草莓病害的生物防治研究进展
2023-01-05严聪文苏代发代庆忠张振荣田云霞董琼娥周文星陈杉艳童江云崔晓龙
严聪文 苏代发 代庆忠 张振荣 田云霞 董琼娥 周文星 陈杉艳 童江云 崔晓龙
(1.云南大学生命科学学院,昆明 650091;2.昆明市农业科学研究院,昆明 650034)
草莓(Fragariaspp.)为蔷薇科(Rosaceae)多年生常绿草本植物,在园艺学上属于浆果类果树[1]。草莓果实具有独特的风味且含有丰富的维生素、矿物质、不饱和脂肪酸、多酚类天然化合物以及丰富的抗氧化物质,如花青素和鞣花单宁等[2-3],因此受到了广大消费者喜爱,其种植面积也逐年增加。根据联合国粮食及农业组织(FAO)数据显示(http://www.fao.org/faostat/en/#home),截止2020年,中国草莓种植面积超过12.7万hm2,产量超333.6万t,产量居世界第一。目前,我国草莓种植主要以日光温室和塑料大棚为主,具有密闭、高温和高湿等特点,同时连作生产使得病原菌积累增多,导致草莓病害频发,造成大量损失[4]。草莓真菌性病害包括根腐病、炭疽病、灰霉病、白粉病、疫病、叶斑病等[5-9];细菌性病害如细菌性角斑病[10];病毒病包括草莓斑驳病毒、草莓轻型黄边病毒、草莓镶脉病毒、草莓皱缩病毒和草莓潜隐环斑病毒等[11]。草莓病害防治主要采用物理方法、化学方法和生物防治等方法,当前化学防治仍然为草莓病害防治的主要手段[12]。化学防治虽然能在短时间内起到较好的防治作用,但长期使用易诱发产生抗药性,导致防效下降,此外,农药残留还会危害环境和人体健康。生物防治具有对环境友好和病害防控过程中不易产生抗性等优点,是化学防治的重要补充手段。本文对草莓生物防治的微生物来源与相关机制以及未来可能的研究方向进行了综述,以期为草莓相关产业人员提供参考。
1 草莓病害生物防治的微生物来源
植物根际周围存在一类具有自生(游离或共生)固氮、溶磷、分泌抗生素或植物激素、拮抗病原菌和促进植物生长的植物益生菌,被称为植物促生根际细菌(plant growth promoting rhizobacteria,PGPR)[13-14]。丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)与世界上80%的陆生植物共生,可以通过增加根系对水分的吸收和利用效率来改善宿主植物的生长,通过形成菌根菌丝来增加根系的酶活性,扩大根系吸收养分的表面积,也是一类重要的植物有益菌[15-16],PGPR 和AMF 均具有生物防治的潜力,是生物防治微生物的主要来源[17-18]。此外,研究发现部分植物内生菌具有生物防治能力且与作物具有良好的适应性,受到研究人员关注[19-20]。目前的研究中,草莓生物防治微生物主要包括细菌的厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)和放线菌门(Actinobacteria),真菌的子囊菌门(Ascomycota)和毛霉门(Mucoromycota)(表1)。

表1 草莓生防菌及其防治病原菌Table 1 Biological control microorganisms for strawberry and their control pathogens
2 草莓生物防治机制与策略
在国内外研究人员的努力下,已揭示了应用于草莓病害防治的生防制剂的相关作用机制,包括抗生作用、竞争作用、真菌重寄生、促生作用和诱导抗性系统[14,55-56]。基于近年来微生物群落与草莓植物病害关系的相关研究[25,57-58],我们认为调节微生物群落结构(regulation of microbial community structure,RMCS)也是一类重要的生物防治策略。我们对生物防治相关机制及调节微生物群落作为生物防治策略进行综述,并绘制了草莓病害生物防治机制图(图1)。
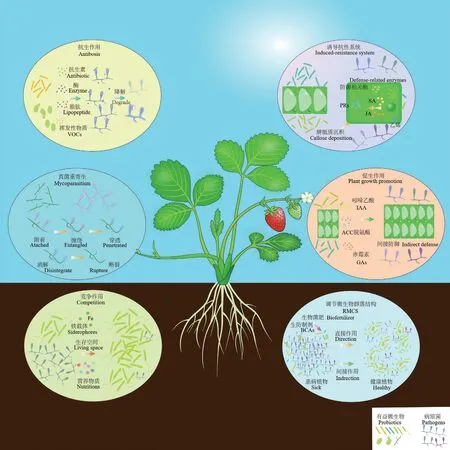
图1 草莓病害的生物防治机制与策略Fig.1 Biological control mechanisms and strategies of diseases to strawberries
2.1 抗生作用(antibiosis)
也叫做拮抗作用,指某些微生物能够分泌一些次生代谢产物或生物活性物质(如抗生素、抗菌蛋白等抗菌活性物质)来抑制其他病原微生物生长的现象[59]。这些代谢产物可通过破坏病原菌菌丝的细胞壁,导致菌丝畸形、断裂或细胞内液泡变大,从而抑制病原菌的生长和繁殖[32]。吸水链霉菌(Streptomyces hygroscopicus)B04 可产生几丁质酶、β-1,3-葡聚糖酶、纤维素酶和蛋白酶等细胞壁降解酶,抑制尖孢镰刀菌(Fusarium oxysporum)的生长[43]。几丁质酶能降解真菌细胞壁,被认为是一种可能替代化学杀菌剂的潜在生防制剂。草莓白粉病菌(Podosphaera aphanis)能被几丁质酶降解,有效抑制草莓白粉病的发生[60]。生防酵母如假丝酵母菌(Candida oleophila)、出芽短梗霉菌(Aureobasidium pullulans)和酿酒酵母(Saccharomyces cerevisiae)等[55],主要是通过产生挥发性物质(volatile organic compounds,VOCs)来抑制病原菌生长,这是一类脂溶性的易挥发的有机小分子化合物,包括烃类、醇类、硫醇类、酮类、硫酯类、环己烷类、杂环化合物、酚类和苯类衍生物[61]。其中乙酸乙酯和桂皮醛可能是主要的有效成分,能抑制灰葡萄孢(Botrytis cinerea)、果生链核盘菌(Monilinia fructicola)、互隔链格孢菌(Alternaria alternata)、炭黑曲霉(Aspergillus carbonarius)、枝孢霉属(Cladosporiumspp.)和炭疽菌属(Colletotrichumspp.)等草莓病害病原菌的菌丝生长,从而抑制草莓灰霉病、炭疽病、根腐病和叶斑病等病害[45,49-50,62-63]。尽管生防菌产生的多种代谢产物具有生防效果,但是生防菌产生的抗生素可能会诱导病原菌产生抗性或残留在农产品上对人体健康造成危害[64],因此,需要对这类生防菌进行环境及人体安全性评估。
2.2 竞争作用(competition)
草莓病害发生多数是由病原菌入侵引起的,通过气孔或机械损伤造成的伤口和产生附着孢等方式入侵植物体,利用植物体中的养分,快速繁殖而占据生存空间,引发病害[65]。生防菌与病原菌起初可能占据同样的生态位,但不同的生长速度、营养捕获效率或者攻击性方面的生理优势,可以使生防菌能更快争夺营养物质和生存空间,使病原菌无法定殖或繁殖,从而抑制病害发生[66]。棘孢木霉(Trichoderma asperellum)能迅速占领营养空间,对草莓炭疽菌(Colletotrichum fragariae)具有较强的竞争能力[46]。深绿木霉(T.atroviride)可以通过增强或者降低孢子萌发前的呼吸作用,与病原菌竞争生存空间[67]。绿色木霉(T.viride)在草莓叶片中与灰葡萄孢竞争营养,抑制灰葡萄孢菌丝生长和孢子产生[68]。植物乳杆菌(Lactobacillus plantarum)是一种草莓内生菌,其在草莓组织上定殖速度快于灰葡萄孢,能更快获得生存空间并抑制灰葡萄孢的孢子萌发,从而对草莓采后灰霉病具有防治效果[34]。低毒的病原菌也具有潜在的生防能力,如低毒的灰葡萄孢能通过产生大量菌丝和抗菌挥发性物质,来与强毒力的灰葡萄孢和油菜菌核病菌(Sclerotinia sclerotiorum)竞争,从而有效降低草莓果腐病的病情[69]。生防菌产生的铁载体(在低铁胁迫下生长的细菌和真菌所形成的相对低分子量500-1 500 Da 的铁离子特异性螯合剂[70])不仅可以为植物提供铁元素,还可以与病原菌竞争铜、锰、锌等元素,尤其是铁元素,使得病原菌不能正常生长,降低其致病性,从而达到防治病害的作用[71-72]。Henry 等[40]发现恶臭假单胞菌(Pseudomonas putida)KT2440 对草莓细菌性角斑病病原菌草莓黄单胞菌(Xanthomonas fragariae)具有抑制作用,缺乏铁载体生物合成途径的KT2440 菌株对其的抑制作用消失,重新加入假单胞菌铁载体后又能恢复其抑制作用,说明铁载体在与病原菌竞争的过程中起十分重要的作用。
2.3 真菌重寄生(mycoparasitism)
生防真菌直接和病原真菌接触,定殖并寄生于病原真菌上,并利用其营养,最终杀死病原真菌并占据其空间位置[73]。生防真菌菌丝沿寄主菌丝生长或缠绕寄主菌丝,产生的分枝菌丝穿透并嵌入病原菌菌丝内,导致病原菌丝细胞壁降解、内容物减少消解,菌丝体断裂、孢子瓦解或死亡,产生溶菌现象[46]。溶菌现象涉及多种酶作用,包括葡聚糖、几丁质酶、蛋白酶和脂酶[73-74]。宿白粉菌(Ampelomyces quisqualis)定殖和破坏白粉病菌菌丝的分生孢子梗和未成熟的子囊果,被宿白粉菌寄生的菌丝虽然还能生长,但不再产生无性和有性孢子[75]。哈茨木霉(T.harzianum)可以侵入菌丝,寄生于灰葡萄孢菌丝上[76]。Pseudozyma aphidis能寄生于白粉病菌和灰葡萄孢菌丝上,并与其竞争营养,另外,该菌分泌的化合物能使病原菌菌丝卷曲,液泡化和分枝等形态变化,从而防治白粉病和灰霉病[77-78]。尽管生防菌可以通过重寄生抑制草莓病害发生,但具有重寄生机制的生防菌生长至无病原菌区域可能造成外观品质下降等不利影响,导致这类生防菌应用范围有限的问题[66]。
2.4 促生作用(plant growth promotion)
PGPR 和AMF 等植物附生或内生有益菌可以通过固氮、溶磷、协助吸收铁等方式协助植物获取营养物质,产生维生素和调节植物激素水平来促进植物生长和增强防御反应,通过增加植物自身的抗病能力,间接抵抗病原菌侵染[54,58,79-81]。部分细菌能产生调节植物乙烯水平的1-氨基环丙烷-1 羧酸(1-aminocyclopropane-1-carboxylate,ACC)脱氨酶,ACC 脱氨酶作用于乙烯前体ACC,可降低植物体中的乙烯水平,与植物生长素如吲哚乙酸(indole-3-acetic acid,IAA)协同作用可以促进植物生长,提高植物的抗逆性,还可以抵抗病原菌危害[82]。嗜麦芽寡养单胞菌(Stenotrophomonas maltophilia)是从草莓叶中分离得到的一种内生菌,可产生铁载体、IAA 和赤霉素(gibberellins,GAs)以促进植物生长,并能防治病原菌C.nymphaeae造成的病害[36]。用解淀粉芽孢杆菌(Bacillus amyloliquefacien)、枯草芽孢杆菌(B.subtilis)和粉红黏帚菌(Clonostachys rosea)预处理草莓根系,具有明显的促生增产作用,可防治草莓灰霉病、炭疽病、白粉病和柱隔孢属叶斑病[24,26,33]。棘孢木霉对草莓幼苗具有明显的促生作用,草莓的根长、株高、植株总鲜重和总干重均明显增加[46]。尽管PGPR 和AMF 等植物有益菌能促生长并间接防治植物病害,但是在促进草莓生长的相关因子和草莓病害防治之间联系机制方面的文献报道并不多,后续研究可以进一步分析有益微生物产生的植物激素等在促生长和诱导抗病之间关联性的机制。
2.5 诱导抗性系统(induced-resistance system)
是由特定的环境刺激引起的一种“防御能力增强”的生理状态,通过这种状态增强植物的先天防御能力以应对后续的生物胁迫[83]。这种抗性增强的状态能有效抵抗多种病原体与寄生物,包括真菌、细菌、病毒、线虫、寄生植物甚至是植食性昆虫[84]。诱导抗性系统主要包括两种形式:系统获得性抗性(systemic acquired resistance,SAR)和诱导系统抗性(induced systemic resistance,ISR)。当植物暴露于有毒、无毒和非致病性微生物时可诱导植物触发SAR[85]。根据植物和诱导子的不同,SAR 建立所需的时间与病程相关蛋白(pathogenesis-related proteins,PRs)和水杨酸(salicylic acid,SA)的积累时间一致[86-88];ISR 则是通过PGPR 等非致病性根际细菌诱导的[83],与SAR 不同的是,ISR 并不依赖于PRs 或者SA 的累积,而是依赖于茉莉酸(jasmonic acid,JA)和乙烯(ethylene)的调控[89-90]。诱导抗性系统被证明具有3 条普遍性通路,其中两条与PRs 有关,一条与JA 和乙烯相关[91]。植物病原菌攻击和因病原菌攻击产生的创口或坏死都会诱导植物产生PRs,不同的是病原菌攻击是通过SA 信号通路诱导产生PRs,而创口或坏死植物体则是通过JA 信号通路产生PRs[91],这两种通路相互制约,可能是为了让植物根据病原菌来微调防御反应[89]。第三条通路则是由非致病性根际细菌诱导的依赖于植物激素JA 和乙烯的ISR 通路[14]。
诱导抗性系统与防御相关基因诱导表达、防御相关酶活性、活性氧的积累和植物抗毒素的合成等过程相关,限制病原菌生长与增殖,最终提高植物对疾病的抵抗能力[92-98]。贝莱斯芽孢杆菌(B.velezensis)XT1 不仅可以促进草莓细胞壁中胼胝质沉积,增强细胞壁对病原菌的抗性反应,还可以激活JA 和乙烯介导的防御通路[99]。在诱导防御反应过程中,由多种防御相关酶(defense-related enzymes)共同作用,包括过氧化物酶(peroxidase,POD)、过氧化氢酶(catalase,CAT)、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)、超氧化物歧化酶(superoxide dismutase,SOD)、多酚氧化酶(polyphenol oxidase,PPO)和苯丙氨酸解氨酶(phenylalanine ammonialyase,PAL)等[100]。POD、CAT 和APX 可以去除植物中的H2O2,SOD 是植物中重要的活性氧清除酶之一,POD 和PPO 可催化木质素的生物合成和酚类化合物氧化为剧毒的醌类,有助于形成抵御病原菌攻击的防御屏障,PAL 在苯丙类代谢中起着关键作用,参与酚类化合物和植物抗毒素的合成[101-103]。SA 不仅可参与调控草莓体内与防御相关酶的活性,如SOD、POD、CAT、PAL 和PPO,还可以调控某些次生代谢物的含量来增强对草莓白粉病的抗性,此外,SA 诱导初期可以与JA 协同作用激活多种防御基因的表达,使得草莓产生对白粉病的系统抗性[104]。接种葡萄汁有孢汉逊酵母(H.uvarum)后再接种灰葡萄孢,酵母产生的桂皮醛能诱导FAAPX、FAPPO和FaCAT等相关抗性基因表达,草莓果实中SA 信号通路相关基因PR1、PR5、WRKY1、GLU、NPR1和PAL表达量显著上调,果实中防御相关酶SOD、APX、CAT、PAL 活性上升,可降低草莓果实在冷藏期间的劣变速度,抑制草莓灰霉病的发生[49,52]。堆肥可以抑制草莓黄萎病菌大丽轮枝菌(Verticillium dahliae)菌丝生长和孢子萌发,还可以诱导草莓防御相关酶POD、PPO 和PAL 的活性增加,并降低丙二醛的积累,减轻草莓黄萎病的病症[105-106]。泡叶藻(Ascophyllum nodosum)提取物可以诱导POD、PPO、PAL 等酶的活性表达,抑制草莓白粉病[107]。哈茨木霉T22 接种草莓植株后,编码几丁质酶(FaChi2-1,FaChi2-2)和苯丙氨酸解氨酶(FaPAL)的基因表达显著升高,提高草莓对灰霉病的抗性[47]。草莓炭疽菌非致病性菌株可以通过积累活性氧(如过氧化氢和超氧阴离子)和沉积木质素与胼胝质来诱导草莓植株抗性,从而抑制草莓炭疽病[98]。总之,诱导抗性系统涉及多个通路,与抗性基因表达、病程相关蛋白和防御相关酶的调控密切相关,各个环节协同作用从而增强草莓对各种病害的抵抗能力。
2.6 调节微生物群落结构
随着宏基因组技术的发展,研究人员可对环境中的微生物组群落组成、结构和功能进行分析,越来越多的研究表明,微生物组与草莓病害的发生密切相关。如Huang 等[108]研究了连作10年的草莓大棚中的土壤微生物组,发现变形菌门细菌数量随着连作年限增加而逐渐减少,连作10年时木霉属和芽孢杆菌属丰度减少,而引起草莓枯萎病的尖孢镰刀菌丰度增加。杨俊誉等[109]比较研究了健康与发生白粉病草莓植株的根际原核微生物群落,结果表明,罹病草莓植株的土壤原核微生物的OTUs 数目和多样性均低于健康草莓植株。Mendes 等[110]研究发现变形菌门、厚壁菌门和放线菌门与抑制病害发生密切相关。Lazcano 等[57]研究发现,高抗病性草莓的根际微生物群落中对大丽轮枝菌和菜豆壳球孢菌(Macrophomina phaseolina)具有抑制作用的放线菌、酸杆菌和假单胞菌等在内的已知的生防菌丰度较高。基于前人的研究,我们提出调节微生物群落结构是一类重要的生物防治策略,即改变草莓相关微生物群落结构和微生态环境,增加有益菌丰度和减少病原菌数量,间接抑制草莓病害发生[23,111-115]。
微生物组对植物的生长和健康起着重要的作用,在改善植物生长、抗逆性和健康方面具有很高的潜力[56]。由链霉菌、芽孢杆菌和木霉等生防菌发酵而成的生物肥料,除对草莓植株具有抗病促生的效果外,还可促产保质[23,25,43,47]。草莓果面施用由解淀粉芽孢杆菌、哈茨木霉、球孢白僵菌(Beauveria bassiana)和Metschnikowia fructicol组成的生防菌后,可改变果面微生物群落组成和结构,抑制草莓采后病害发生[112,116]。使用蚯蚓和AMF 可以降低土壤中尖孢镰刀菌数量,同时增加土壤中放线菌的多样性,从而降低草莓枯萎病发病率[111]。经生物熏蒸后,土壤中放线菌门、假单胞菌属、芽孢杆菌属和毛壳菌属(Chaetomium)等生防菌的相对丰度增加,而土壤病原菌丰度减少,进而降低草莓病害的发生率[117]。Ye 等[118]分析发现捕食性粘细菌可通过捕食调节土壤微生物降低黄瓜枯萎病原菌尖孢镰刀菌的丰度,从而防控黄瓜枯萎病。Hu 等[113]研究表明,外源假单胞菌属细菌的加入可降低番茄根际中病原菌密度,进而降低了病害发生率。Li 等[119]研究了发生根腐病的黄芪(Astragalus mongholicus)根和根际的微生物群落组成后,通过筛选构建出了一种简化的人工微生物组,经抑制病原真菌生长、诱导植物产生系统抗性后,可防控黄芪根腐病。可见,调整微生物群落结构或人工构建微生物组(synthetic microbiome)在防治植物病害方面的潜力,然而,将其应用于草莓病害防治上的研究相对较少,这或许是未来的研究方向之一。
3 生物防治可能的研究方向
目前,制约生物防治发展的主要的原因包括田间应用时防治效果不稳定,缺少广谱的抗病活性,产品生产储存要求严格,或者对比化学制剂见效慢和效果不突出等原因[120]。生物防治涉及复杂的生态学动态过程,大部分有效的生物防治是在温室条件下取得的,而在针对大田作物病害时防治效果却很有限。同时,从环境中筛选的生物防治菌并不能直接作为生防产品使用,需根据其生理特点选择合适的载体、剂型和制备工艺等或开发为生物菌肥。因此,要开发具有高防效、高稳定性和高安全性的生防产品还需要大量应用基础研究。近年来,生防噬菌体和人工构建微生物组的研究也为生物防治提供了新的策略,值得深入研究。
3.1 噬菌体的生物防治研究
近年来,噬菌体被用作生防制剂定向防控食品中的各种细菌病原体,从熟食肉类到新鲜水果和蔬菜,同时,被批准用于食品的含有噬菌体的商业生防制剂产品也在增加[121]。噬菌体作为生防制剂能特异性作用于某种病原菌,可减少化学农药的使用,避免病原菌产生耐药性,降低成本和有利于维持田间生态平衡等优点[122-123]。利用噬菌体可以降低草莓果实上肠道病原菌的数量,降低人体因肠道病原菌而患病的风险,但也应当考虑噬菌体对人体有益肠道菌群的影响[124]。噬菌体蛋白也可以作为抗菌剂使用[125],如溶菌酶是一种通过降解细菌细胞壁肽聚糖以促使噬菌体释放的水解酶,对某些特定类型的肽聚糖具有特异性,从而可以特异性作用于某个属或某个种的菌[126]。然而,目前以噬菌体或其蛋白作为生防制剂应用于草莓的研究并不多,可能是因为草莓主要病害大多为真菌性病害,细菌性病害不突出,而真菌病毒的研究起步较晚且绝大部分真菌病毒的传播限于细胞内机制(特定类群存在体外传播和昆虫媒介传播),需要依靠菌丝融合水平传播,无性或有性孢子垂直传播,不同于噬菌体主要以裂殖方式快速传播[127]。目前真菌病毒的研究主要以分离鉴定和多样性研究为主,对其功能的研究较少[122]。另外,噬菌体作为生防制剂也面临着许多问题,如噬菌体无法自我复制繁殖、结构不稳、易突变以及缺乏对人体及环境安全性影响的相关评估[123],因此,噬菌体作为生防制剂任重道远。
3.2 人工微生物组的构建与应用
PGPR 与AMF 等植物相关微生物群具有促进植物生长和提高作物产量等功能,其中一些微生物已被分离出来,并作为生物肥料、生物刺激素和生防制剂等应用于农业生产中[25,106,128-131]。然而,PGPR 和AMF 等微生物在农业应用还有很大的局限性,为寻找能够在动态和应激环境中持续支持植物发育、获取营养、健康以及控制病害的微生物[56,132-134],越来越多的科学家开始对不同微生物菌株进行整合,依据相互作用、空间协调、菌群稳定和生物遏制等原则,人工构建微生物群落,这种群落被称为人工构建微生物组(或合成微生物群落)[135-136]。目前人工构建微生物组可应用于人体微生物组、植物微生物组和环境微生物组的改造,用于维持人体健康、提高植物的抗逆性及抗病能力和修复环境,还可应用于工业生产菌群的构建,解决代谢产物产率低以及毒素积累等问题[137-138]。Wei等[139]构建了与病原菌生态位相互重叠人工微生物组,降低了茄科雷尔氏菌(Ralstonia solanacearum)入侵番茄植株的成功率,抑制其生长,降低了番茄青枯病的发病率。Li 等[119]参照患病植物根部细菌群落特点构建了一组包含10 种高丰度和3 种低丰度细菌的抗病微生物组,根据在环境中的存活情况简化为含3 种高丰度和1 种低丰度细菌的微生物组,这两种人工微生物组对黄芪根腐病具有相似的防治效果,说明简化的人工构建微生物组具有应用潜力。在以往的生防菌的相关研究中,多从1-2 种菌株的有效成分、生防机制、防治病害以及对群落组成的影响等方面进行研究分析。若结合前人对生防菌性质、作用机制、相互作用和对群落影响的研究,结合PGPR 和AMF 等植物有益菌构建人工构建微生物组,应用于可持续农业,或可解决单一生防菌防效不稳定、有效时间短和作用范围小等问题。
4 总结与展望
草莓作为一种重要的经济作物,产生了巨大的经济价值,带动了相关产业人员就业致富,但是草莓的病害问题给草莓产业造成巨大损失。尽管草莓病害的生物防治研究取得了一定的成果,相关机制不断被揭示,但是也存在认识不足之处,应进一步研究草莓各种病害的致病机制与生防机制,探究环境因子与植物病害发生的关系,为实际生产中防控病害提供指导,同时研发能够用于实际生产的生防产品,深入研究人工微生物组的构建和应用,开发建立细菌性病害噬菌体疗法,减少或替代化学农药,避免污染环境和危害人体健康等问题。环境微生物群落结构复杂,相互作用方式多样,而微生物群落组成、结构和功能与植物生长、病害发生密切相关。草莓生物防治相关机制与应用的研究为人工构建微生物组提供了一定的理论基础。通过人工构建微生物组调节整个微生物群落结构与多样性,形成有效、稳定的促进植物生长和抗病的植株微生物组。未来,人工构建微生物组可能为防治农业病害提供更加有效、安全和稳定的策略和措施,提高作物产量和改善作物品质,促进农业的可持续发展。
