共设置12个调查站位,分布于风电运行区(S5和S6)、风电建设区(S8和S9)、外围区(S4和S7)、上游区(S1、S2和S3)和下游区(S10、S11和S12),其中外围区、上游区和下游区均在风电场外(图1)。
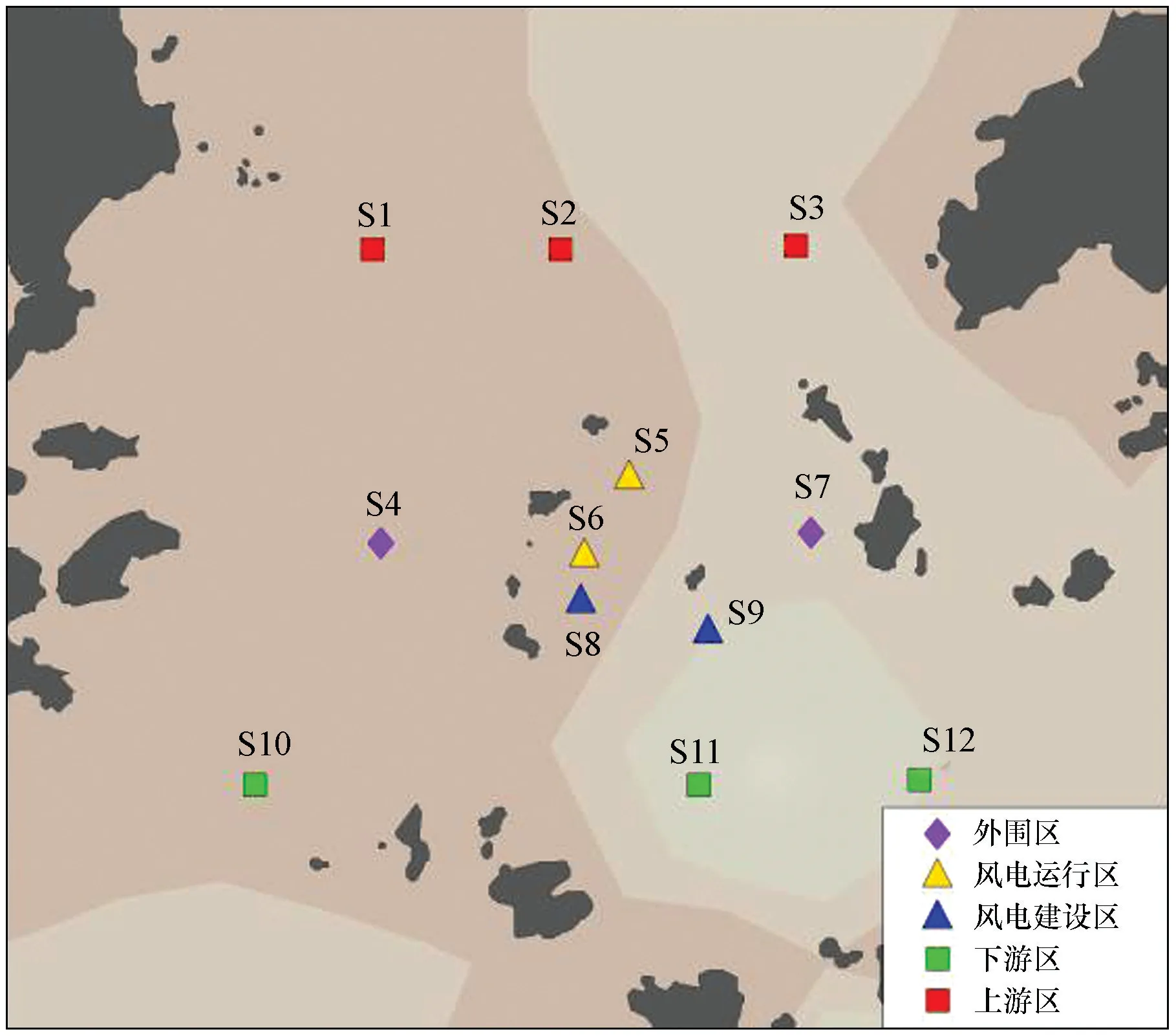
图1 调查站位
历史数据来源于海上风电工程论证期的现场调查(与本研究的调查方式相同,网具尺寸相近)。历史数据和现场调查数据的分析均按照《海洋监测规范》(GB 17378-2007)和《海洋调查规范:海洋生物调查》(GB/T 12763.6-2007)中规定的方法执行。现场对渔获物进行分类并记录种类,将样品冰冻保存带回实验室并测定生物学数据。此外,在12月的调查航次利用刺网在典型风电桩基附近设置调查站位,并在距离风电场内采样点2 000 m处设置对照站位,对周围水体进行渔业调查,用于分析风电桩基的鱼礁效应以及风电场的敏感物种和适应物种,刺网规格为长40 m、高1.2 m、网目2 cm。
1.2 指标选取与数据分析
生物多样性指标在生态环境监测和决策管理等方面具有量化指示作用。本研究的生物多样性指标用于指示渔业资源在海上风电场建设和运行各阶段不同层面的生物多样性变化趋势。基于现场调查数据,计算各航次渔业资源游泳动物的Shannon-Wiener指数(H')[18]、Pielou均匀度(J')[19]和丰富度指数(D)[20],评估风电场海域渔业资源的群落多样性。
各指标的计算公式为:
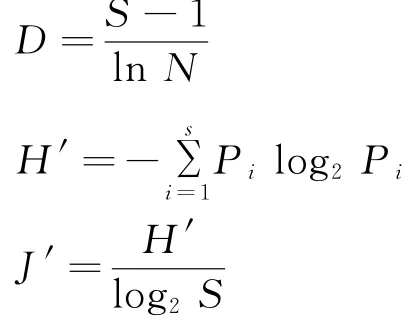
式中:N表示渔获物种的总个体数;S表示渔获物种的总种类数;P i表示第i种渔获物种的个体数占渔获物种总个体数的比重。
相对重要性指数(IRI)常用于评估生物群落的优势种成分。根据相关研究[21]计算游泳动物的相对重要性指数:
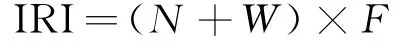
式中:N表示某种类渔获物种的个体数占渔获物种总个体数的比重;W表示某种类渔获物种的重量占渔获物种总重量的比重;F表示某种类渔获物种的出现频率。
定义IRI≥500的物种为优势种,100≤IRI<500的物种为常见种,10≤IRI<100的物种为一般种,IRI<10的物种为稀有种。
通过各航次的游泳动物种类数量计算更替率(A)[22],对比分析各航次物种的变化情况。更替率的计算公式为:
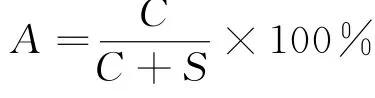
式中:C表示2个航次的种类增加和减少数量之和;S表示2个航次共有物种的种类数量。
资源密度(S)是表征种群状态的重要指标之一。根据底拖网扫海面积法,估算调查海域游泳动物的资源密度。资源密度的计算公式为:
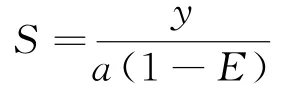
式中:y表示平均渔获率;a表示底拖网每小时的扫海面积(扫海宽度取浮纲长度的2/3);E表示逃逸率,取0.5。
2 结果与分析
2.1 渔业资源种类组成及空间分布
2021年7月共捕获游泳动物10目35科61属84种,其中包括鱼类48种、甲壳类34种和头足类2种,分别占本航次总种数的57.1%、40.5%和2.4%。8月共捕获游泳动物20目40科67属88种,其中包括鱼类52种、甲壳类34种和头足类2种,分别占本航次总种数的59.1%、38.6%和2.3%。9月共捕获游泳动物17目43科71属105种,其中包括鱼类73种、甲壳类26种和头足类6种,分别占本航次总种数的69.5%、24.8%和5.7%。12月共捕获游泳动物16目46科78属99种,其中包括鱼类57种、甲壳类39种和头足类3种,分别占本航次总种数的57.6%、39.4%和3.0%。
各航次和各站位鱼类、甲壳类和头足类的种类数量如图2所示,可以看出相同航次不同站位以及相同站位不同航次的渔获种类数量各异。
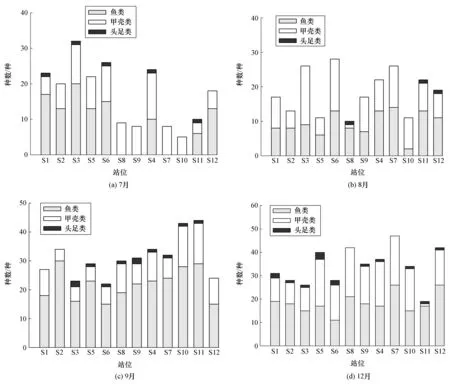
图2 各航次和各站位渔业资源的种类数量
2.2 物种更替率
2021年7-9月和12月的渔获种类数量分别为84种、88种、105种和99种,将两两航次的种类数量进行比较后计算更替率。结果显示:8月与7月、9月与8月以及12月与9月的物种更替率分别为57.5%、76.4%和93.4%;7月与9月、8月与12月以及7月与12月的物种更替率分别为41.0%、66.3%和58.1%。
2.3 主要优势种
4个航次的渔获种类数量由多到少依次为9月、12月、8月、7月,其中12月的常见种最多,7月的一般种最多,9月的优势种最多(图3)。
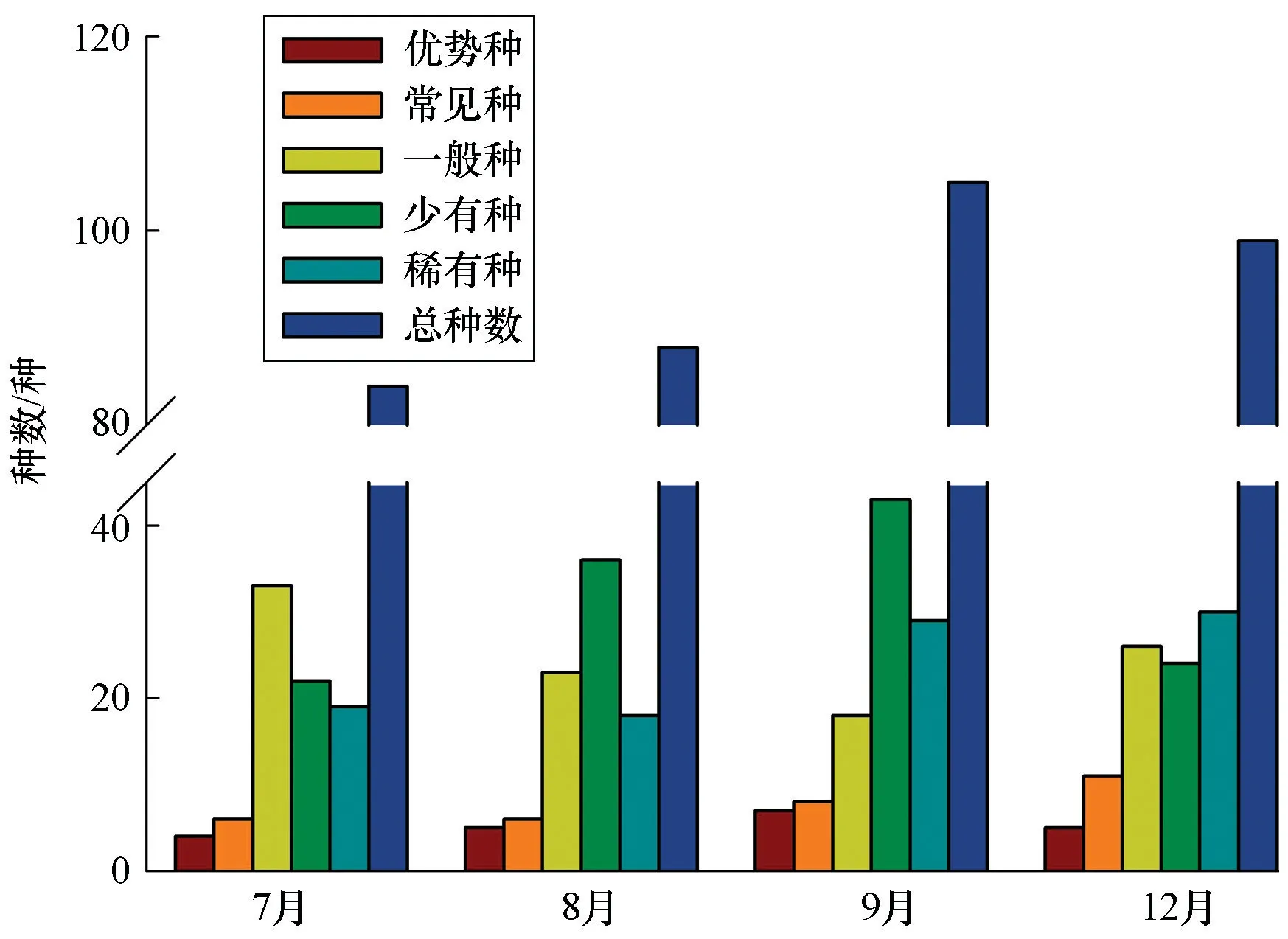
图3 各航次不同优势度的渔业资源种类数量
各航次的优势种信息如表1所示。
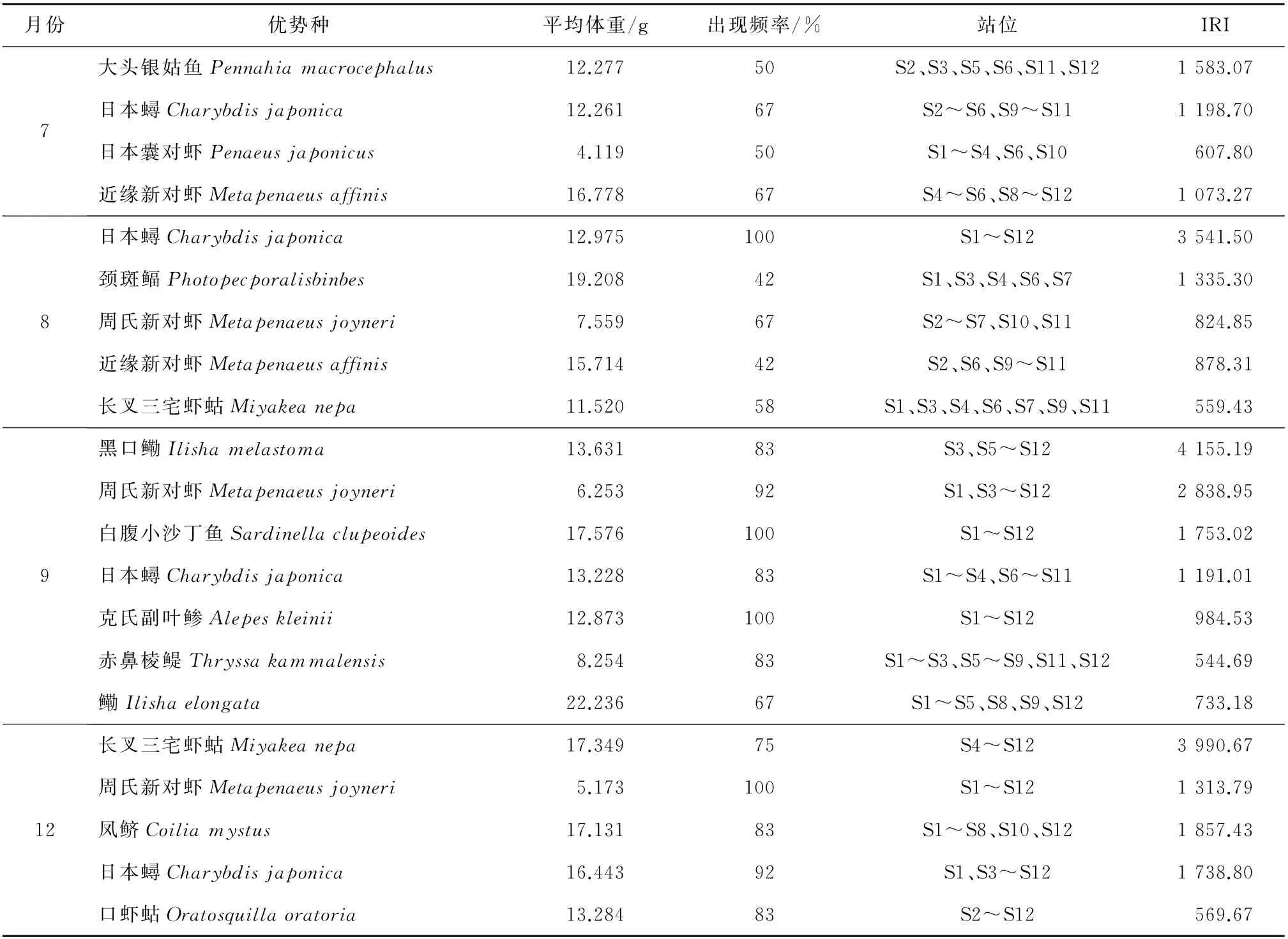
表1 各航次的优势种信息
日本蟳为各航次的共有优势种,IRI均超过1 000且8月最高,平均体重为12月最高、7月最低。7月的大头银姑鱼在调查海域的上游区、下游区和风电运行区出现。8月的颈斑鲾在调查海域的上游区、风电运行区和外围区出现。9月IRI最高的为黑口鳓,最低的为赤鼻棱鳀。12月IRI最高的为长叉三宅虾蛄,最低的为口虾蛄。长叉三宅虾蛄是8月和12月的共有优势种,但其在12月的平均体重高于8月。周氏新对虾为8月、9月和12月的共有优势种,但其在8月的平均体重最高。
2.4 群落多样性
各航次和各区域的渔业资源群落多样性指数如表2和图4所示。
表2 各航次渔业资源的群落多样性指数
图4 各区域渔业资源的群落多样性指数
表2显示,各类指数均为12月较高,且J'的差异较小。7月运行区的H'最高,下游区最低;8月建设区的H'最低,上游区和下游区相当;9月建设区的H'最低,下游区最高;12月上游区的H'高于下游区;7月和8月建设区的D均低于运行区,而9月和12月建设区的D高于运行区(图4)。
2.5 资源密度
调查海域各航次的渔业资源密度均存在不同程度的聚集性特征。7-9月和12月的平均资源密度分 别 为240.59 kg/km2、275.46 kg/km2、807.27 kg/km2和689.37 kg/km2,总体表现为9月最高、7月最低。从空间分布来看,7月上游区的S3略高于S2和S1,风电运行区和建设区的S5最高、S9最低,下游区的S11高于S10和S12;8月外围区的S7最高;9月上游区的S2和S3、风电运行区的S6和外围区的S7较高;12月上游区的S1和S2高于S3,风电运行区的S5较低,外围区的S7高于S4。
3 讨论
3.1 群落结构
在各秋季航次中,风电场项目Ⅰ期建设前(2012年)、项目Ⅰ期建设期(2017年)、项目Ⅰ期运行期(2018年)和项目Ⅱ期建设期(2021年)的渔业资源对比如表3所示。
表3 各秋季航次渔业资源的群落结构和多样性指数
项目Ⅰ期和Ⅱ期建设期的渔获物种数均少于建设前。2017年秋季的甲壳类种数最少,表明风电场建设施工对甲壳类的影响可能更明显;2021年9月鱼类种数恢复至73种,甲壳类种数恢复不明显,头足类种数恢复至6种;尽管群落多样性有所恢复,但与风电场建设前相比仍较低。因此,风电场建设和运行在短期内会改变渔业资源群落结构,但该影响随着时间的延长而逐渐减弱。在风电场建设期结束后,风电场运行成为调查海域的主要影响因素。风电场内的风电桩基在一定程度上改变生物的食物结构和栖息地等生境条件[15],可能有利于吸引适应性物种,从而改变渔业资源群落结构。
风电场建设后(2021年秋季)与建设前(2012年秋季)相比的渔获物种更替率为31.7%,小于2021年任意2个航次的物种更替率(41.0%~93.4%),表明2021年各航次的月份差异对调查海域渔获物种结构的影响可能大于风电场建设和运行的影响。风电桩基为各类附着生物提供栖息地,而桩基作为人工鱼礁可能通过聚集效应改变食物网而影响能量流动,从而调节渔获物种的群落结构[16]。
海水温度和盐度也会直接影响鱼类种数[23-24]。根据现有文献报道,2009年8月至2010年4月的调查结果表明温度是珠江口伶仃洋海域游泳动物种类和数量发生变化的主要影响因子[25]。本研究4个航次的海水温度有明显差异(数据未展示),进一步表明季节(温度)变化是调查海域渔业资源群落结构的主要影响因子之一。
3.2 优势种
风电场建设前(2012年秋季)调查海域的优势种有8种,按IRI由大到小分别为棘头梅童鱼(Collichthys lucidus)、凤鲚、周氏新对虾、近缘新对虾、黑斑口虾蛄(Oratosquilla kempi)、四指马鲅(Eleutheronema tetradactylum)、中华管鞭虾(Solenocera crassicornis)和六指马鲅(Polynemus sextarius)。项目Ⅰ期建设期(2017年秋季)的优势种有6种,包括凤鲚、棘头梅童鱼、鹿斑鲾(Secutor ruconius)、断脊口虾蛄(Oratosquillina interrupta)、哈氏仿对虾(Parapenaeopsis hardwickii)和鳓,与建设前相比仅有凤鲚和棘头梅童鱼为共有优势种。项目Ⅱ期建设期(2021年秋季)的优势种恢复到7种(表1),与建设前相比仅有周氏新对虾为共有优势种。珠江河口段棘头梅童鱼的摄食等级受季节影响,各季节水域环境差异显著[26]。2021年的调查结果显示风电场在不同建设和运行阶段导致的水域环境差异在一定程度上改变优势种组成,进一步证明风电场建设和运行会改变渔业资源群落结构[27]。日本蟳为2021年各航次的共有优势种(表1),这与风电场建设前(2009年8月至2010年4月)日本蟳为珠江口伶仃洋海域的全年优势种相一致[25],表明风电场建设对日本蟳生态位的影响可能不明显;日本蟳作为甲壳类生物,其较高的优势度也在相关研究[16]中得到证实。
根据2021年12月的刺网调查结果,从渔业资源物种个体数量的角度判断,风电桩基具有一定的人工鱼礁效应。隆线强蟹(Eucrata crenata)、口虾蛄和舌鳎(Soleidae)均聚集于风电桩基周边水体,由此推断风电桩基附近的新栖息环境可能吸引这些物种。疣尾小绿虾蛄(Odontodactylus scyllarus)和窝纹网虾蛄(Dictyosquilla foveolata)仅出现在风电场外的对照站位,表明这些物种可能对风电场内的水下环境较敏感。风电桩基形成的人工鱼礁会改变礁区周边水体的水动力、能量流动和营养结构,形成的新栖息地会直接调节鱼类的优势种组成和群落结构[12-13,15,28],结合渔业资源群落结构同时受潮流和物种生活习性的影响[29],推断风电桩基形成的人工鱼礁效应可能同时受多种因子的协调作用,促使某些适应性物种在此聚集。
3.3 物种数量
根据风电场建设的实际情况,2021年各航次采样期间均无水下打桩施工,且8-9月已临近建设期尾声,12月已进入全面运行阶段。7月上游区的鱼类种数多于风电运行区,这可能源于夏季丰水期从近岸径流汇入丰富的营养物质从而调整食物结构;风电建设区和外围区均只监测到甲壳类,表明鱼类和头足类可能对风电施工更加敏感[30]。8月和9月各站位的鱼类种数无明显规律。12月风电建设区和运行区以及外围区的渔获种数高于上、下游区,可能主要得益于风电桩基形成的人工鱼礁效应[31-32],这与苏文等[33]的研究结果一致。风电桩基周围的物种数量多且丰度高[15,34],为渔业生物提供丰富的食物来源,同时为渔业生物种类数量和结构的改变提供能量基础。
3.4 群落多样性
根据2021年各站位的多样性指数(图4),各航次风电运行区的Shannon-Wiener指数均较高,进一步证明风电桩基具有人工鱼礁效应。除7月外,其他航次风电运行区的Pielou均匀度均高于外围区和建设区。7月风电建设区仅监测到甲壳类,因此该航次的丰富度指数偏低;相同航次不同站位的丰富度指数差异不显著,但不同航次相同站位的丰富度指数差异显著,表明2021年各航次季节差异对渔业资源的影响比空间差异更显著。
3.5 资源密度
根据历史数据和调查数据,各秋季航次的渔业资源密度如表4所示。
表4 各秋季航次的渔业资源密度
表4显示,项目Ⅰ期(2017年秋季)的平均资源密度低于风电场建设前,表明风电场建设活动降低海域的渔业资源量;而项目Ⅰ期运行阶段(2018年秋季)和Ⅱ期建设阶段(2021年秋季)的平均资源密度均高于风电场建设前,且2018年秋季是建设前的2.49倍,这主要得益于2018年秋季甲壳类的资源密度较高。相似的结果也出现在运行2年后的德国北海贝特风电场[35]。
为进一步分析风电场对渔业资源密度的影响,结合历史数据,对风电场内、风电场外和风电场分区进行对比分析。2012年11月风电场内的渔业资源尾数密度低于风电场外,而2013年3月则相反;2021年7月和8月风电场内、外的渔业资源尾数密度相当,可能由于7月和8月同属夏季,海水温度和盐度的差别较小,且均处于风电场建设期;7月风电运行区的渔业资源尾数密度最高,而风电建设区最低,考虑是因为建设期的噪声驱赶对声音敏感的物种[26];8月、9月和12月风电建设区的渔业资源密度均高于风电运行区,外围区的渔业资源密度均高于风电运行区,而上、下游区的渔业资源密度则无明显规律。这些结果表明调查海域的渔业资源密度可能受风电场建设和运行、站位地理位置以及调查月份等因素的综合影响。
对相同季节进行比较可知,风电场建设后(2021年秋季)调查海域的渔业资源尾数密度和资源密度均高于风电场建设前(2012年秋季),在江苏省如东县的海上风电场也有相似的研究结果[15]。在风电场建设前的2012年秋季和2013年春季,风电场内的渔业资源密度均高于风电场外。2021年7月和8月风电运行区的渔业资源密度均低于风电场外,但尾数密度差别不大,推测主要由物种个体重量的差异导致。2021年9月风电场内的渔业资源尾数密度和资源密度均高于风电场外,而12月风电场外的渔业资源尾数密度和资源密度均高于风电场内,这种差异可能源于季节的影响。
4 结语
根据2021年7-9月和12月的渔业资源游泳动物调查结果,渔获种类主要为鱼类和甲壳类,其中9月的渔获种数最多。与历史调查数据的对比结果表明,海上风电场建设对渔业资源具有短期影响,即在短期内会改变游泳动物的群落结构和优势种组成,同时降低群落多样性和资源密度。风电场运行后渔业资源群落多样性和资源密度有所恢复,风电桩基具有一定的人工鱼礁效应。风电场海域的渔业资源群落结构和群落多样性受季节的影响大于风电场的影响。调查海域的渔业资源可能受风电场建设和运行、站位地理位置以及调查月份等因素的综合影响。
