杉虎斑Galectin-8基因克隆及其在不同阿魏酸水平饲料下的表达响应
2023-01-05付伟杰邝杰华罗君黄建盛陈有铭陈刚
付伟杰 邝杰华 罗君 黄建盛 陈有铭 陈刚
(1.广东海洋大学水产学院,湛江 524088;2.广东蓝粮种业有限公司,湛江 524000)
半乳糖凝集素(galectins)属于糖结合蛋白家族中的一员,通过保守的糖识别结构域(carbohydrate binding domain,CRD)结合β-半乳糖苷类,不依赖Ca2+的活性,并具有多种生物活性功能,包括细胞迁移、凋亡、免疫反应、炎症细胞信号和神经保护等[1-2]。半乳糖凝集素在进化上是保守的,广泛存在于从原始到高级的各种生物中,并在不同的组织与细胞中表达[3-5]。半乳糖凝集素主要位于细胞内,也存在于细胞外,其缺乏一个N 端信号肽序列来引导它们通过内质网进行分泌,但可以通过非经典途径分泌到细胞外,从而具有细胞外功能[6]。此外,细胞内的半乳糖凝集素参与细胞功能过程,如前体mRNA 剪接、细胞生长调节和细胞周期过程[7-8]。半乳糖凝集素不但在微生物感染中发挥关键作用,也在先天性和适应性免疫反应中发挥潜在作用。迄今为止,已经在哺乳动物中鉴定出15 种半乳糖凝集素。根据结构特征、糖结合特异性和生物学功能,半乳糖凝集素可分为3 种类型:原型(Gale ctin-1、-2、-5、-7、-10、-11、-13、-14 及-15),含有一个CRD;串联重复类型(Galectin-4、-6、-8、-9 和-12),包含两个由连接肽连接不完全相同的CRD;嵌合体型(Galectin-3),由N 端的非凝集素结构域和C 端的CRD 所组成[9-10]。
Galectin-8 是一种串联重复型半乳糖凝集素,具有氨基酸序列相似性约35%的N 端糖识别结构域(N-CRD)和C 端糖识别结构域(C-CRD),并且两个结构域中均含有保守基序H-NPR 与WG-EE[11]。Galectin-8 在先天性和适应性免疫中具有重要作用。哺乳动物Galectin-8 可以通过监测核内体和溶酶体的完整性来检测入侵的细菌,然后激活抗菌自噬以保护细胞免受细菌的感染[12]。据报道,半滑舌鳎(Cynoglossus semilaevis)的Galectin-8 对某些革兰氏阴性菌病原体具有强大的杀菌活性,可直接破坏它们的膜和结构[13]。Galectin-8 可促进浆细胞的形成和调节其分化,并在促进抗体产生的过程中起到附加和冗余作用[14-15]。Galectin-8 通过串联重复结构与白细胞介素2 和转化生长因子-β 受体相互作用,促进T 细胞的增殖与分化[16]。此外,Galectin-8 被认为是一种早期的脑保护剂,不仅能抑制炎症信号,促进抗氧化活性,还能减少β 淀粉样蛋白寡聚化与神经原纤维缠结沉淀,增强对阿尔茨海默病的预防作用[2]。目前,关于Galectin-8在硬骨鱼类中的研究仅见于尼罗罗非鱼(Oreochromis niloticus)[17-18]、大弹涂鱼(Boleophthalmus pectinirostris)[19]、黑岩鱼(Sebastes schlegelii)[20]、牙鲆(Paralichthys olivaceus)[21]、鳜(Siniperca chuatsi)[22]和半滑舌鳎[13],缺乏关于石斑鱼Galectin-8的研究报道。
杉虎斑是由雌性褐点石斑鱼(Epinephelus fuscoguttatus)与雄性清水石斑鱼(E.polyphekadion)杂交的子一代,具有杂交优势[23]。石斑鱼具有营养丰富、肉质鲜美和经济价值高等特点,已在十几个沿海国家中广泛养殖[24]。据联合国粮食及农业组织(FAO)统计,2019年全球石斑鱼养殖总产值为820 794 707 美元,养殖总产量更是达234 828.45 t,其中中国的养殖产量最大,占总产量的78%,多达183 127 t[25]。然而,高密度的集约化养殖与不恰当的投喂会损害鱼类的免疫系统,使其易受病原体的感染[26]。研究报道,饲料中添加免疫刺激剂可以作为预防和治疗水生动物感染性疾病的有效措施。草本植物及其衍生物可作为一种廉价的抗生素替代物,可以提高鱼类的免疫和生长性能[27]。阿魏酸(ferulic acid,FA)属于天然的植物化学物质,是一种低毒酚酸,可在植物界中广泛获得。阿魏酸具有多种生理活性功能,如抗菌、抗炎、抗氧化、抗癌、抗病毒和保护肝脏等[28]。研究石斑鱼的免疫相关基因和增强免疫的方法对于获得高产量至关重要。因此,本研究运用RACE 技术首次克隆出杉虎斑Galectin-8的全长cDNA 序列,然后对其进行生物信息学分析,并通过实时定量PCR(RT-qPCR)分析Galectin-8mRNA 的组织特异性分布;此外,还研究了饲料中添加不同水平阿魏酸对Galectin-8基因表达的影响,以期为进一步探究鱼类免疫防御机制提供一定的依据,同时为控制水产养殖疾病暴发提供新的思路和策略。
1 材料与方法
1.1 材料
试验用杉虎斑幼鱼购自广东湛江东简镇石斑鱼苗场,在湛江恒兴南方海洋科技有限公司室内1m3玻璃钢桶中暂养驯化15 d,期间投喂石斑鱼商品配合饲料。配制7 组添加不同水平FA 的等氮等脂试验饲料,FA 水平分别为0(对照组)、40、80、160、320、640 和1 280 mg/kg,试验饲料组成及营养水平见表1。暂养结束后,将试验鱼饥饿24 h,称重后将初始体重为(9.55 ± 0.60)g 的试验鱼分组,挑选大小基本一致且健康的试验鱼,随机分配到21 个玻璃钢纤维桶中,养殖10周。试验共设计7个处理组,每处理3 个重复,每重复1个0.5 m3的玻璃钢纤维桶,每桶放30 尾鱼。每天表观饱食投喂2 次(8:00 和17:00),日投喂量约为石斑鱼体重的3%-5%。试验用水为经过沙滤和沉淀的天然海水,光照周期为自然光照,试验期间不间断充氧气,溶解氧量不低于6 mg/L,盐度为29%,温度范围为25-30℃,pH 7.6-8.2,且氨氮和亚盐含量不大于0.05 mg/L。
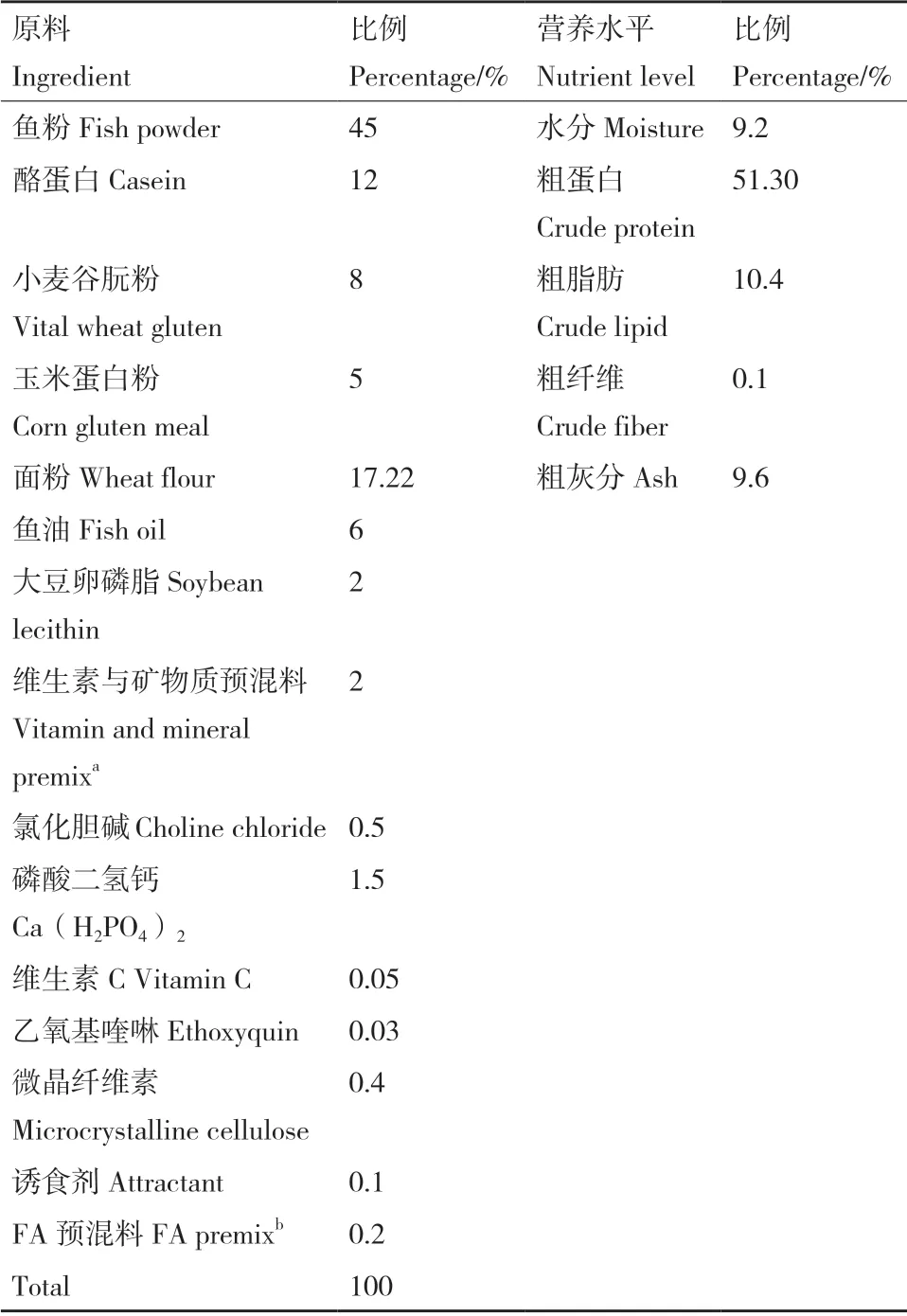
表1 试验饲料组成及营养水平Table 1 Ingredient composition and nutrient levels of the experimental diets
1.2 方法
1.2.1 样品采集 养殖试验结束后,将试验鱼禁食24 h,采样时使用丁香酚进行麻醉。首先在对照组中进行取样,每桶随机取3 尾鱼,解剖分离得到脑、胃、肝脏、鳃、肠道、中肾、头肾、心脏、脾脏、肌肉和皮肤。在其余的处理组中,每桶取3 尾鱼进行解剖获得肝脏、肠道、脾脏及头肾。各组织样品收集于装有RNA Later 的2 mL 无菌离心管中,液氮快速冷冻后置于-80℃超低温冰箱中用于RNA 的提取。
1.2.2 RNA 提取及第一链cDNA 的制备 按照说明书,使用TranZol Up 试剂(TransGen)提取各组织总RNA。采用超微量分光光度计(SimpliNano,Biochrom,UK),在260 nm 条件下检测RNA 的浓度,并用260 nm 与280 nm 的比值(范围在1.8-2.1)测定其的纯度。通过1.5%的琼脂糖凝胶进行电泳,检测提取的RNA 质量。通过反转录试剂盒EasyScript®One-Step gDNA Removal and cDNA Synthesis SuperMix(TransGen),在PCR 扩增仪中设置42℃ 15 min;85℃ 5 s,将1 μg RNA反转录成cDNA,并保存于-80℃用于之后的基因克隆和RT-qPCR 分析。
1.2.3Galectin-8基因全长cDNA 的克隆 从课题组的杉虎斑转录组数据中筛选出注释为Galectin-8的Unigene,并在NCBI 中进行BLAST 比对确定为鱼类的Galectin-8基因片段。使用Primer Premier 5.0 设计特异性引物Gal-8-F 和Gal-8-R 克隆中间片段,PCR 程序设置为:95℃ 5 min;95℃ 30 s,60℃ 30 s,72℃ 45 s,共循环35 次;72℃ 10 min。用1.5%的琼脂糖凝胶电泳检测PCR 产物,并使用EasyPure Quick Gel Extraction Kit(TransGen) 进行目的片段切胶回收,并连接到pMD-18T Vector(TaKaRa),转化到DH-5α 感受态细胞(TaKaRa)后,挑选出单克隆阳性菌落,随后送至上海生工生物工程公司进行测序。按照SMARTer®RACE 5′/3′ Kit(TaKaRa)的操作说明,并根据获得的中间片段序列,分别设计两个基因特异性引物(Gal-8-5′GSP 和Gal-8-3′GSP)和两个巢氏基因特异性引物(Gal-8-5′NGSP 和Gal-8-3′NGSP)。5′-RACE和3′-RACE cDNA 模板合成完后,进行cDNA 5′端和3′端的扩增,PCR 产物经纯化后,进行切胶回收,连接,转化与测序。根据重叠部分将测序得到的5′端和3′端序列与中间片段序列进行拼接,得到Galectin-8基因全长cDNA 序列。引物序列详见表2。
1.2.4 生物信息学分析 使用ORF finder 在线工具(https://www.ncbi.nlm.nih.gov/orffinder/)来识别开放阅读框序列(ORF)和预测氨基酸序列。应用Simple Modular Architecture Research Tool(SMART)(http://smart.embl-heidelberg.de)对Galectin-8 的结构域进行分析。信号肽和跨膜区的预测分别使用signalIP 5.0(https://services.healthtech.dtu.dk/service.php?SignalP-5.0) 和TMHMM 2.0(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)。利用BLAST 程序(http://blast.ncbi.nlm.nih.gov/Blast.cgi/)对cDNA 序列和氨基酸序列进行同源分析。通过DNAMAN 9.0 对氨基酸序列进行多序列比对分析,并用MEGA X 软件,采用邻接法(Neighbor-Joining)构建系统发育进化树,bootstrap 值设置为1 000。多序列比对与构建系统发育进化树所用的氨基酸序列见表3。采用SWISS-MODEL(http://www.swissmodel.expasy.org)程序进行蛋白质三级结构的构建,ProtParam tool(http://web.expasy.org/protparam/)预测蛋白质的理化性质(分子量、理论等电点和亲水性)。蛋白质二级结构和亚细胞定位的预测分别采用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html/) 和PSORT II Prediction(http://psort.hgc.jp/form2.html)。使用ExPASy 服务器的NetNGlyc 1.0 server(https://services.healthtech.dtu.dk/service.php?NetNGlyc-1.0) 和NetPhos 3.1server(https://services.healthtech.dtu.dk/service.php?NetPhos-3.1)分别进行N-糖基化位点和磷酸化位点的预测。
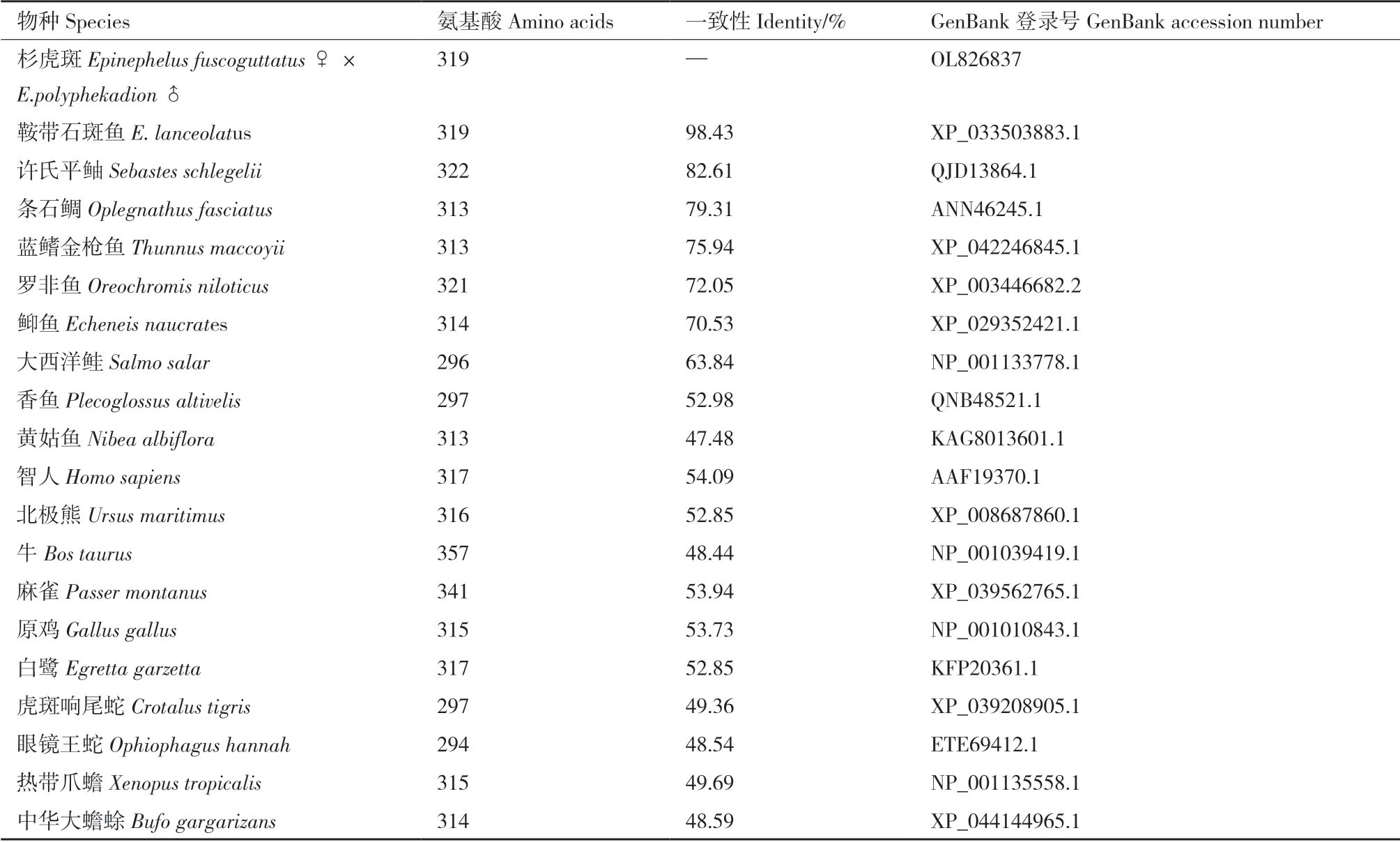
表3 杉虎斑Galectin-8 氨基酸序列多重比对与构建系统进化树所用到的氨基酸序列Table 3 Amino acid sequences used for multiple sequence alignment and phylogenetic tree construction for Galectin-8 from hybrid grouper
1.2.5 实时定量PCR 根据克隆获得的cDNA 序列,设计实时荧光定量引物Gal-8-RT-F 和Gal-8-RT-R(表2)。在LightCycler 96(Roche,Switzerland)仪器中进行RT-qPCR,使用的是PerfectStartTMGreen qPCR Super Mix(TransGen)试剂盒。10 μL 反应体系包括5 μL 的2×PerfectStartTMGreen qPCR Super Mix、0.5 μL 的正向引物、0.5 μL 的反向引物、0.5 μL 的cDNA和3.5 μL 的ddH2O(Double distilled water)。设置反应程序为94℃ 30 s(1 个循环);94℃ 5 s,60℃ 15 s,72℃ 10 s(40 个循环);95℃ 10 s,65℃ 60 s,95℃1 s(1 个循环);37℃ 30 s(1 个循环)。通过Light-Cycler® 96 Software 分析荧光定量数据,获得Cq 值。以β-actin为内参基因,用2-ΔΔCt方法计算基因的相对表达量。在统计分析之前,对数据进行了正态性和同质性检验,并使用SPSS 22.0 软件进行单因素方差分析(One-Way ANOVA),组间若有显著性差异,采用Duncan 氏法进行多重比较,P<0.05 表示差异显著。每个处理组均有3 个生物学重复。
2 结果
2.1 Galectin-8基因cDNA全长克隆及序列分析
杉虎斑Galectin-8基因的cDNA 全长为1 368 bp(GenBank 登录号:OL826837),其中5′非翻译区(5′untranslated region,5′UTR)为16 bp,3′UTR 为394 bp,ORF 长960 bp,共编码319 个氨基酸,还包括一个典型的加尾信号序列(AATAAA)和一个长为27 bp 的poly(A)尾巴(图1)。Galectin-8 蛋白分子质量为35.26 kD,理论等电点为7.65,总平均亲水性为-0.149(亲水),不稳定系数为40.82(不稳定)。TMHMM 2.0 和signalIP 5.0 预测结果显示,杉虎斑Galectin-8 蛋白无跨膜结构域与信号肽。亚细胞定位结果表明,Galectin-8 蛋白定位于细胞质、细胞核、线粒体、内质网和分泌小泡的概率分别为60.9%、21.7%、8.7%、4.3%和4.3%。NetNGlyc 1.0 server 和NetPhos 3.1server 预测结果表明,Galectin-8 蛋白含有1 个N-糖基化位点和29 个磷酸化位点(图1)。
SMART 预测结果显示,Galectin-8 蛋白含有两个CRD,分别位于N 末端(15-150,136 aa)和C末端(187-319,133 aa),属于串联重复型,由序列长为36 个氨基酸的连接肽(Linker peptide)连接。此外,在两个CRD 上均鉴定出两个与β-半乳糖苷类结合的保守基序,分别为 HXNPR 和WGXEE,其中X 表示任意氨基酸(图1)。利用SWISS-MODEL对杉虎斑Galectin-8 蛋白进行同源建模(图2),发现与人的Galectin-8 三级结构相似,序列相似性为56.94%,GMQE 为0.78。其中,α- 螺旋占2.51%,β-转角为8.46%,延伸链占33.86%,无规则卷曲为55.17%。

图1 杉虎斑Galectin-8 的全长cDNA 核苷酸序列和推导的氨基酸序列Fig.1 Full length cDNA nucleotide and deduced amino acid sequences of Galectin-8 in hybrid grouper
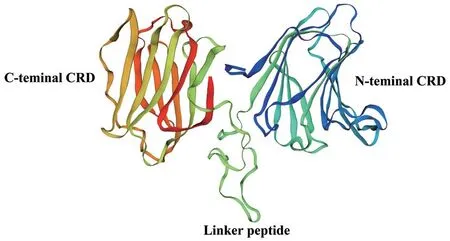
图2 同源建模预测的Galectin-8 蛋白三维结构Fig.2 Tertiary structure of Galectin-8 predicted by homology modeling
2.2 同源性及系统发育进化树分析
杉虎斑与其他物种Galectin-8 的氨基酸序列同源性分析结果如图3所示。通过NCBI 上blastp 比对,分别获得杉虎斑Galectin-8 推断的氨基酸序列与鞍带石斑鱼、许氏平鲉、蓝鳍金枪鱼、条石鲷、䲟鱼、大西洋鲑、智人、原鸡和热带爪蟾的一致性(表3),其中与鞍带石斑鱼的一致性最高,达到98.43%,许氏平鲉次之为82.61%,而与热带爪蟾的一致性最低为49.69%。同源性分析表明,Galectin-8 在不同动物中均具有保守性。参与氨基酸多序列比对的各物种Galectin-8 均具有串联重复半乳糖凝集素的典型结构特征,包括一个N-CRD 和一个C-CRD,并且两个CRD 均有两个保守基序H-NPR 和WG-EE。
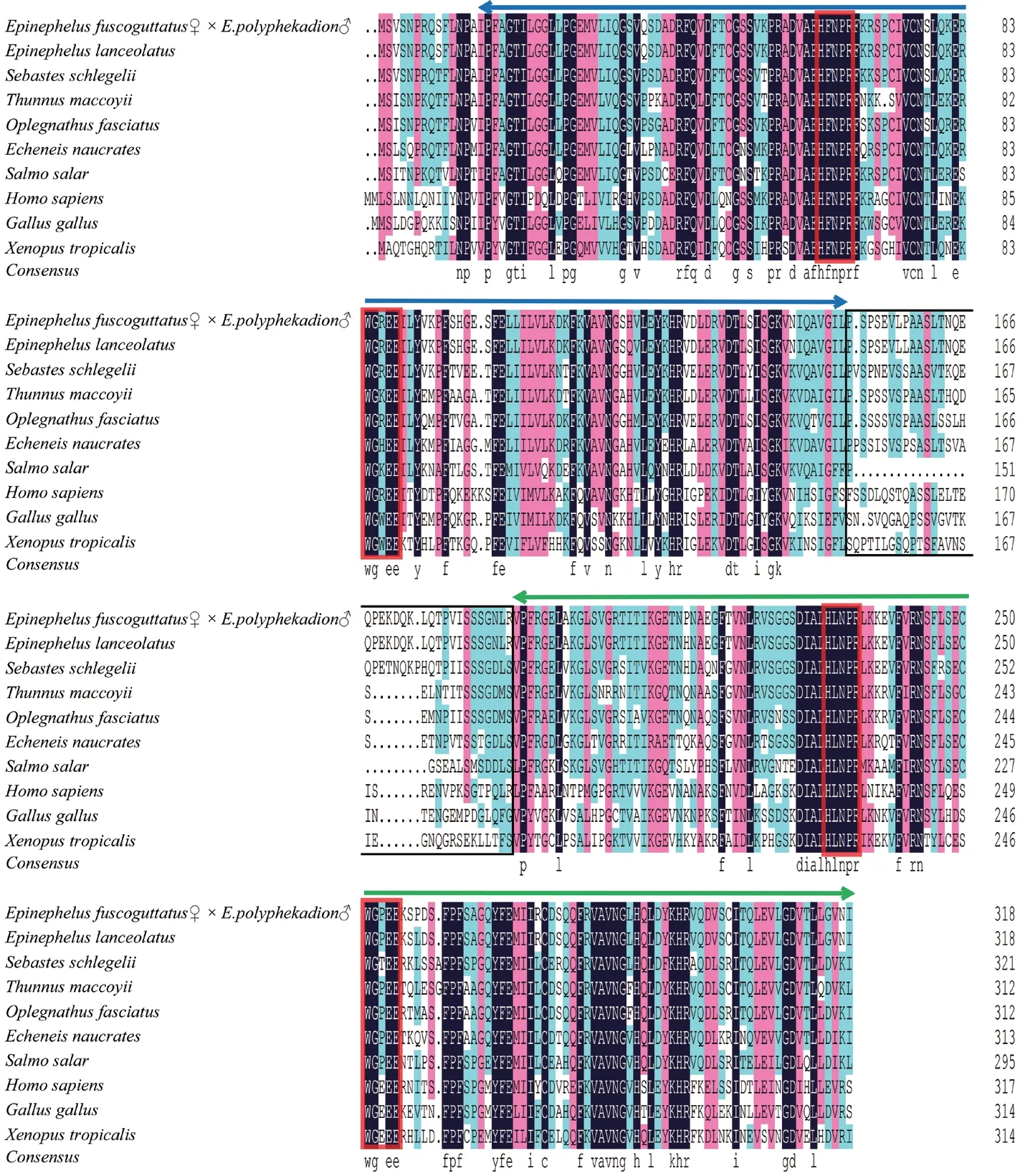
图3 杉虎斑与其他物种 Galectin-8 氨基酸序列的多重比对Fig.3 Multiple sequence alignment of Galectin-8 amino acid sequences between hybrid grouper and other species
为了研究杉虎斑Galectin-8 与其他脊椎动物的进化关系,采用邻接法构建系统发育进化树(图4),共包括20 个代表物种的氨基酸序列。结果表明,杉虎斑与其他硬骨鱼类的Galectin-8 单独聚为一支,爬行纲、两栖纲、哺乳纲和鸟纲动物的Galectin-8 各自聚为一支。杉虎斑Galectin-8 与鞍带石斑鱼具有最近的亲缘关系,许氏平鲉次之,而与黄姑鱼的亲缘关系相对较远,与爬行纲、两栖纲、哺乳纲和鸟纲其他动物的亲缘关系相对更远。
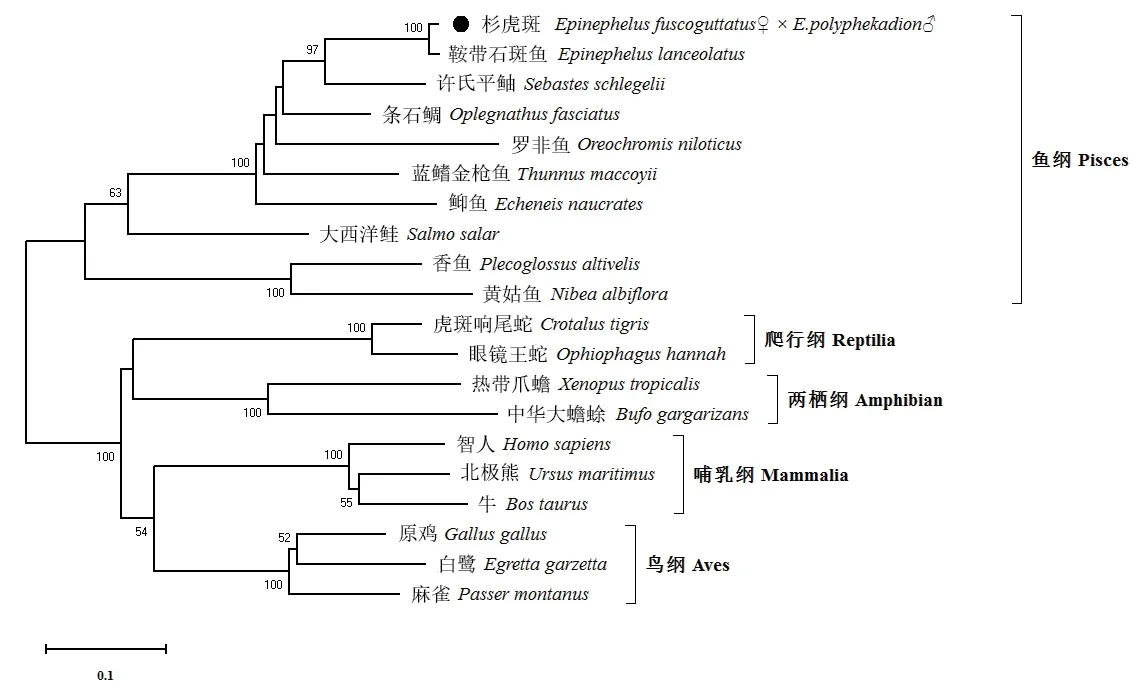
图4 邻接法构建Galectin-8 基因氨基酸序列的系统发育进化树Fig.4 Phylogenetic tree of the amino sequences of Galectin-8 genes constructed by neighbour-joining method
2.3 杉虎斑Galectin-8基因组织表达分析
利用RT-qPCR 分析了Galectin-8在杉虎斑不同组织类型中的表达情况。结果显示(图5),Galectin-8在所有被检测的组织中均有表达,且其在不同组织中的表达水平不同。Galectin-8在脾脏中表达量最高,其次是中肾、心脏、肠道、肝脏、头肾和鳃,而在脑、胃、皮肤和肌肉组织中的表达量相对较少。
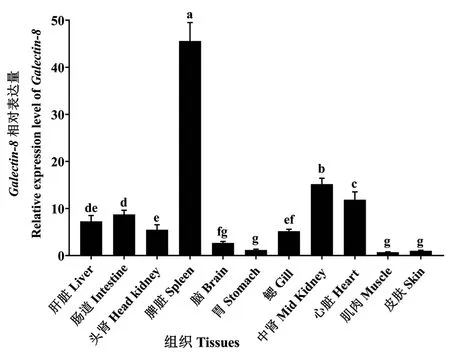
图5 杉虎斑Galectin-8 在各组织的相对表达量Fig.5 Relative expressions of Galectin-8 in the different tissues from hybrid grouper
2.4 饲料中添加不同水平阿魏酸对杉虎斑Galectin-8基因表达的影响
根据图6可知,杉虎斑Galectin-8基因的表达受到阿魏酸(FA)的影响。随着饲料中FA 水平的增加,Galectin-8表达量在肝脏、头肾和脾脏中均呈现先升高后降低的趋势。FA 添加水平为40、80、160 和320 mg/kg 时,肝脏、头肾和脾脏的Galectin-8表达量均显著高于对照组和高剂量添加组(640 与1 280 mg/kg)(P< 0.05)。在肠道中,与对照组相比,添加FA 会显著上调Galectin-8的表达量,其中FA 水平为1 280 mg/kg 时表达量最高,并显著高于其他组(P< 0.05)。
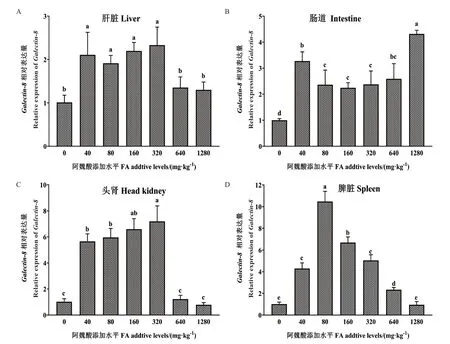
图6 饲料阿魏酸不同添加水平对杉虎斑肝脏、肠道、头肾和脾脏的Galectin-8 基因表达量的影响Fig.6 Effects of dietary supplementation of ferulic acid(FA)on the relative mRNA expressions of Galectin-8 in the liver,intestine,head kidney and spleen of hybrid grouper
3 讨论
凝集素是先天免疫系统中的重要的体液组成成分,具有一个或多个CRD,可作为模式识别受体(pattern recognition receptors,PRRs),识别潜在致病细菌细胞表面上的病原体相关分子模式(pathogenassociated molecular patterns,PAMPs),在先天免疫和抗病方面起着重要作用[17,29-31]。Galectin-8 具有多种功能,包括促进浆细胞形成、T 细胞和B 细胞增殖、肿瘤调控、凋亡诱导以及免疫调节[17]。本研究描述了杉虎斑Galectin-8的序列特征,检测了其组织分布,并探究不同FA 水平对其表达量的影响。
人类Galectin-8基因通过可变剪接产生14 种不同的转录本,共编码6 种蛋白质,并且大多数转录本具有短的5′UTR 和长的3′UTR[11]。通过RACE 技术获知杉虎斑Galectin-8基因的cDNA 全长为1 368 bp,5′UTR 为16 bp,3′UTR 为394 bp,共编码319个氨基酸。半乳糖凝集素家族成员不含信号肽序列,主要定位于细胞质和细胞核,然而它们可通过非经典途径分泌到细胞外,如直接易位和细胞外囊泡中释放,具有细胞内和细胞外的功能[9,32]。此外,半乳糖凝集素的特定功能取决于其表达的细胞类型与在细胞或细胞外的定位,因此它们的分泌必须受到严格的调控才能正常发挥作用[33]。序列分析表明,杉虎斑Galectin-8 是一种分泌蛋白,但未发现跨膜结构域与信号肽,主要位于细胞质中。目前,半乳糖凝集素分泌的机制尚不清楚,为了对Galectin-8 的分泌有一个全面的了解,还需要进一步的研究。基于结构域分析,发现杉虎斑Galectin-8 属于串联重复类型半乳糖凝集素,包含两个由36 个氨基酸的连接肽所连接的CRD,位于蛋白质序列的两端。同时,CRD 结构域中有两个保守的基序,H-NPR 和WGEE,这与其他半乳糖凝集素家族成员一致[11]。翻译后修饰分析显示杉虎斑Galectin-8 中有1 个潜在的N-糖基化位点和29 个预测的磷酸化位点。在小鼠的Galectin-8 中已观察到广泛的翻译后修饰,包括糖基化与磷酸化[34]。多重比对结果表明,在Galectin-8中两个β-半乳糖苷类结合基序在进化中是高度保守的;此外,虽然各物种的连接肽之间无序列同源性,但该区域中的氨基酸数量会导致蛋白质大小的变化[20]。系统进化树分析显示,杉虎斑Galectin-8与其他硬骨鱼类单独聚为一支,其中与鞍带石斑鱼的亲缘关系最近,而爬行类、两栖类、哺乳类和鸟类动物分别聚为一支。
在鱼类中,已发现半乳糖凝集素在多种组织和细胞中表达[20]。本文中杉虎斑Galectin-8在检测的组织中均有不同程度的表达,其中在脾脏的表达量最高。这与先前的研究一致,罗非鱼和牙鲆的Galectin-8也在脾脏中具有最高的表达量[18,21]。同时在头肾和肝脏中,杉虎斑Galectin-8也有较高的表达。脾脏、头肾和肝脏是鱼类重要的免疫器官[35]。鱼的脾脏也是一个重要的周围淋巴器官,在抗原捕获中起着关键作用,且与头肾相比,脾脏是体液免疫反应的主要器官[36]。头肾类似于哺乳动物的骨髓,具有造血功能,可负责吞噬、抗原处理以及IgM 的形成[37]。尽管脊椎动物的肝脏传统上被视为代谢、营养储存和解毒中心,但它还是一个重要的免疫器官,产生细胞因子、趋化因子、补体成分和急性时相蛋白以应对病原体感染[38]。此外,杉虎斑Galectin-8在脑、鳃和肠道中有着相对较高的表达量。脑中的小胶质细胞可积极参与对损伤和致病性感染的快速应答,而鳃和肠道属于先天免疫系统中的物理屏障[36,39]。综上所述,这些结果提示Galectin-8可能在杉虎斑的免疫系统中发挥作用[13,18]。
免疫刺激剂是一种能增强防御机制或免疫系统(先天性和适应性)的物质,从而使动物对疾病和外部胁迫有更强的抵抗力[40]。一些研究认为,功能性饲料添加剂可以通过两种方式激活水生动物的先天免疫系统:(1)直接刺激先天免疫系统;(2)促进共生微生物菌群的生长。饲料添加剂对鱼类的免疫刺激作用评估主要集中在非特异性免疫参数上[41]。FA 是一种有效的免疫刺激剂,并且被认为是提高动物免疫力和改善动物生理条件的最重要的添加剂之一,这部分归因于其抗氧化能力对细胞的保护作用[42-43]。本研究中,与对照组相比,饲料中添加FA 为40-320 mg/kg 时,肠道、肝脏、头肾和脾脏的Galectin-8表达量显著上调。类似地,金头鲷(Sparus aurataL.)摄入含有NEXT ENHANCE®150(由香芹酚和百里香酚组成)的试验饲料后,Galectin-1在后肠的表达量显著增加[44];用添加椰枣提取物的饲料喂养欧洲海鲈(Dicentrarchus labrax)4 周后,岩藻糖结合凝集素的表达量显著上调[45]。半乳糖凝集素转录水平的增加,能够促进微生物的识别和启动下游免疫反应,如活化T 细胞的增殖、细胞因子的产生、补体激活和白细胞募集[20]。因此,FA 能够促进Galectin-8基因的表达,这表明杉虎斑的先天免疫得到增强。然而,较高水平的FA(640 与1 280 mg/kg)对Galectin-8的表达量没有显著影响。有报道称,长期补充高浓度的免疫刺激剂可能会导致免疫抑制[46]。此外,Galectin-8 在杉虎斑中的免疫功能尚不清楚。因此需要进一步的研究来阐明杉虎斑Galectin-8 的免疫调节功能,以确定其在防御病原体入侵中的作用。
4 结论
本研究克隆的杉虎斑Galectin-8 属于串联重复型半乳糖凝集素,N 端与C 端的CRD 由连接肽所连接,与鞍带石斑鱼具有同源性和高度的一致性,并广泛表达于各种组织中,其中脾脏的表达量最高。饲料中添加FA 为40-320 mg/kg 时显著上调肠道、肝脏、头肾和脾脏的Galectin-8表达量,表明适量添加免疫刺激剂FA 能够增强杉虎斑的先天性免疫。
