点篮子鱼形态特征及6种组织乳酸脱氢酶的电泳分析
2023-01-10张涛周剑光甘金华何力
张涛,周剑光,甘金华,何力
(农业农村部水产品质量安全风险评估实验室(武汉),农业农村部淡水鱼类种质监督检验测试中心,中国水产科学研究院长江水产研究所,湖北 武汉 430223)
点篮子鱼(Siganusguttatus)隶属鲈形目(Perciformes)、篮子鱼科(Siganidae)、篮子鱼属(Siganus),主要分布在热带、亚热带的印度-太平洋及南海海域,从珊瑚礁到河口水域均有分布[1]。由于点篮子鱼体色艳丽、无肌间刺、营养价值高而备受人们青睐,是一种兼具观赏价值和较高经济价值的鱼类。同时点篮子鱼广盐、食性杂偏植食性、饵料成本低、生态幅度宽,使其成为一种优良的养殖品种引起人们的关注。目前在中国的海南、广东、江苏、浙江、山东、辽宁等地区均有养殖,已形成“南北共养”的局面[2-3]。另一方面,点篮子鱼喜食浒苔等藻类,与其他名贵养殖品种如刺参(Stichopusjaponicus)套养或混养,可有效控制养殖池中浒苔等藻类的过度生长,减少水质破坏,既有利于主养品种的生长,又有利于点篮子鱼的生长,属环境友好型鱼类[4]。目前有关点篮子鱼的报道主要集中在养殖繁育[1, 5-7]、生理学特性[8-11]、营养学特性[12-14]、发育生物学[15]以及环境生态学[16]等方面,关于点篮子鱼形态特征方面仅有少量报道[17-19],未见生化遗传特性方面的相关文献。研究形态特征可为探讨鱼类生理功能提供基础资料,同时也是种质资源鉴定最为简单直观的首选指标[18, 20]。同工酶作为一种生化遗传参数,广泛应用在鱼类的亲缘关系与分类、物种鉴定中的基因表达与调控研究以及群体遗传结构分析等方面[21]。本研究借助形态学描述观察、测定点篮子鱼可数和可量性状,并与已有研究结果比较,同时结合聚丙烯酰胺凝胶电泳技术分析不同组织中乳酸脱氢酶的表达情况,旨在进一步丰富点篮子鱼有关形态特征和生化遗传方面的研究内容,同时筛选出点篮子鱼种质的特征生化遗传参数,为制定其种质标准、促进点篮子鱼标准化生产和良种选育中的应用提供数据支撑。
1 材料与方法
1.1 实验材料
实验用点篮子鱼于2019年1月采自海南,共30尾,所有试验鱼均为天然海域捕获后经人工驯养所得,无伤残、无畸形,体重79.8~183.2 g,均值(134.68±28.06) g,体长14.2~27.4 cm,均值(16.52±2.31) cm。
1.2 实验方法
1.2.1 形态测定
按照《养殖鱼类种质检验 第3部分:性状测定》(GB/T 18654.3—2008)的规定,用游标卡尺(精确到0.01 mm)对30尾样本鱼进行测定。可数性状包括背鳍、胸鳍、腹鳍和臀鳍的鳍式、脊椎骨数以及左侧第一鳃弓外侧鳃耙数,共计6个参数。可量性状包括全长、体长、体高、体厚、头长、吻长、眼径、眼间距、尾柄长和尾柄高,共计10个参数。
1.2.2 组织酶液的制备、电泳及染色
组织酶液的制备、电泳及染色参照张涛等[22]的方法。
1.2.5 模式图的绘制
采用Bandscan 5.0电泳图谱中的酶带进行灰度识别,并根据识别灰度绘制电泳图谱模式图。
1.3 数据处理
采用SPSS 20.0(IBM公司,美国)对实验所得可量性状数据进行分析,结果以(平均值±标准差)表示。
2 结果与分析
2.1 形态描述及可数、可量性状
观测30尾点篮子鱼(图1)体呈椭圆形,侧扁,背高,体背缘和腹缘浅弧形,头较小,吻端至眼的背缘微凸;吻较短,口下位,口裂小,鳃盖膜与峡部相连;眼中等偏大,上侧位;侧线完全,位高,与背缘平行,向后延伸至尾鳍基部。背鳍长,分为两部分,前部分为棘组成,相连的后部分为分枝鳍条,起点位于腹鳍起点之前。胸鳍较大,后缘斜圆稍尖。腹鳍腹位,末端不达肛门。臀鳍长,后缘近圆弧形。尾柄短,尾鳍不分叉,外缘浅凹形。鳍式分别为:背鳍D. Ⅻ~XIV-9~11,胸鳍P. ⅱ-14~15,腹鳍V. Ⅰ-3-Ⅰ,臀鳍A. Ⅶ~Ⅷ-8~10。身体两侧上半部褐色,分布着许多橙黄色斑块,下半部灰褐色,背鳍、胸鳍和腹鳍浅黄色,臀鳍和尾鳍浅灰褐色。

图1 点篮子鱼外观形态Fig.1 Morphological measurement of Siganus guttatus
鳔1室,椭圆形。下咽齿3行。腹膜为灰黑色。第一鳃弓外侧鳃耙数为21~25。点篮子鱼可数、可量性状分别见表1,表中范围给出了所测各指标的上下限,均值反映了所测数据的集中程度。可数性状中侧线鳞数变动范围最大,背鳍条数不变。可量性状中主要以体长和头长为参照,给出了吻长、眼径和眼间距等头部主要参数与头长的比例关系,也反映了体高、尾柄长和尾柄高等躯干部主要参数与体长的比例关系。
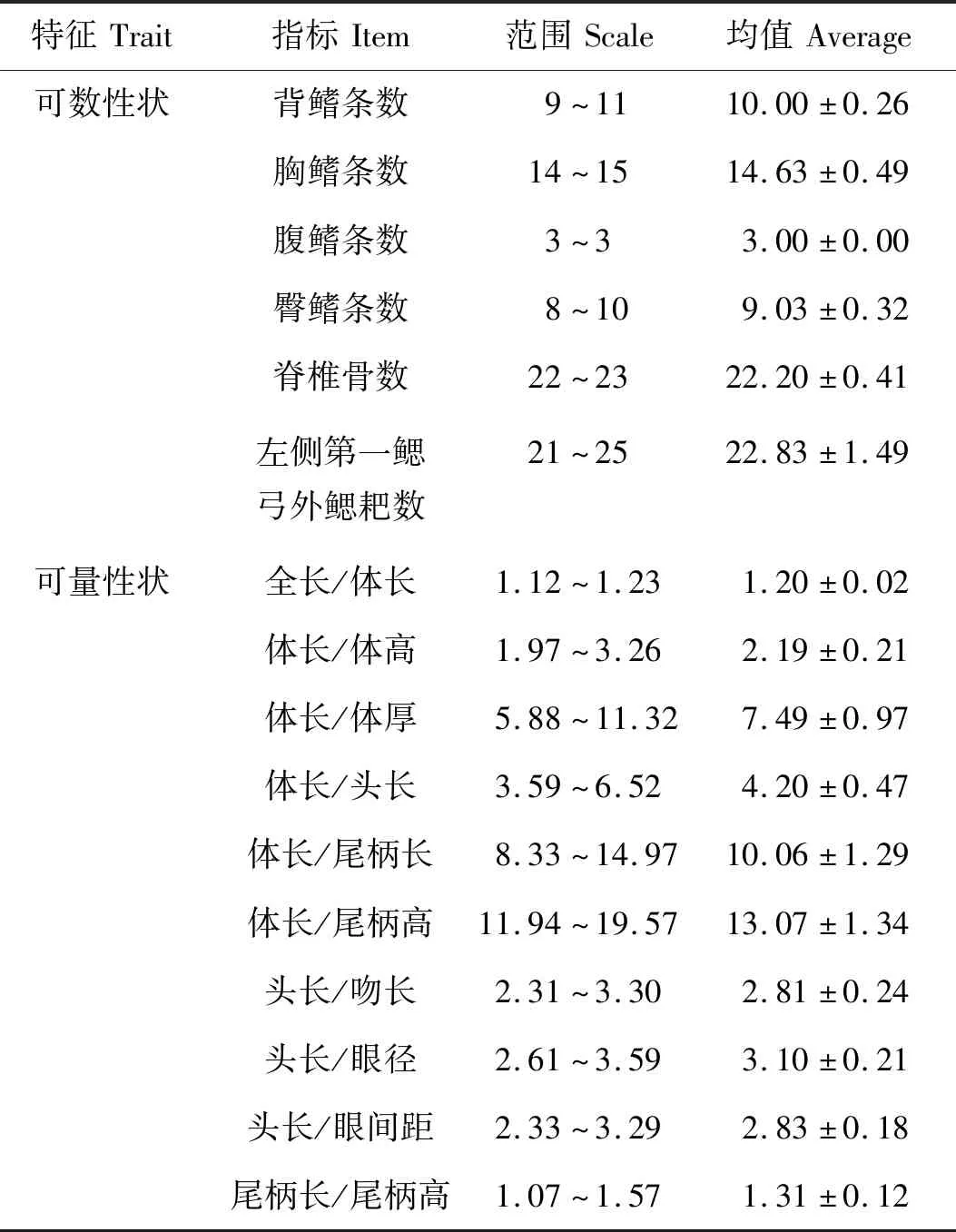
表1 点篮子鱼的可数性状和可量性状的均值与标准偏差Tab.1 The mean values and standard deviation of countable and measurable characters of Siganus guttatus n=30
2.2 乳酸脱氢酶同工酶的表达
点篮子鱼的LDH同工酶表达结果如图2所示,在点篮子鱼6种组织中共检测到7条LDH酶带,将靠近阳极最近的一条带定义为LDH1,自阳极向阴极方向依次编号为LDH2、LDH3…LDH7。不同组织中酶带数目和活性不同,呈现出明显的组织特异性。心脏组织中共检测到4条LDH酶带,其中LDH1和LDH2活性较弱,LDH4活性最强,LDH3居中;眼睛晶状体组织中共检测到7条LDH酶带,5尾样本鱼的LDH1和LDH2表达活性均较弱,LDH7表达活性最强,LDH6表达活性仅次于LDH7,5尾样本鱼的酶带数和表达活性程度相同;肌肉组织中共检测到3条LDH酶带,LDH1表达活性最强,也是5尾样本鱼的唯一共有酶带,LDH3仅在1号鱼中有检出,且表达活性较弱,LDH2在除1号鱼和2号鱼外的其余3尾样本鱼中有检出,且4号鱼中LDH2的表达活性程度较其余2尾鱼弱;肝脏中共检测到3条LDH酶带,LDH3为5尾样本鱼的共有酶带,且在2号鱼和3号鱼中表达活性程度较其余样本鱼强,LDH2仅在2号鱼和3号鱼中有检出,且表达活性程度较弱,LDH1在除4号鱼和5号鱼外的其余3尾鱼中有检出,且在1号鱼中的表达活性程度较其余2尾样本鱼弱;肾脏组织中共检测到4条LDH酶带,LDH1仅在1号鱼中有检出,表达活性程度较强,LDH2、LDH3和LDH4为5尾样本鱼的共有酶带,5尾样本鱼中LDH4的表达活性程度较其余2条共有酶带强,且2号鱼和3号鱼中LDH4表达活性程度较其余3尾鱼强,1号鱼中LDH2的表达活性程度较其余4尾样本鱼强;脾脏中共检测到3条LDH酶带,LDH3为5尾样本鱼的共有酶带,5号鱼中仅有LDH3检出,LDH3在2号鱼中的表达活性程度较其余4尾样本鱼强,LDH2的表达活性程度较其余2条酶带弱,2号鱼和3号鱼中LDH1和LDH2的表达活性程度均较1号鱼和4号鱼强。
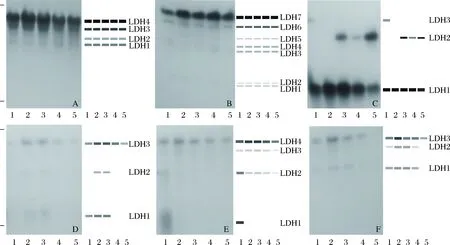
图2 点篮子鱼LDH电泳图谱A、B、C、D、E和F分别表示点篮子鱼心脏、眼睛晶状体、肌肉、肝脏、肾脏和脾脏的LDH酶谱;1~5泳道分别表示不同个体的酶谱。Fig.2 Electrophoretogram of LDH isozymes in Siganus guttatusA, B, C, D, E, and F show the electrophoretograms of LDH isozymes expressed in heart, eye, muscle, liver, kidney and spleen, respectively;1-5 show the zymograms in different individuals.
2.3 点篮子鱼特征生化遗传参数
本研究中5尾样本鱼肌肉、肝脏、肾脏和脾脏组织LDH酶带都有个体差异性,体现在不同样本鱼LDH酶带数不同,或者即使酶带数相同但不同样本鱼表达活性程度不同。虽然5尾样本鱼的心脏组织的LDH为单态即各样本鱼的LDH酶带数和表达活性程度都相同,但有拖带,分离效果并不是最佳,且制备心脏组织酶液时容易引入杂质,影响酶带分离,而所有样本鱼眼睛晶状体LDH酶带数和表达活性程度相同,各酶带分离效果相对较好而且稳定,所以初步确定以眼睛晶状体LDH作为从生化遗传特征层面鉴定点篮子鱼种质的备选对象。再选择30尾样本鱼的眼睛晶状体进一步电泳以验证眼睛晶状体LDH酶带表达情况,30尾样本鱼的眼睛晶状体LDH酶带表达情况一致,均为单态,8尾样本鱼眼睛晶状体LDH同工酶图谱(图3)表达稳定,分离效果好。鉴于此,本研究采用眼睛晶状体LDH作为鉴定点篮子鱼种质的特征生化遗传参数。

图3 点篮子鱼眼晶状体LDH电泳图谱1~8泳道分别表示不同个体的酶谱。Fig.3 Electrophoretogram of LDH isozymes expressed in eyes from Siganus guttatus1-8 show the zymograms in different individuals.
3 讨论
3.1 点篮子鱼的形态特征
形态特征是鱼类等动物最直观的种质表现之一[23]。鱼类身体的结构是其生理功能的基础,通过分析形态特征有助于了解、分析其捕食策略、摄食方式、运动行为和繁殖习性等生态特性。本研究中点篮子鱼背高、侧扁、椭圆形的外形与其作为一种生活在近海、性情较温顺的鱼类相适应,其游泳速度不及纺锤型鱼类如金枪鱼属鱼类(Thunnus)。大多数鱼类为1个背鳍,但鲈形目大多数鱼类背鳍分前后两部分,点篮子鱼背鳍分前后两部分也是其分类上属于鲈形目的体现之一,此外前部分带毒腺的棘也有利于其防御敌害。点篮子鱼中等偏大的眼睛与其作为近海鱼类,长期生存的水域光线相对充足有关,不同于生活在光照极弱或黑暗环境眼睛小甚至退化如穴居鱼类[24]。点篮子鱼口下位、口裂小便于其刮食珊瑚礁以及养殖网箱上附着藻类。形态特征是遗传和环境因素共同作用的产物[25]。
形态度量是研究生物表观形态性状的手段,可提供目标种类鉴别的关键形态指标,同时也为其系统分类和进化、种质鉴定和遗传育种目标性状筛选等研究提供技术支持[26]。本研究结果与文献中已报道点篮子鱼的形态特征进行比较,在可数性状方面如鳍棘数、不分支鳍条数和鳃耙数等不同于邹雄等[18]的研究,见表2。造成可数性状差异的原因除了与样本量大小有关外,还可能与计数者的主观判断有关,尤其对于小规格鱼类的计数、鳍棘的计数以及相邻不分支鳍条根部是否联会的判断,不同的研究者很容易出现研究结果不同,这一现象也在鲫属鱼类(Carassius)中出现过[27]。为避免一些埋于肌肉中的短小棘以及辨别数目不明显的分枝鳍条被少数或多数,本研究将样本鱼通过高温煮熟后在维持鱼体自身骨骼结构的前提下逐一计数鳍棘、不分支鳍条数,这种计数方法和邹雄等[18]的研究方法都成本低廉,但容易出现主观判断误差,以及人为剥离肌肉(无论煮熟与否)时的操作失误。X光片观察虽然相对昂贵,但直观、准确度高;愈豪祥[27]认为不同研究者在鲫属鱼类鳍棘数目计数上的判断误差源于研究者未采用X光片观察。笔者认为在研究鱼类可数性状时结合X光片观察能提高计数的准确度。

表2 点篮子鱼可数性状的比较Tab.2 Comparison of countable characters of Siganus guttatus
在可量性状方面,本研究体长/头长、头长/吻长和头长/眼径所得结果不同于邹雄等[18]和马强等[19]的研究结果,见表3,造成这一差异的原因除了与所测样本鱼的规格和样本量大小有关,还可能与测量方法有关。当然不排除养殖鱼类与野生鱼类在外部形态特征上的差异造成的。野生种群和养殖种群获取饵料难易程度不同,养殖种群饵料充分有保障,受敌害、环境胁迫相对少,有利于其生长,而野生种群饵料供应保障程度上不及养殖种群,而且相对养殖种群长期处于复杂多变的水生态环境,不利于其又好又快的生长[28]。

表3 点篮子鱼可量性状的比较Tab.3 Comparison of measurable characters of Siganus guttatus
3.2 点篮子鱼乳酸脱氢酶同工酶的表达
本研究发现LDH同工酶在点篮子鱼的各种组织中均有表达,说明LDH同工酶在点篮子鱼体各组织中广泛分布。从本研究结果来看,在点篮子鱼各组织中表达的LDH同工酶具有组织特异性,具体体现在LDH在同一个体的不同组织或器官中表达的酶带数和表达活性程度不同。同工酶是基因的生化反应表现,在电泳图谱上基因位点的表现形式以酶带呈现,由于基因位点的表达对组织具有选择性,致使某些基因位点仅在特定的某种或某几种组织中有表达,反应在不同组织电泳图谱上酶带数目就不同。本研究认为同工酶组织特异性表现在不同组织酶带数不同,与刘磊等[29]认为位点的表达体现同工酶的组织特异性相同。不同组织LDH表达活性程度不同,可能是由于同一基因在不同组织中的表达含量不同造成的。同工酶是基因表达的产物,其表达受到温度、压力、激素、氧容量和营养等内外因素的时空调控,致使其基因在各组织间的表达时间和强度不相一致,造成了不同组织同工酶酶谱的特异性[30]。
点篮子鱼LDH同工酶组织特异性与各组织的生理功能相关。LDH是由A、B、C3个基因位点编码的四聚体蛋白质。A、B基因在脊椎动物各组织器官中普遍表达,其表达产物依次对应电泳图谱上自阳极向阴极的5条酶带,即LDH1=B4、LDH2=AB3、LDH3=A2B2、LDH4=A3B和LDH5=A4,但也有物种只表达A4、B4两种和A4、A2B2和B4三种[31]。不同种同工酶代谢作用不同,其含量的差异能反应组织器官所处的生理状态。LDH1通过催化乳酸转变成丙酮酸,丙酮酸在线粒体中进行有氧氧化产生能量供机体所需,LDH1一般在有氧组织中表现出较高的活性。LDH5主要参与无氧酵解,催化丙酮酸转变成乳酸以产生能量。LDH5主要在无氧条件下起作用。本研究中点篮子鱼眼睛晶状体LDH7对应A4,在眼睛晶状体这种厌氧组织中参与无氧酵解。肌肉组织LDH1对应B4,其较高的含量说明点篮子鱼肌肉运动过程中消耗了大量的氧气。点篮子鱼心脏LDH4对应A4,心脏作为非厌氧器官而A4含量较高,反应了点篮子鱼处于低氧或无氧条件时心脏极力通过血液循坏为机体提供氧分和能量。C基因的表达具有高度的组织选择性,在绿色太阳鱼(Lepomiscyanellus)、鳟(Salmo playtcephalus)、黑鲈(Micropterussalmoides)等鱼类中,主要在神经视网膜光接收细胞和脑组织中表达,称为“眼带”[31]。本研究中点篮子鱼没出现经典的5条带,可能因为部分基因表达含量过低未检出或者未表达,对于眼睛晶状体组织中共有7条酶带,可能由于亚基的存在、某些位点产生了等位基因导致LDH酶带数目的增多。唐章生等[31]在研究巴西鲷(Prochilodusscrofa)时发现了类似的现象。本研究中点篮子鱼眼睛晶状体LDH1和LDH2可能是同一个位点的两个等位基因,LDH1和LDH2表现出阳极高迁移率,符合“眼带”的电泳特性,推测LDH1和LDH2是由C基因表达的产物。
4 结论
本研究采用传统形态学方法观察描述了点篮子鱼的形态特征,测定其可数、可量性状,并与已有研究结果进行比较分析。点篮子鱼形态结构与其生态特性密切相关,不同研究者关于可数、可量性状的测定结果不同,可能与样本规格差异、样本量、样本来源和测量方法有关。同时通过聚丙烯酰胺凝胶垂直板电泳较为系统的对点篮子鱼6种主要组织的LDH同工酶进行了分析,初步确定了眼睛晶状体组织中LDH表达丰富且活性稳定,可作为鉴定点篮子鱼种质的特征生化遗传参数。点篮子鱼LDH同工酶具有组织特异性,表现在LDH在同一个体的不同组织器官中表达的酶带数和表达活性程度不同,分析认为基因的表达以及表达含量在不同组织中具有选择性,导致LDH同工酶的组织特异性。这种特异性与组织执行不同的生理功能是相适应的。点篮子鱼眼睛晶状体组织中LDH1和LDH2可能是C基因表达的产物。
