2019—2020山东小麦区试品系55个基因的等位基因分布
2023-01-14宋国琦李玉莲程敦公李豪圣李根英
李 玮 张 敏 宋国琦 李玉莲 高 洁 程敦公 李豪圣,* 李根英,*
(1 山东省农业科学院作物研究所/农业农村部黄淮北部小麦生物学与遗传育种重点实验室/小麦玉米国家工程实验室,山东 济南 250100;2 青岛农业大学农学院,山东 青岛 266109)
小麦(Triticum aestivumL.)是我国三大粮食作物之一,同时是我国北方最重要的口粮作物[1]。不断培育高产优质的小麦品种是小麦生产的基础,对增强种业竞争力,保障我国粮食安全具有十分重要的意义。山东是全国小麦生产第二大省,据山东统计年鉴数据显示,2019年山东省小麦播种面积400.18 万公顷,总产量255.29 万吨[2]。优良品种是保障小麦生产的基础,山东省小麦品种审定区域试验代表了山东小麦育种的最新进展和育种方向。对区试品系进行分析,在了解山东小麦育种特点和变化基础上,可以为品种改良提供参考。
区试参试品系常被育种家作为亲本材料进行杂交组配,前人对区试品系的研究较多,对河北、河南、山东和四川4 个省份的75 份小麦区试品系和78 份国家冬小麦区试品系的抗条锈病鉴定结果表明,四川省区试品系条锈病抗性较好,其他区试品系相对较差[3-4],可能与四川省条锈病自然发病较重,而北方省份自然发病较轻有关。对山东省和河南小麦区试品系的品质性状进行研究,发现达到国家优质强筋标准的品系较少,主要限制指标是稳定时间[5-6]。王步云等[3]研究认为河北、河南、山东小麦区试品系的遗传相似性较高;宋晓霞等[7]也认为黄淮南片小麦区试品系遗传基础狭窄;而高晓慧等[8]分析表明黄淮南片麦区78 个小麦品系遗传距离变幅较大,可能与所选材料不同有关。对国家区试参试品系产量相关性状研究表明,品系年际间产量变幅大,整体稳步增加,生育期和千粒重与产量关联度最大[7,9-10]。以上研究主要集中在产量、品质、抗病性等表型性状,对品系所携带的优异等位基因关注相对较少。
2016年Rasheed 等[11]发表了70 个小麦竞争性等位基因特异聚合酶链式反应(kompetitive allele specific polymerase chain reaction,KASP)功能标记,涉及小麦适应性、产量、品质和抗逆性等性状,对之前小麦中已开发KASP 功能标记进行了系统总结。2019年Khalid等[12]报道小麦KASP 功能标记的数量已增加至124 个,为小麦功能基因的鉴定和育种奠定了基础。随后KASP 标记逐渐在我国开始应用。邹景伟等[13]对120 份小麦品种(系)的株高等15 个基因进行了KASP标记检测,认为对常规手段育成的高代品系进行KASP标记辅助选择可作为一种重要的分子育种策略。单子龙等[14]、王志伟等[15]和Zhang 等[16]分别对河北、云南、宁夏的小麦品种(系)进行了KASP 标记检测,结果表明不同基因的优异等位变异占比差异较大,明确了优异等位基因的载体材料,为各省小麦品种改良提供了参考。Zhao等[17]对来自全球的1 152份小麦材料的47个基因进行了KASP 标记分析,明确了已被育种选择的36 个优异等位基因,促进了小麦分子育种发展。上述前人研究均主要关注小麦品种,但对区试品系的分析较少。
为了明确已知功能基因在区试品系中利用情况,并为亲本选配和后代辅助选择提供参考,本研究对2019—2020年山东省区试高产组和强筋组的品系进行64 个KASP 标记的分子检测,旨在为利用分子标记辅助选择,为促进小麦精准化育种,提高小麦育种效率奠定理论基础。
1 材料与方法
1.1 试验材料
试验材料为2019—2020年山东省小麦区域试验参试品系,共计126 份(表1),其中高产组111 份(含对照品种济麦22,高产区试品系编号前两位为GQ),强筋专用组15 份(含对照品种济南17,强筋区试品系编号前两位为QQ)。上述材料均于2019年11月播种于山东省农业科学院济南试验基地。
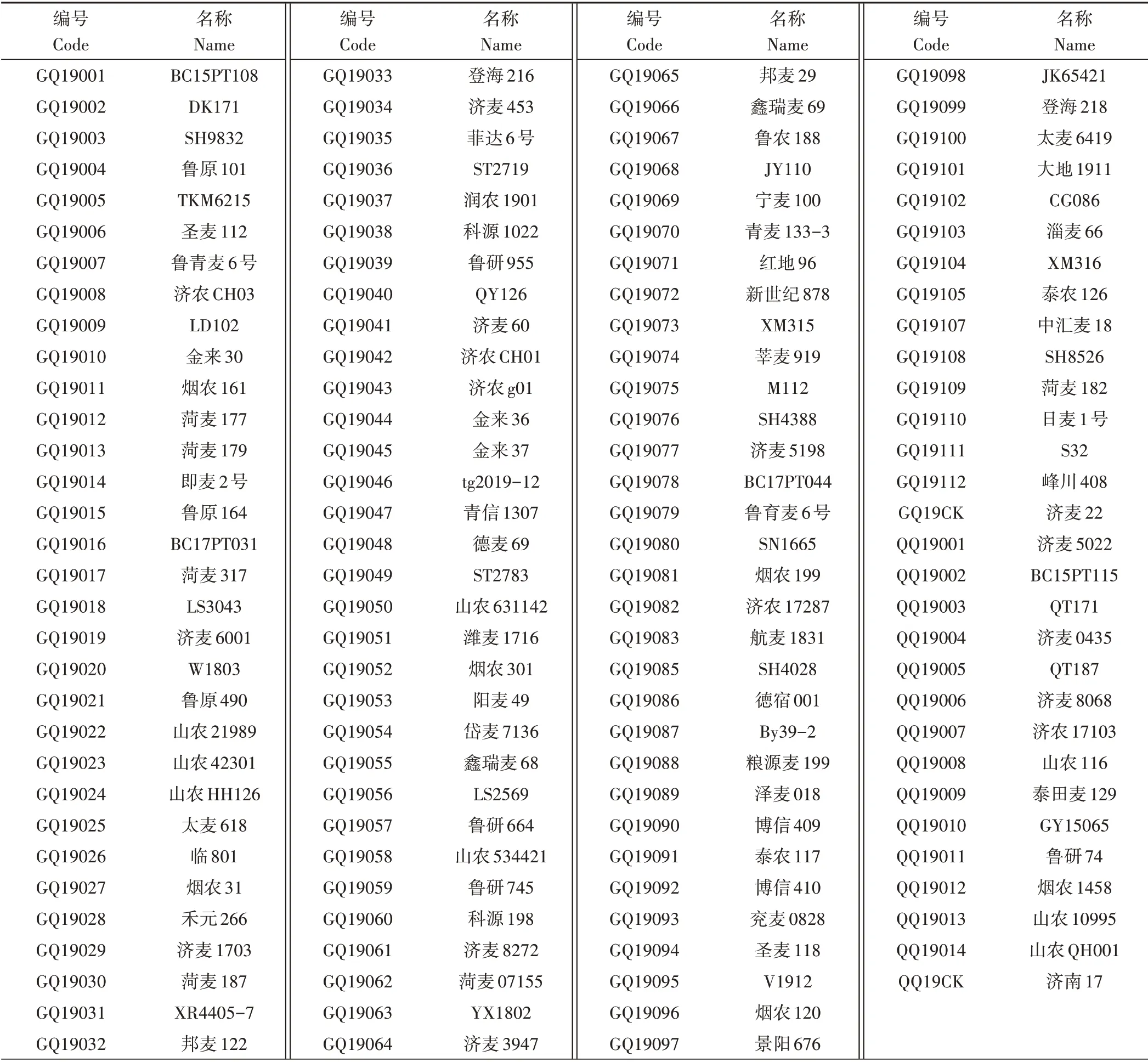
表1 参试品系Table 1 Tested wheat lines
1.2 试验方法
1.2.1 DNA 提取 采用简化的高盐低pH 法,试验步骤参考文献[18]。
1.2.2 KASP 标记检测与分析 KASP 标记检测反应体系、程序参考高洁等[18]的方法。KASP 标记名称、来源和对应基因详见表2,引物由青岛擎科生物技术有限公司合成。KASP 反应结束后用PHERAstar 荧光读板仪(英国LGC 公司)检测荧光信号,然后导入仪器自带的Kluster Caller软件进行分型。
1.2.3 优异等位基因 大光周期基因和春化基因的优异等位基因和生态区有关,本研究中将所检测材料中光周期和春化基因中占优势的等位基因作为优异等位基因,其他基因的优异等位基因在分子标记开发时已给出。详见表2。
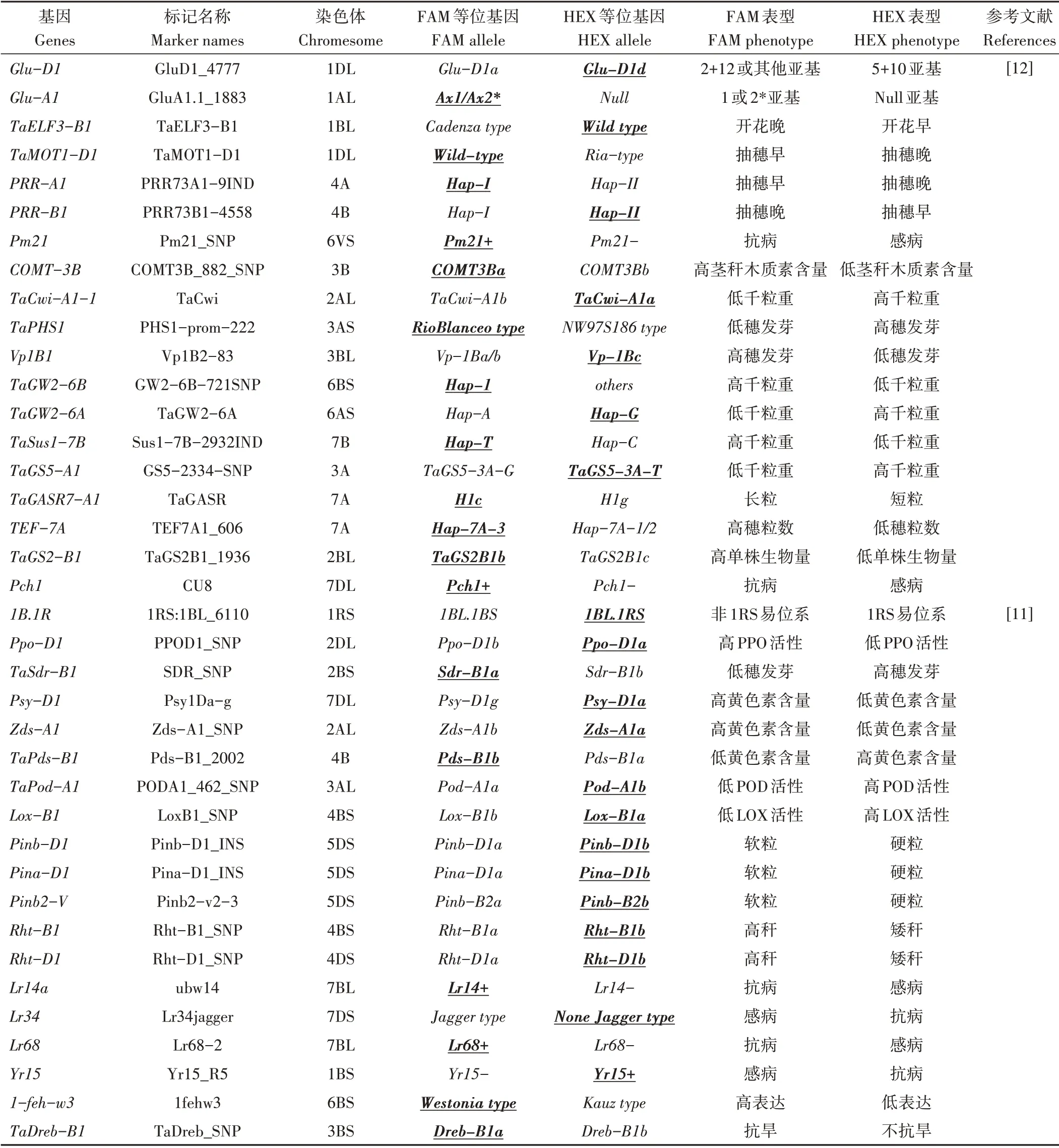
表2 KASP标记信息Table 2 KASP marker details
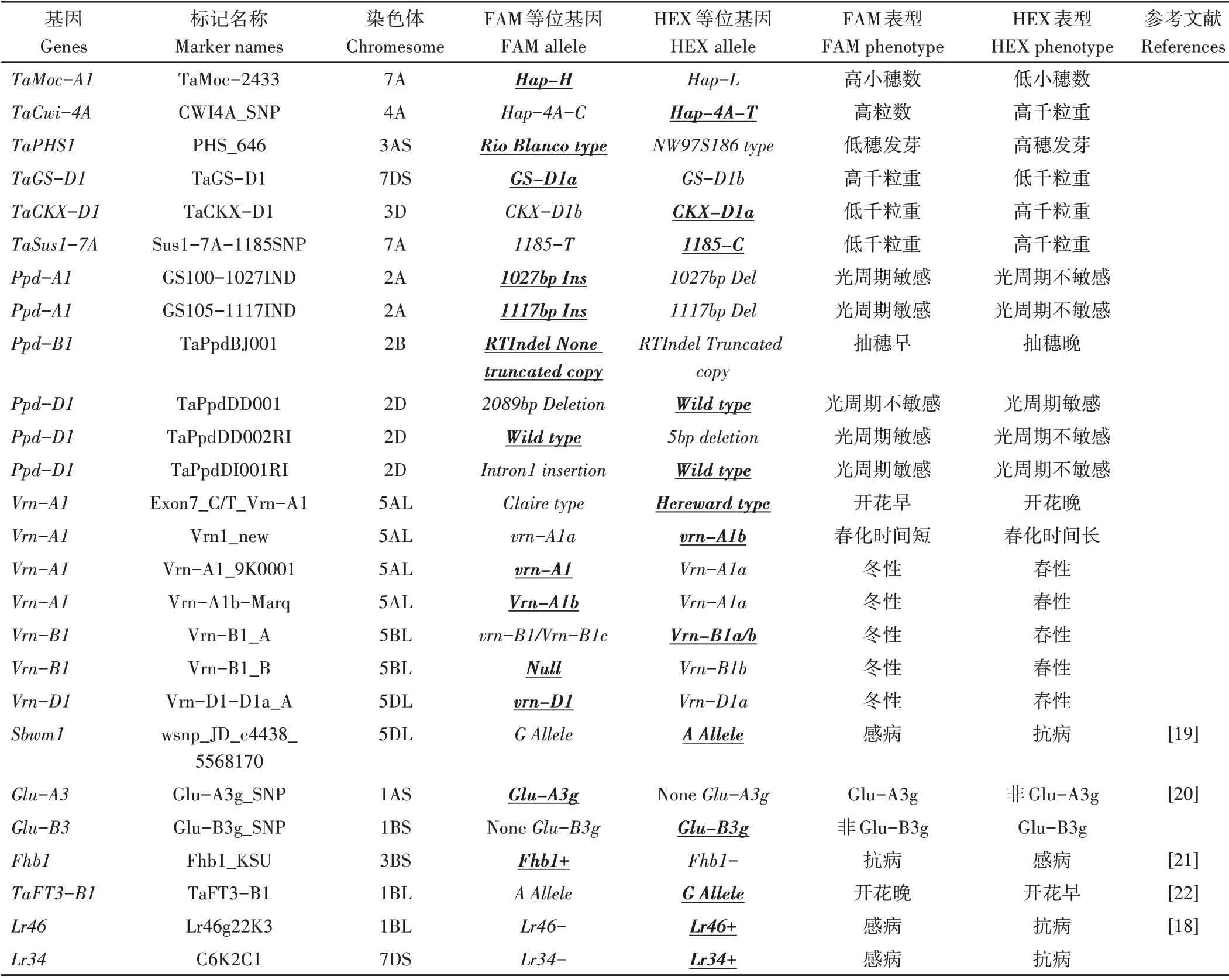
表2(续)
2 结果与分析
2.1 KASP标记检测概况
本研究共检测了64 个KASP 标记,其中43 个标记分型效果较好,可以将全部样品明显分成蓝色(FAM)和红色(HEX)两种基因型,相同基因型内个体较集中(图1-a)。15个标记分型效果尚可,虽可将待测样品分成两种基因型,但相同基因型内个体较分散,基因型间距离较近(图1-b)。6个标记分型存在问题,其中GluA1.1_1883、PPOD1_SNP、PODA1_462_SNP、COMT3B_882_SNP和TaGS-D1 5个标记检测到较多的杂合基因型(绿色)与高代品系多为纯合的结果不符(图1-c),TaGS2B1_1936有41个未知基因型数据点(粉色),推测与引物结合位点存在变异有关(图1-d)。
本研究中单个标记的缺失数据点一般为0~3 个,除TaGS2B1_1936 外,平均每个标记的缺失数据点为1.1 个。共有20 个标记全部样品均为一种等位基因,主要涉及抗病、千粒重、春化和光周期相关基因。其他大部分标记的等位基因呈两极分化,以其中一种等位变异占多数(图2)。
2.2 产量相关基因检测结果
1RS:1BL_6110 用于检测1B/1R 易位,1B/1R 易位系共有13 份(占比10.32%),这些品系均来自高产组。Rht-B1_SNP 和Rht-D1_SNP 用于检测株高基因,有120份携带Rht-D1b矮秆等位基因,占比95.24%,其他4.76%(6份)为Rht-B1b矮秆等位基因。与千粒重相关的标记共10 个。TaGASR7-A1 H1c长粒等位基因占比100%,TaCwi-A1a、TaGW2-6B Hap-1、TaSus1-7B Hap-T高千粒重等位基因占比较高,分别是88.89%、100.00%和97.62%,其次是TaGW2-6A Hap-A、TaSus1-7A 1185-C高千粒重等位基因分别占比38.10%、41.27%,TaCwi-4A Hap-4A-T、TaGS5-3A-T高千粒重等位基因占比较低,分别为12.70%和13.49%,仅济农CH03携带CKXD1a高千粒重等位基因,占比为0.79%,TaGS-D1 仅检测到杂合和GS-D1b两种基因型,GS-D1a高千粒重等位基因未检测到。与穗粒数相关的标记有2个,TEF7A1_606Hap-7A-3高穗粒数等位基因占比仅为18.25%。未检测到TaMoc-7A的Hap-H多小穗数等位基因。与生物量相关的标记有1 个,为TaGS2B1_1936。TaGS2B1_1936 的检测结果中未知基因型比例较高(图1-d),TaGS2B1b高生物产量等位基因占比60.32%。
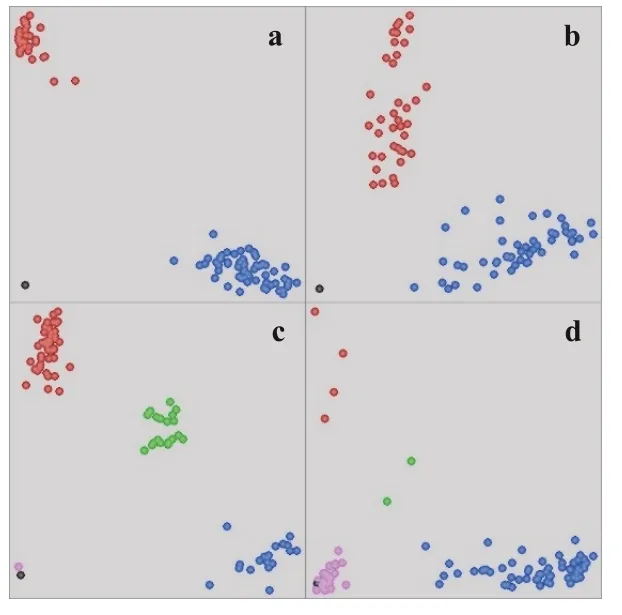
图1 KASP标记分型图Fig.1 Genotyping diagram of KASP markers
从以上结果可以看出,参试品系主要利用的矮秆基因为Rht-D1b,与产量相关的基因除TaCwi-A1-1、TaSus1-7B、TaGW2-6B外,其他基因的利用率都不高。
2.3 品质相关基因检测结果
结果如图2所示,GluA1.1-1883 和Glu-D1_4777用于检测高分子量麦谷蛋白亚基,优异等位基因Ax1/Ax2*和Glu-D1d分别占比42.86%和38.89%,如果仅统计强筋专用组的参试品系,占比分别为66.67%和80.00%。Glu-A3g_SNP 和Glu-B3g_SNP 用于检测低分子量麦谷蛋白,低分子量谷蛋白优异等位基因Glu-A3g和Glu-B3g分别占比96.03%和15.08%。Psy1Dag、Pds-B1_2002 和Zds-A1_SNP 与黄色素含量相关,低黄色素含量等位基因有利于增加面粉白度,低黄色素含量Psy-D1a、Pds-B1b和Zds-A1a等位基因占比分别为96.03%、18.25%和19.84%。LoxB1_SNP、PODA1_462_SNP 和PPOD1_SNP 与面粉及其制品白度相关。高活性Lox-B1a等位基因和高活性Pod-A1b等位基因分别占比53.17%、56.35%。多酚氧化酶(polyphenol oxidase,PPO)低活性Ppo-D1a等位基因占比22.22%。Pina-D1_INS、Pinb-D1_INS 和Pinb2-v2-3 与籽粒硬度相关,硬质等位基因Pinb-D1b、Pina-D1b和Pinb-B2b占比分别为2.38%、89.68%和78.57%。参试品系所含硬度基因以Pinb-D1位点为主。
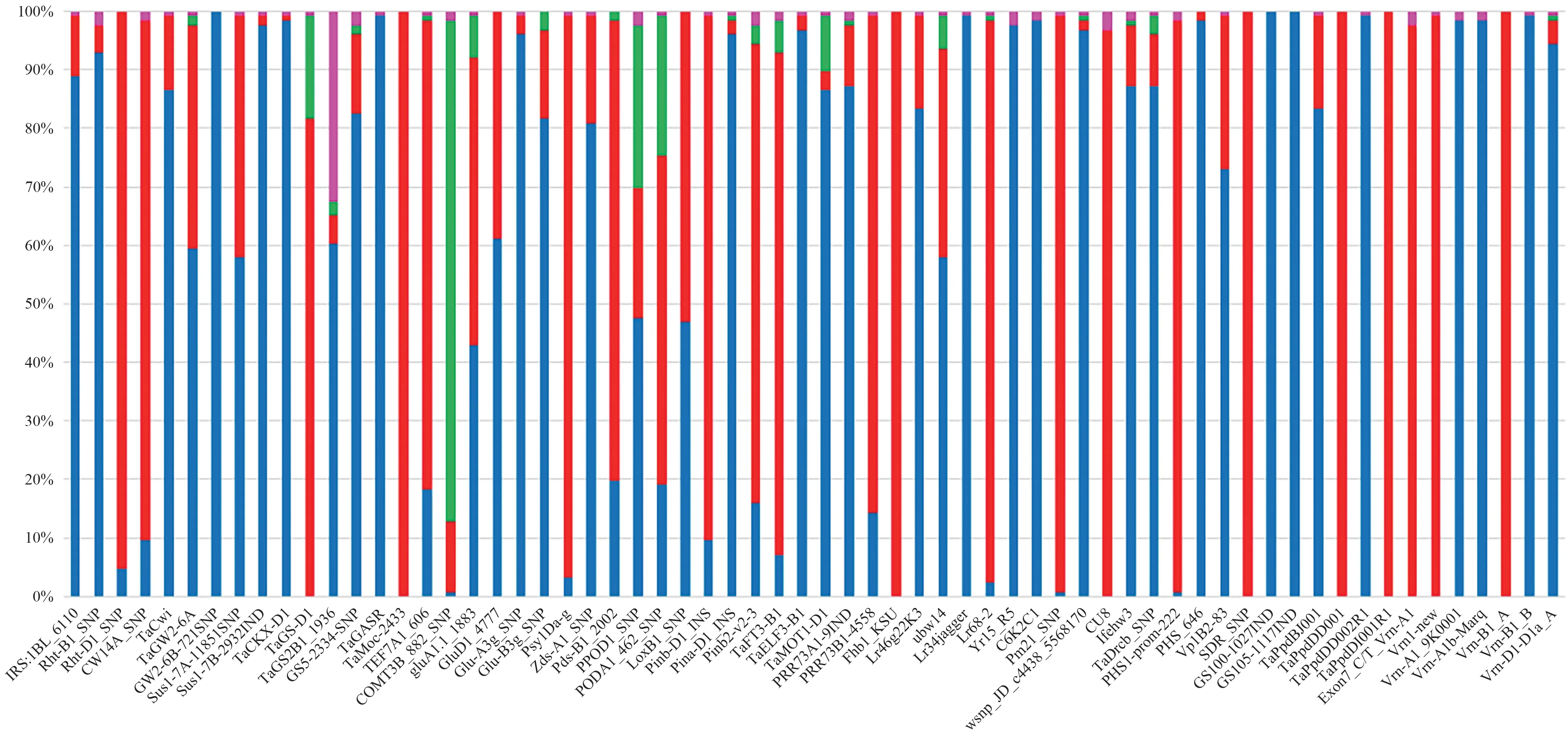
图2 等位基因占比Fig.2 Allele percentage
2.4 抗性相关基因检测结果
Fhb1_KSU 与抗赤霉病有关,所有参试品系均为Fhb1-感病等位基因。Lr14a抗叶锈病等位基因Lr14+占比57.94%,检测Lr34的标记有Lr34jagger 和C6K2C1,2 个标记的抗病等位基因占比均为0,故抗病等位基因Lr34+未检测到,Lr46抗病等位基因Lr46+占比15.87%,岱麦7136、济麦60 和鲁研74 三个品系携带Lr68抗病等位基因Lr68+,占比2.38%。CU8与眼斑病有关[12],未检测到抗病等位基因Pch1+。Pm21_SNP与白粉病有关[12],仅CG086 为抗病等位基因Pm21+,占比0.79%。wsnp_JD_c4438_5568170 与小麦黄花叶病有关[19],M112 和济麦8068 携带抗病等位基因Sbwm1-A,占比1.59%。Yr15_R5 是抗条锈基因Yr15的连锁标记,抗病等位基因Yr15+未检测到。以上结果表明,与抗病相关的已知基因位点在参试品系中含量较低。1fehw3 和TaDreb_SNP 与干旱条件下的籽粒产量有关,高表达等位基因Westonia type和高籽粒产量等位基因Dreb-B1a占比均为87.30%。PHS1-prom-222、PHS_646、SDR_SNP、Vp1B2-83 与穗发芽有关,PHS1-prom-222 和PHS_646 均是TaPHS1的标记,抗穗发芽等位基因RioBlanceo type、RioBlanceo type、Sdr-B1a、Vp-1Bc分别占比0.79%、98.41%、0.00% 和26.19%,表明部分抗穗发芽基因的利用率较低。COMT3B_882_SNP 与茎秆木质素含量有关,该标记存在杂合基因型多的问题,参试品系中仅有1份携带高茎秆木质素含量纯合等位基因COMT3Ba,为济农CH01。
2.5 开花相关基因检测结果
GS100-1027IND、GS105-1117IND、TaPpdBJ001、TaPpdDD001、TaPpdDD002RI、TaPpdDI001RI与光周期相关,GS100-1027IND 和GS105-1117IND 均是Ppd-A1的标记,TaPpdDD001、TaPpdDD002RI和TaPpdDI001RI均是Ppd-D1的标记。除TaPpdBJ001(Ppd-B1)有15.87%为晚抽穗等位基因RTIndel Truncated copy外,其他位点参试品系均为相同等位基因。Exon7_C/T_Vrn-A1、Vrn1_new、Vrn-A1_9K0001、Vrn-A1b-Marq均是春化基因Vrn-A1的标记,Vrn-B1_A 和Vrn-B1_B均是春化基因Vrn-B1的标记,Vrn-D1-D1a_A 是春化基因Vrn-D1的标记。除Vrn-D1-D1a_A(Vrn-D1)有3.97%为Vrn-D1a春性等位基因外,其他位点参试品系均为相同等位基因。PRR73A1-9IND、PRR73B1-4558、TaELF3-B1、TaFT3-B1、TaMOT1-D1与开花期相关,除TaELF3-B1 Wild type早花等位基因占比仅为2.38%外,PRR-A1 Hap-I、PRR-B1 Hap-II、TaFT3-B1 G Allele、TaMOT1-D1 Wild-type早花等位基因占比均较高,分别为87.30%、84.92%、85.71%、86.51%。携带TaELF3-B1 Wild type早花等位基因的品系有S32、菏麦182和山农116。
3 讨论
3.1 区试品系优异等位基因利用情况
1B/1R 易位系具有高产、抗病等优点,曾作为优异种质资源在世界范围内被广泛利用[13,23]。20世纪70年代1B/1R 易位系被引入中国,1980年丰抗2 号等第一批育成品种开始应用于生产[24-25]。王晓军等[24]对1980—2008年间黄淮麦区育成品种(系)进行了分子标记检测,发现66份大面积推广品种中45.5%携带1B/1R易位。2020年宫文萍等[26]对来自全国的1 293份小麦品种(系)进行分子检测,发现542份能扩增出1B/1R 易位系特异条带,占比41.92%,表明1B/1R 易位系在黄淮麦区乃至全国被广泛应用。本研究中仅高产组有13 份品系为1B/1R 易位系,而强筋专用组未检测到。由此推测出由于受1RS上携带的抗病基因丧失抗性和黑麦碱对小麦加工品质不利影响,1B/1R 易位系在高产育种中利用率逐步降低,在优质育种中极少使用。李韬等[27]研究认为1RS 可能携带赤霉病扩展抗性相关基因,与Fhb1基因有累加效应,1B/1R易位系在今后高产育种中可能仍有利用价值。矮秆基因被称为“绿色革命”基因,是高产育种的关键,目前生产上主要利用Rht-B1b和Rht-D1b[13,28]。本研究中参试品系携带的矮种基因以Rht-D1b矮秆等位基因为主,与前人的研究结果一致。产量三要素中千粒重相关基因被报道最多,本研究选用的相关标记有10 个,其中TaGASR7-A1 H1c、TaCwi-A1a、TaGW2-6B Hap-1、TaSus1-7B Hap-T优异等位基因的利用率较高(>85%);TaGW2-6A Hap-A和TaSus1-7A 1185-C利用率略f低,在40%左右;TaGS5-3A-T、TaCwi-4A Hap-4A-T、TaCKXD1a、TaGS-D1a利用率小于15%,有待提高。虽然千粒重相关基因的报道较多,但从小麦生长过程分析,育种家会先对穗数和穗粒数进行选择,千粒重在收获后才能有效选择。育种需要综合考虑产量三要素,因此加强对穗数和穗粒数的遗传解析,才能利用分子标记对亩穗数、穗粒数和千粒重同步选择,更好地满足育种的需求。亩穗数和穗粒数基因调控复杂,研究进展缓慢。本研究仅检测了TaMoc-A1、TEF-7A2 个与穗粒数相关的标记,其优异等位基因Hap-H和Hap-7A-3利用率均较低。
小麦品质相关基因研究开始较早,主要围绕面粉白度、籽粒硬度和麦谷蛋白等开展。本研究中,除硬度基因和黄色素位点Psy-D1优异等位基因占比较高外,TaPds-B1b、Zds-A1a优异等位基因利用均较少。此外,脂肪氧化酶(lipoxygenase,LOX)活性与种子耐储藏性有关,低LOX 活性可有效减轻籽粒的氧化变质,从而延长其储藏期[29]。PPO 活性与抗穗发芽有关,PPO活性和Ppo-A1相对表达量均与发芽指数呈显著正相关[30]。随着人民生活水平的提高,对优质小麦提出了不同的要求。小麦黄色素中的叶黄素和玉米黄质具有保护视力、抗氧化等保健功效[31],因此利用分子标记辅助选育高黄色素小麦有市场应用前景。高分子量谷蛋白亚基是影响面包加工品质的关键因素,由Glu-A1、Glu-B1和Glu-D13个位点控制,本研究成功检测了Glu-A1和Glu-D12 个位点,而Glu-B1最复杂,有超过20 种等位基因且序列相似性较高[32],导致分子标记开发困难,在分子育种中缺乏准确、实用的KASP 标记。低分子量谷蛋白亚基等位基因更多,也存在基因序列间相似性高、标记开发困难的问题,需要进一步研究。
倒伏、病害、干旱等因素都会导致小麦减产,保障高产的前提是提高小麦抗性。本研究中与抗病相关的优异等位基因的占比均较低,反映出已知基因在育种中利用不足,有必要加强对抗病基因Lr46+、Lr68+、Sbwm1-A、Pm21+、Pch1+、Lr34+、Yr15+、Fhb1+的利用。与抗旱相关的2 个标记优异等位基因占比较高,可能是因其与产量相关联而被间接选择。COMT-3B是为数不多与抗倒相关的功能基因,高茎秆木质素含量优异等位基因COMT3Ba有利于提高小麦抗倒伏性[33]。虽然单个基因的作用有限,但是随着更多相关基因功能标记的开发,分子标记辅助选择将在小麦抗倒育种中发挥越来越重要的作用。本研究共检测了3 个与抗穗发芽相关位点,Vp1B1和TaSdr-B1抗穗发芽Vp-1Bc和Sdr-B1a优异等位基因占比均较低;TaPHS1位点检测了2 个标记,一个标记优异等位基因占比较高,而另一个标记占比很低。Liu等[34]最新研究发现TaPHS1共有4 个单核苷酸多态性(single nucleotide polymorphism,SNP),综合4个SNP才能获得抗性最好的等位基因,即在本研究基础上,增加另外2 个SNP 的分子标记检测才能对TaPHS1进行更有效的辅助选择。
开花相关的光周期和春化基因决定了小麦品种(系)的生态适应性,因此在一定生态区域内大部分品种(系)的等位基因是一致的,只有个别位点存在差异。本研究中存在差异的位点为Ppd-B1和Vrn-D1a,仅少数品系为Ppd-B1 RTIndel Truncated copy和Vrn-D1a。而与早花相关的基因多为微效基因,对开花期有一定的调节作用,是相同生态区早熟类型的关键基因。本研究中除TaELF3-B1外,其他早花等位基因的比例均超过了85%,在早熟育种方面,可考虑利用优异等位基因TaELF3-B1 Wild type。
3.2 对区试品系进行分子检测的意义
区试品系代表了区域内的育种水平,通过对区试品系的研究分析可以了解该区域的育种进展。济麦22 是山东省小麦区试高产组对照品种,分子检测表明济麦22 是非1B/1R 易位系,矮秆基因是Rht-D1b,含有与产量相关的优异等位基因:TaCwi-A1a、TaCwi-4A Hap-4A-C、TaGW2-6B Hap-1、TaSus1-7A 1185-C、TaSus1-7B Hap-T、TaGASR7-A1 H1c;与品质相关的优异等位基因:Glu-A3g、Psy-D1a、Pinb-D1b、Pinb-B2b;与抗性相关的优异等位基因:Lr14+、1fehw3 RioBlanceo type、Dreb-B1a、TaPHS1 RioBlanceo type;与开花相关的优异等位基因:PRR-A1 Hap-I、PRR-B1 Hap-II、TaFT3-B1 G Allele、TaMOT1-D1 Wild-type;光周期和春化基因均为优势基因型(优异等位基因)。在高产组中超过80%的品系与济麦22 具有相似的等位基因组成,而差异主要在于TaGW2-6A、TaSus1-7A、Glu-A1、Glu-D1、Lox-B1、Lr14、Vp1B17 个基因。这7 个基因的优异等位变异占比不足50%,有必要在高产育种中提高利用率。济南17 是山东省小麦区试强筋组对照品种,除与济麦22 的绝大部分优异等位基因相同外,还含有TaGW2-6A Hap-A、TEF-7A Hap-7A-3、GluA1 Ax1/Ax2*、Pds-B1b、Lox-B1a、Lr46+优异等位基因。在强筋组中大部分基因的等位基因分布与高产组相同,区别在于强筋组均不携带1B/1R 易位系,TaGW2-6A、Glu-A1、Glu-D1、Lox-B1、Vp1B1的优异等位基因占比达到75%以上,表明这些基因在优质育种中已被育种家选择;而TaGS5、TEF-7A、Glu-A3g、Pds-B1、PRR-A1、PRR-B1、Lr46优异等位基因占比在50%左右,在品质育种中需要加强选择。虽然只有少部分参试品系能通过审定成为品种,但明确参试品系重要性状功能基因的组成对未审定品系的改良和审定品种的育种利用均有重要意义。明确区试品系已知基因的等位基因,不仅可以指导实践,用于亲本组配,还可以在后代选择过程中按照基因互补原则取长补短,将更多的优异等位基因聚合在一起,从而提高育种效率。
3.3 KASP标记应用
KASP 标记与需要通过凝胶电泳检测的分子标记相比,具有快速、简便、高通量等优势[11],目前已得到广泛使用。但已开发的部分KASP 标记可能存在问题,在进行基因分型时需要谨慎。如张维军等[35]对粒重相关的4 个KASP 标记进行了检测,其中TaGW2-6B的分型图中Hap-1等位基因与杂合基因型分群界限不明显,容易产生误判;TaGASR的分型图仅有H1c等位基因和杂合基因型,从品种品系多为纯合考虑,杂合基因型是否应为另一种等位基因需要进一步验证。本研究选用的KASP 标记先通过23 个DNA 样品测试,筛选分型效果好的标记再进行全部样品检测,但仍有部分标记存在数据点分散,杂合基因型较多和扩增失败的问题。除受DNA 质量和浓度影响外,主要原因可能是引物结合位点特异性不足,导致非特异扩增,从而影响分型效果。从多效抗病基因KASP 标记改良[13]的结果来看,可以通过重新设计引物来改良已开发标记,从而达到更好的分型效果。
4 结论
本研究对126份山东省小麦区试品系进行了64个KASP 标记的检测,结果表明与株高相关的Rht-D1,与千粒重相关的TaCwi-A1-1、TaGW2-6B、TaSus1-7B、TaGASR7-A1,与低分子量谷蛋白相关的Glu-A3g,与黄色素相关的Psy-D1,与硬度相关的Pinb-D1、Pinb2-V,与抗旱相关的1-feh-w3、TaDreb-B1,与抗穗发芽相关的PHS1-prom-222,与光周期相关的Ppd-A1、Ppd-B1、Ppd-D1,与春化相关的Vrn-A1、Vrn-B1、Vrn-D1,和与早花相关的TaFT3-B1、TaMOT1-D1、PRR-A1、PRR-B1优异等位基因利用率较高,其他基因的利用率较低,特别是与千粒重相关的TaGS-D1,与小穗数相关的TaMoc-7A,与抗赤霉病相关的Fhb1,与抗病相关的Lr34、Pch1、Yr15,与抗穗发芽相关的TaSdr-B1优异等位基因未检测到。
