强冻融作用下土壤微生物残体碳的累积特征及其对玉米秸秆输入的响应
2023-01-13曹鑫鑫刘丽君查丽霞卢溆阳徐英德
曹鑫鑫,刘丽君,查丽霞,卢溆阳,常 坤,徐英德*
(1 沈阳农业大学土地与环境学院,辽宁沈阳 110866;2 营口市农业农村综合发展服务中心,辽宁营口 115003)
土壤冻融作用是由于季节或者昼夜热量的变化,在表土及以下一定深度产生的反复冻结和解冻的过程,即“昼融夜冻”的现象,易发生在中、高纬度及高海拔地区,全球超过70%的陆地易受冻融作用的影响[1]。东北黑土区是我国粮食生产的压舱石,在保障我国粮食安全方面发挥着极为重要的作用[2]。该地区冬季时间长、气温低且降水量少,冬季土壤的冻结厚度较深(1.1~2.0 m)且持续时间很长(150天以上)[3]。而到春季后,昼夜温差较大,季节性冻融相当明显。冻融作用是影响土壤碳的生物地球化学循环过程的关键因素之一,探究冻融作用介导下的土壤有机碳周转过程,对深入理解东北黑土区土壤生态过程具有重要意义。
冻融作用主要通过影响土壤的物理、化学及生物学性质而对农田生态系统碳循环产生影响。它能够通过破坏土壤团聚体结构和微生物区系等途径促进土壤呼吸,进而直接影响到农田生态系统碳收支[4–5]。此外,土壤由冻结变为融化时,部分死亡的微生物会溶解并释放出细胞内所含有的有机质,增加了土壤可溶性碳含量,也会促进土壤有机碳的矿化[6–9]。但值得关注的是,微生物作为土壤有机碳循环的引擎,在参与有机碳分解的同时,自身也会以微生物残体的形式在土壤中累积,从而促进碳的固定并影响土壤碳库的平衡[10]。因此,冻融作用虽然会在一定程度上增加土壤微生物死亡的比例[11],但同时也可能对微生物源有机碳在土壤中的形成和累积产生遗留效应,这对于土壤碳库的长期截获具有重要作用。目前,微生物残体对土壤稳定有机碳的重要贡献已受到广泛关注,即随着微生物繁殖—生长—死亡的迭代过程,有机碳被微生物不断同化,并以微生物死亡残体和代谢产物的形式在土壤中持续累积,从而贡献于土壤有机碳固定[12]。越来越多的证据表明,微生物残体碳是土壤稳定碳库的主要组成部分[10, 12]。因此,深入探究冻融作用影响下的微生物残体碳累积特征及动态成为亟待解决的科学问题。此外,作物秸秆还田是农田土壤有机碳补充和更新的重要来源,也是实现联合国气候变化公约“千分之四”增碳计划的潜在方法[13–14]。作物秸秆能够为微生物活动提供养分和能源,从而调控微生物的群落结构及代谢过程[15],这势必会影响到微生物残体碳的产生和累积。然而,在冻融作用的干扰下,土壤不同类群微生物残体碳累积动态对作物秸秆输入的响应过程尚不明确。
本研究以典型农田黑土为研究对象,通过室内培养方法,采用微生物细胞壁来源的氨基糖作为微生物残体的标识物,研究在冻融作用的干扰下,土壤真菌和细菌残体碳的累积特征及其对玉米秸秆输入的响应,解析微生物残体碳对土壤有机碳的贡献,以期加深对冻融作用下土壤有机碳循环过程和微生物调控机理的认知,并为东北黑土区土壤肥力提升提供理论支撑。
1 材料与方法
1.1 供试材料
供试土壤采自黑龙江省海伦市(126°38´E,47°26´N)。该地区属于温带大陆性季风气候,年均温1.5℃,年均降水量550 mm。该地区土壤为发育于黄土母质的典型黑土,种植作物为玉米,产量水平在8.1 t/hm2。于冬季冻结前采集0—20 cm土壤样品,随机散点取样并混合后,将新鲜土样保存于低温条件下迅速带回实验室,用镊子剔除可见根系、石块等杂物,混匀后过4 mm筛,备用。供试土壤和玉米秸秆基本理化性质见表1。
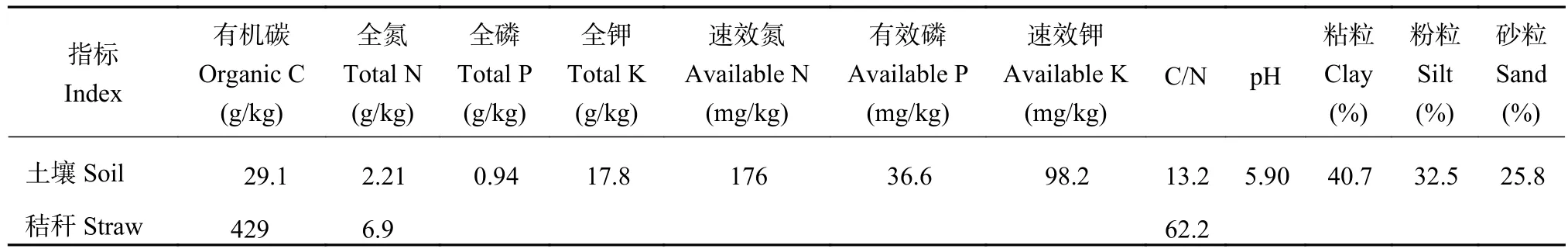
表1 供试土壤和玉米秸秆基础理化性质Table 1 Basic properties of test soil and corn straw
1.2 试验设计
采用室内模拟冻融培养法。以取样点近五年初春冻融时期的月平均高温(5℃)作为土壤融冻温度,三月下旬与四月上旬各半月平均低温作为土壤冻结温度(−4℃、−9℃),组成两个融冻温度/冻结温度组合处理:弱冻融(融冻温度/冻结温度为5℃/−4℃)、强冻融(融冻温度/冻结温度为5℃/−9℃),将没有冻结温度处理作为对照(5℃);各温度组合下分别设置添加玉米秸秆(S处理)和不添加玉米秸秆(CK)处理,共计6个处理。
具体室内培养方法如下:称取100 g风干土样,置于250 mL聚氯乙烯塑料瓶中,调节含水量至20%,用Parafilm封口膜封口并用针扎几个小孔,放入5℃恒温培养箱预培养30天。预培养结束后向设计为添加玉米秸秆处理的土壤中均匀掺入粉碎的玉米秸秆5.8 mg/g。由于玉米秸秆C/N较高,其腐解过程易受氮素限制,因此,以硫酸铵溶液调节其C/N值到25~30;同时,设计为不添加玉米秸秆的CK处理也加入等量硫酸铵溶液。用蒸馏水将每个塑料瓶中土壤含水量调节到28%后封口。先将进行冻融的土壤样品在5℃条件下培养24 h,随后逐渐降温,在−4℃或−9℃条件下冻结48 h,温度再逐渐上升,达到5℃后消融24 h,以此为一个冻融循环(4天)。不进行冻融的处理则始终置于5℃恒温培养箱中培养。培养过程中根据重量法定期调节土壤含水量。在试验开始后第0、3、8、12和16次冻融循环(即试验开始后第1、13、33、49和65天)进行土壤样品采集,将土壤样品风干后用于土壤有机碳及氨基糖含量测定。
1.3 测定方法
土壤有机碳和全氮含量通过元素分析仪(VARIO EL Ⅲ,德国)测定;土壤全磷含量采用HClO4–H2SO4消化,钼锑抗比色法测定;土壤全钾含量采用NaOH熔融法测定;土壤有效氮含量采用碱解扩散法测定;土壤有效磷含量采用Olsen法[16]测定;土壤速效钾含量采用乙酸铵提取—火焰光度计测定;土壤pH通过电极法(雷磁PHS-3B,上海)测定(土水比1∶2.5)。
土壤氨基糖含量采用Zhang等[17]提供的方法进行测定,简要步骤如下:称取含0.4 mg氮的风干土置于水解瓶中,加入 10 mL 6 mol/L HCl后水解 8 h。待冷却至室温后加入100 μL肌醇(内标1),振荡摇匀后过滤至心形瓶,在52℃真空状态下旋转蒸干。加入少量水将残余物溶解并转移至50 mL离心管,用KOH和稀HCl调节溶液pH至6.6~6.8。然后将离心管在3000 r/min条件下离心10 min,将上清液转移至5 mL衍生瓶并在45℃下用氮气吹干,干燥后溶解至 1 mL 水和 100 μL N-甲基氨基葡萄糖 (内标2),用Parafilm封口膜扎紧后冷冻干燥8 h。另取3个5 mL的衍生瓶作为标准样品,均加入100 μL胞壁酸,氮气吹干后加入100 μL氨基糖混合标准液[D-(+)-氨基葡萄糖、D-(+)-氨基半乳糖、D-(+)-甘露糖胺]、100 μL 肌醇、100 μL N-甲基氨基葡萄糖和1 mL水,封口后同样进行冷冻干燥。向干燥后的衍生瓶中加入300 μL衍生试剂,涡旋后置于80℃水浴锅中加热30 min,待冷却后,加入1 mL乙酸酐,再次涡旋后在80℃水浴锅中加热1 h,冷却后向各衍生瓶加入1.5 mL二氯甲烷,拧紧瓶盖涡旋30 s,用移液枪移出上层无机相。以相同方式,用蒸馏水(每次1 mL)进行有机相的提取,最后一次把水彻底去除。将衍生瓶剩余物质在45℃下用氮气吹干,用200 μL乙酸乙酯-正己烷混合液溶解干燥物并转入带有衬管的气相色谱瓶中。通过配有HP-5气相色谱毛细管柱 (30 m × 0.25 mm × 0.25 μm)的气相色谱仪(Agilent Technologies,美国)进行氨基糖检测。
1.4 数据处理方法
土壤中真菌残体碳 (fungal residue carbon,Fresidue-C,g/kg)和细菌残体碳 (bacterial residue carbon,Bresidue-C,g/kg)含量根据 van Groenigen 等[18]和 Liang 等[19]提供的公式进行估算:
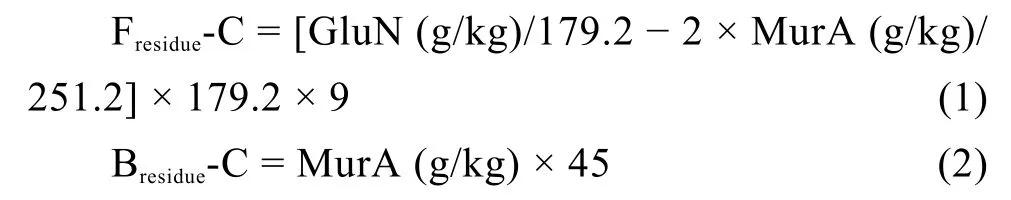
式中,179.2和251.2分别为氨基葡萄糖(GluN)和胞壁酸(MurA)的分子量;系数2假定细菌细胞中氨基葡萄糖和胞壁酸的摩尔比为2∶1[20]。转化系数9用来将氨基葡萄糖转化为真菌残体碳含量,转化系数45用来将胞壁酸转化为细菌残体碳含量[18–19]。真菌残体碳和细菌残体碳含量的总和视为总微生物残体碳含量 (total microbial residue carbon,TMresidue-C)[19]。
本研究中的数据以3个重复的平均值及其标准误差表示,试验数据采用SPSS 19.0进行方差分析。添加秸秆与不添加秸秆处理间的差异显著性通过配对样本t检验进行分析;不同冻融强度间的差异显著性用Duncan法进行多重比较,P<0.05为差异显著。图表的绘制采用Origin 2018软件。
2 结果与分析
2.1 土壤微生物残体碳的动态变化
冻融强度和玉米秸秆均对微生物残体碳含量产生明显影响(图1)。不添加秸秆条件下,强冻融处理的总微生物残体碳和真菌残体碳含量在第3~12次循环显著高于恒温培养处理 (P<0.05);而在试验结束(第16次循环)时,强冻融和弱冻融处理中总微生物残体碳和真菌残体碳含量却低于恒温培养处理,且以强冻融处理差异更为显著(P<0.05)。此外,在恒温培养和弱冻融条件下,在第3、8和12次循环总微生物残体碳和真菌残体碳含量均在添加秸秆处理中更高,但在第16次循环,添加秸秆均显著降低了总微生物残体碳和真菌残体碳含量(P<0.05);对于强冻融处理,添加秸秆在第3和12次循环均不同程度降低了总微生物残体碳和真菌残体碳含量,但在第16次循环,添加秸秆显著增加了总微生物残体碳含量(P<0.05)。同时,在添加秸秆条件下,除第8次冻融循环外,强冻融处理中土壤总微生物残体碳和真菌残体碳含量均低于恒温培养和弱冻融处理。

图1 不同冻融循环次数下土壤微生物残体碳含量的变化Fig. 1 Changes of soil microbial residue carbon contents under different freezing and thawing cycles
在不添加秸秆条件下,强冻融处理中细菌残体碳含量在第3和第8次冻融循环明显高于恒温培养(P<0.05)和弱冻融处理;而在第16次循环,不同冻融强度下细菌残体碳含量顺序为恒温培养(4.3 g/kg)>弱冻融(4.2 g/kg)>强冻融(3.6 g/kg)。此外,在恒温培养和弱冻融条件下,在培养前期(第3、8和12次循环)土壤细菌残体碳含量均在添加秸秆处理中更高,但在试验结束时,添加秸秆显著降低了细菌残体碳含量(P<0.05);在强冻融条件下,添加秸秆在第3次循环降低了细菌残体碳含量(P<0.05),但在试验结束时显著增加了细菌残体碳含量(P<0.05)。同时,在添加秸秆条件下,冻融强度总体上未对细菌残体碳含量产生显著影响(P>0.05)。
2.2 不同冻融循环次数对土壤真菌与细菌残体碳比值(F/B)的影响
由图2可知,土壤真菌残体碳含量平均是细菌残体碳的2.4倍。在不添加秸秆条件下,强冻融处理在第3次冻融循环F/B值最低(P<0.05);而在第12次和第16次冻融循环,恒温培养处理F/B值最低(P<0.05)。此外,在恒温培养和弱冻融条件下,添加秸秆在前期(第3次和第8次冻融循环)降低了F/B值,在后期(第12次和第16次冻融循环)则不同程度增加了F/B值;而在强冻融条件下,添加秸秆则在第12和16次冻融循环明显降低了F/B值(P<0.05)。同时,在添加秸秆的处理中,冻融强度并未在前8次循环中对F/B值产生明显影响;而在试验结束时,弱冻融和强冻融处理中F/B值分别比恒温处理降低7.5%和7.7%。

图2 不同冻融循环下土壤真菌与细菌残体碳比值(F/B)的变化Fig. 2 Dynamics of the residue carbon ratios of fungal to bacterial (F/B) under freezing and thawing cycles
2.3 微生物残体碳对土壤有机碳的贡献
在整个试验过程中,不同处理中总微生物残体碳(TMresidue-C/SOC)、真菌残体碳(Fresidue-C/SOC)和细菌残体碳(Bresidue-C/SOC)对土壤有机碳的贡献分别为44.7%~52.0%、30.8%~37.6%和12.6%~15.1%(图3)。冻融强度和玉米秸秆不同程度地改变了微生物残体碳的贡献率,且TMresidue-C/SOC与Fresidue-C/SOC变化趋势一致。在恒温培养和弱冻融条件下的前12次冻融循环,添加秸秆与不添加秸秆处理之间的TMresidue-C/SOC和Fresidue-C/SOC未表现出一致的规律,而在试验结束时,添加秸秆明显降低了TMresidue-C/SOC和Fresidue-C/SOC (P<0.05)。在强冻融条件下,添加秸秆处理在前期的TMresidue-C/SOC和Fresidue-C/SOC显著低于无秸秆处理,而第16个循环时不同秸秆处理间差异不明显(P>0.05)。同时,在不添加秸秆条件下,强冻融处理中TMresidue-C/SOC和Fresidue-C/SOC在第3次和第12次循环显著高于恒温培养和弱冻融处理(P<0.05),但在第16次循环,强冻融处理中TMresidue-C/SOC和Fresidue-C/SOC显著低于恒温培养和弱冻融处理(P<0.05)。在添加秸秆条件下,冻融强度对TMresidue-C/SOC和Fresidue-C/SOC的影响未表现出明显规律。
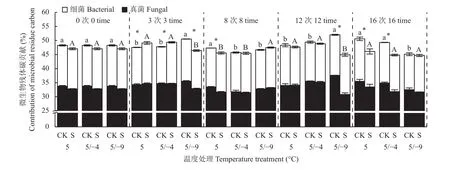
图3 不同冻融循环下微生物残体碳对土壤有机碳的贡献Fig. 3 Contribution of the microbial residue carbon to soil organic carbon under freezing and thawing cycles
此外,在第3和8次冻融循环,添加秸秆促进了恒温培养和弱冻融处理中的Bresidue-C/SOC,但在第12和16次循环,秸秆输入减少了恒温培养和弱冻融处理中的Bresidue-C/SOC;在强冻融处理中,添加秸秆在试验结束时对Bresidue-C/SOC影响不显著(P>0.05)。同时,在不添加秸秆条件下,强冻融处理中Bresidue-C/SOC在第3次和第12次冻融循环显著高于恒温培养和弱冻融处理(P<0.05),但在第16次冻融循环,强冻融处理中Bresidue-C/SOC显著低于恒温培养和弱冻融处理(P<0.05);在添加秸秆条件下,冻融强度对Bresidue-C/SOC的影响未表现出明显的规律,且在试验结束时不同冻融强度间差异不明显(P>0.05)。
3 讨论
3.1 不同微生物残体碳在土壤中的累积
有些研究表明,微生物残体碳能够在土壤中保持相对稳定[21–22],但本研究发现在冻融和添加玉米秸秆的情况下,土壤真菌和细菌残体碳含量均出现了较明显的波动(图1),证明微生物源氨基糖并不是完全惰性的,同样也会积极的参与到土壤碳周转过程中。尤其是存在外界干扰(如土壤环境的改变或人为措施的影响)的情况下,微生物残体碳动力学过程会发生明显变化。因此,微生物残体碳在土壤中的最终累积情况取决于其合成与分解过程之间的平衡。
在不添加秸秆的处理中,不同冻融强度对土壤微生物残体碳累积产生了差异化的影响(图1)。其中,在强冻融条件下,土壤真菌和细菌残体碳累积量在培养前期出现不同程度增加,说明冻融作用促进了土壤微生物繁殖—生长—死亡的迭代过程,微生物合成代谢占据优势地位,从而使微生物细胞组分逐渐以残体的形式在土壤中累积。有研究显示,冻融作用能使土壤团聚体破碎,从而促进可溶性有机碳和矿质养分的释放[6],这些均可作为土壤微生物生长的碳源和养分来源。虽然很多研究表明冻融作用在初期会促进CO2的释放[4–5],但本研究发现微生物在将更多的有机碳用于分解代谢的同时,也增加了合成代谢的效率,从而促进了微生物源有机碳在土壤中的累积。因此,需要辩证地看待微生物在土壤有机碳形成和损失之间的权衡作用,并深入探究二者之间的相对比例关系。此外,在试验结束时,冻融作用降低了土壤真菌和细菌残体碳的含量,尤其是强冻融作用降低效果更明显,说明冻融作用对微生物残体累积的优势作用并不会一直持续。该结果产生的原因可能是冻融作用所释放出的碳源和养分对微生物的促进作用,并不足以抵消土壤温度等环境因子对微生物活性产生的消极作用;同时,冻融作用对土壤结构的破坏会使部分赋存于团聚体中的微生物残体暴露出来,从而使它们更易于被微生物接触并降解。此外,本研究结果显示,强冻融处理对微生物残体碳含量的影响更大,证明冻融强度是调控土壤微生物源碳循环过程的重要因素。在较弱的冻融作用下,土壤及自身的微生物群落可以在一定程度上适应外界温度及水分条件的变化;而在昼夜温差更大的地区或季节,强冻融则会对微生物残体碳的累积产生较大影响,进而影响到整个土壤碳生物地球化学循环过程。
在恒温培养和弱冻融条件下,添加秸秆在试验前期促进了真菌和细菌残体碳的累积(图1);但在第16次循环时,添加秸秆显著降低了真菌和细菌残体碳含量。玉米秸秆在土壤中腐解的过程中,糖类和氨基酸等活性较高的含碳有机组分会优先释放出来,并成为微生物的可利用基质,刺激微生物的活性并促进微生物残体的产生[23–24]。但在本研究这种较低的温度环境条件下,玉米秸秆的腐解可能比常温(25℃)条件下更快进入缓慢分解阶段[25]。有研究显示,作物秸秆在腐解初期会促进微生物残体生成,但一段时间后会对原有微生物源碳产生明显的正激发效应;同时,在土壤中大量累积的植物源碳也会和微生物源碳竞争部分“固碳位点”[24]。此外,还有报道指出,微生物残体的不断积累会导致降解它们的微生物类群达到峰值[26],这些发现解释了本研究中玉米秸秆在试验结束时降低微生物残体碳的现象。对于强冻融处理,添加秸秆在试验前期不同程度降低了土壤真菌和细菌残体碳含量,而在试验结束时增加了细菌残体碳含量,该现象可能是由于冷冻胁迫和不同微生物对底物的竞争造成的微生物群落变化所引起的[27]。有研究显示,在高强度冻融条件下细菌会通过调整自身群落而比真菌更有利于竞争外源底物,从而保持生物量的相对稳定[27–28],这有利于维持细菌残体碳水平,而玉米秸秆的存在进一步促进了细菌残体碳含量的提升。该结果说明外源有机质的添加能够在一定程度上抵消冻融作用对细菌残体碳累积的不利影响。
3.2 真菌残体碳和细菌残体碳的相对比例及其对土壤有机碳的贡献
在本试验中,真菌残体碳含量平均为细菌残体碳的2.4倍(图2),且总微生物残体碳变化趋势与真菌残体碳一致,说明真菌残体碳是土壤微生物源碳库的主要贡献者。造成该现象的原因是真菌细胞聚合物相对稳定,而细菌细胞壁较薄,在土壤中更易被微生物分解,从而拥有更快的周转速率[24, 29]。因此,真菌残体碳对土壤稳定有机碳库的形成具有更重要的意义。
在本研究中,F/B值的变化暗示出土壤微生物群落结构和代谢功能的变化。不添加秸秆条件下,强冻融处理在第3次循环F/B值最低,而在第12次和第16次循环F/B值最高(图2),这说明强冻融处理虽然在试验前期和后期分别促进和降低了微生物残体碳含量(图1),但真菌和细菌残体碳变化的幅度并不一致。具体而言,强冻融处理在前期更有利于细菌残体碳累积,而在后期则加速了细菌残体碳的损失。短期的冻融作用有利于加速土壤中可利用养分的释放,导致C/N值较低的细菌群落增加更快;但较长时间的冻融作用后,细菌比真菌对低温的抗性更弱,细菌活性处于低能量下的代谢状态,养分同化效率显著下降,可能使细菌的比例降低更多[30]。有报道[31]指出,长期冻结会使土壤微生物量C/N值明显增大;同时,微生物群落结构逐渐向以真菌群落为主的方向转变,这支持了本研究的结果。此外,也不排除部分细菌残体被真菌分解,以满足其对碳源和自身生长的需求[32]。以上结果表明,长期高强度的冻融作用不利于细菌残体在土壤中的固定与累积。
值得注意的是,在添加秸秆后,冻融作用对F/B值的影响与不添加秸秆处理呈现出明显差异化的趋势:即冻融作用并未在前8次循环中对F/B值产生明显影响,而在试验结束时却显著降低了F/B值,且强冻融处理降低幅度更大(图2)。首先,添加秸秆会在前期缩小冻融作用对真菌和细菌残体碳相对比例的影响,这主要由于在前8次循环中,添加秸秆降低了恒温培养和弱冻融处理中的F/B值,从而缩小了其与强冻融处理的差异。该现象证明,在恒温和弱冻融条件下,细菌能够快速利用新输入的活性碳、氮进行自身生物合成,因而导致大量胞壁酸等细菌源残体在土壤中累积。其次,添加秸秆情况下,强冻融处理在试验结束时比恒温处理明显降低F/B值,这主要由于秸秆在后期增加了恒温培养处理中的F/B且降低了强冻融处理中的F/B值,暗示了秸秆添加在不同温度条件下对微生物群落不一致的调控模式。以上结果表明,添加秸秆在前期更有利于恒温培养和弱冻融条件下细菌残体碳的累积,而在后期加速了细菌残体碳的损失;在强冻融条件下,秸秆输入在后期促进了细菌残体碳的累积。
本研究显示,总微生物残体碳对土壤有机碳的贡献分布在 44.7%~52.0% (图3),与以往报道[19, 33–34]一致,再次证明微生物残体碳是土壤稳定有机碳库的重要组成部分。冻融作用和外源有机碳输入会进一步调控微生物残体碳对土壤有机碳的贡献。在不添加秸秆条件下,相较恒温培养处理,强冻融处理在前期增加了真菌残体碳(Fresidue-C/SOC)和细菌残体碳(Bresidue-C/SOC)对土壤有机碳的贡献,而在后期降低了Fresidue-C/SOC和Bresidue-C/SOC,这是真菌和细菌残体碳含量变化的结果(图1),也暗示着土壤有机碳组成和质量的变化,即较长时间高强度冻融作用降
低了土壤有机碳中微生物来源碳的比例。鉴于微生物残体碳在土壤稳定有机碳库形成中起到的重要作用[12],冻融作用所引发的微生物残体碳贡献的降低,可能会对土壤有机碳的长期固定带来进一步的不利影响。此外,添加秸秆条件下,不同强度的冻融处理在试验结束时Fresidue-C/SOC和Bresidue-C/SOC与恒温处理差异不大,进一步说明外源有机碳输入能够在一定程度上缓解冻融作用对真菌和细菌源有机碳所占比例的影响,这对于改善处于冻融作用地区土壤有机碳稳定性具有重要的意义。
4 结论
弱冻融处理(融冻温度/冻结温度为5℃/−4℃)对土壤微生物残体碳累积的影响较小,而强冻融(融冻温度/冻结温度为5℃/−9℃)不利于土壤微生物残体碳的长期累积,降低了其对土壤有机碳的贡献。细菌残体碳在长期强冻融作用下的分解程度大于真菌残体碳,因而长期冻融增加了真菌细菌残体碳比值。添加玉米秸秆对多次反复强冻融作用下真菌残体碳含量影响不显著,而明显提高了细菌残体碳的累积,在一定程度上缓解了强冻融对微生物残体碳累积的不利影响。因此,添加秸秆有利于增加强冻融作用下土壤微生物源有机碳的稳定性。
