辣椒适应非生物胁迫的研究进展
2023-01-05胡华冉杜磊张芮豪钟秋月刘发万桂敏
胡华冉 杜磊 张芮豪 钟秋月 刘发万 桂敏
(云南省农业科学院园艺作物研究所,昆明 650205)
辣椒(Capsicum annuumL.)因其丰富的营养价值和良好的风味深受消费者喜爱。除鲜食外,还可干制、酱制和泡制等,加工成辣椒粉、辣椒圈和辣椒酥等调味品,也可深加工提取辣椒素和辣椒红色素等产品,用于食品、饮料、饲料、保健药品、化妆品、医学及军事等领域[1-2]。根据国家特色蔬菜产业技术体系统计数据,我国辣椒播种面积在蔬菜作物中位居第一,成为我国蔬菜产业中第一大产业[3]。伴随着辣椒用途的不断开发和加工型产业的快速发展,辣椒成为我国乡村振兴和精准脱贫的重要抓手。
我国作为辣椒种植大国,对辣椒的研究及开发高度重视。目前,对辣椒的研究主要集中在辣椒种植与栽培、病虫害防治、辣椒性状基因定位、辣椒的药理及风味物质等几个方面。然而,辣椒在生长发育过程中不仅受到生物胁迫,还受到非生物因子的影响,导致辣椒的品质破坏、产量降低,所以对其抗性方面的研究很有必要。因此,本研究主要总结了近几年内辣椒在萌发期和苗期响应非生物胁迫的研究进展,分别在形态、生理生化、分子层面以及缓解措施等方面对其研究近况进行综述,有助于较全面了解辣椒在非生物胁迫条件下的最新研究进展,分析其存在的问题与未来的研究方向,以期为辣椒抗逆品种的培育及开发利用提供参考。
1 温度胁迫
1.1 辣椒耐高低温材料的筛选
温度胁迫包括高温胁迫和低温胁迫,危害作物的生长发育,造成作物减产,筛选耐高低温的品种具有重要的生产意义,相关研究主要集中于以萌发期指标、苗期形态指标和生理生化指标为依据进行筛选。王静等[4]对20 份辣椒材料的生理指标进行分析,筛选高度耐热、中度耐热、不耐热和极不耐热4 个等级品种。张慧静等[5]发现不同制干辣椒品种在胁迫条件下发芽势、发芽率、发芽指数的差异性显著,并利用这些指标鉴定品种间的耐冷性。胡能兵等[6]通过生理指标的隶属度分析法和叶绿素荧光参数降幅的排名获得可用于抗(耐)高温的辣椒品种。而通过辣椒资源种子萌发期、幼苗及坐果期的相关形态指标和生理生化特性综合分析,筛选耐高温材料的研究也有报道[7]。此外,苟秉调等[8]在低温弱光胁迫下获得可用于鉴定和预测辣椒杂种优势的光合指标,为预测低温胁迫下的辣椒杂种优势提供了理论依据。
1.2 生理指标响应
温度胁迫对辣椒生理生化的影响主要表现在渗透调节物质、保护酶系统、膜脂过氧化和光合作用上。张子学等[9]研究发现,经高、低温胁迫辣椒叶片中的可溶性糖(soluble sugars,SS)、脯氨酸(proline,Pro)和丙二醛(malondialdehyde,MDA)含量品种之间均存在极显著差异,其中,SS、Pro 含量大幅度提高、MDA 含量明显下降的品种,其耐热或抗寒能力强。潘宝贵[10]研究发现随着高温胁迫温度的升高和胁迫时间的延长,净光合速率迅速下降,气孔导度先降后升,超氧化物歧化酶(superoxide dismutase,SOD)活性急剧下降,过氧化氢酶(catalase,CAT)和抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性也随之下降,而过氧化物酶(peroxidase,POD)活性持续上升且增加幅度较大,耐热品种叶片中的酶活性(SOD、POD、APX、CAT)>耐热性中等的品种>热敏品种。该结果同何铁光等[11]、马宝鹏等[12]结果相似,说明高温胁迫下,氧化酶活性以及MDA、Pro 含量的变化在不同辣椒品种中有较大的差异,可作为辣椒耐热鉴定的辅助生理指标。
1.3 抗冷耐热基因的研究
在形态生理研究的基础上,学者开始在分子水平上对辣椒进行深入研究,目前已克隆得到多个与温度胁迫相关的基因。例如,WRKY 家族基因CaWRKY13、CaWRKY27和CaWRKY41,HSP 家族基因CaHSP18、CaHSP24、CaHSP26和HSP90,以及CaERF18、CaERF109、CaSYT5、KCS、NAT、CaVIN02和CaCWIN03,通过生物信息学与表达量分析发现,这些基因对低温和高温具有较强的响应[13-21]。有学者对这些基因的机理进行了初步的探索,姬俏华[22]利用农杆菌介导的遗传转化方法获得了CaWRKY20的过表达烟草植株,发现过表达CaWRKY20可显著提高烟草对高温的耐受性。颜坤[23]研究发现高温胁迫下转正义CaGPAT烟草的光合能力强于野生型。而郭尚敬[24]从甜椒叶片中分离到小分子量热激蛋白基因CaHSP26和CaHSP18,低温条件下过量表达对烟草、大肠杆菌均有保护作用。此外,也有学者探讨DNA 甲基化在辣椒高温多湿胁迫反应中的作用[25],以及在转录水平和转录后水平上阐明CaWRKY40介导的辣椒应答RSI 和HTHH 信号通路相互作用分子机制[26]。Li 等[27]通过比较转录组分析揭示了热胁迫下敏感辣椒和耐热辣椒的转录差异。上述基因在提高植物的温度胁迫耐受性方面有一定的作用,可用于辣椒抗逆基因工程改良研究。辣椒温度胁迫相关研究见表1。
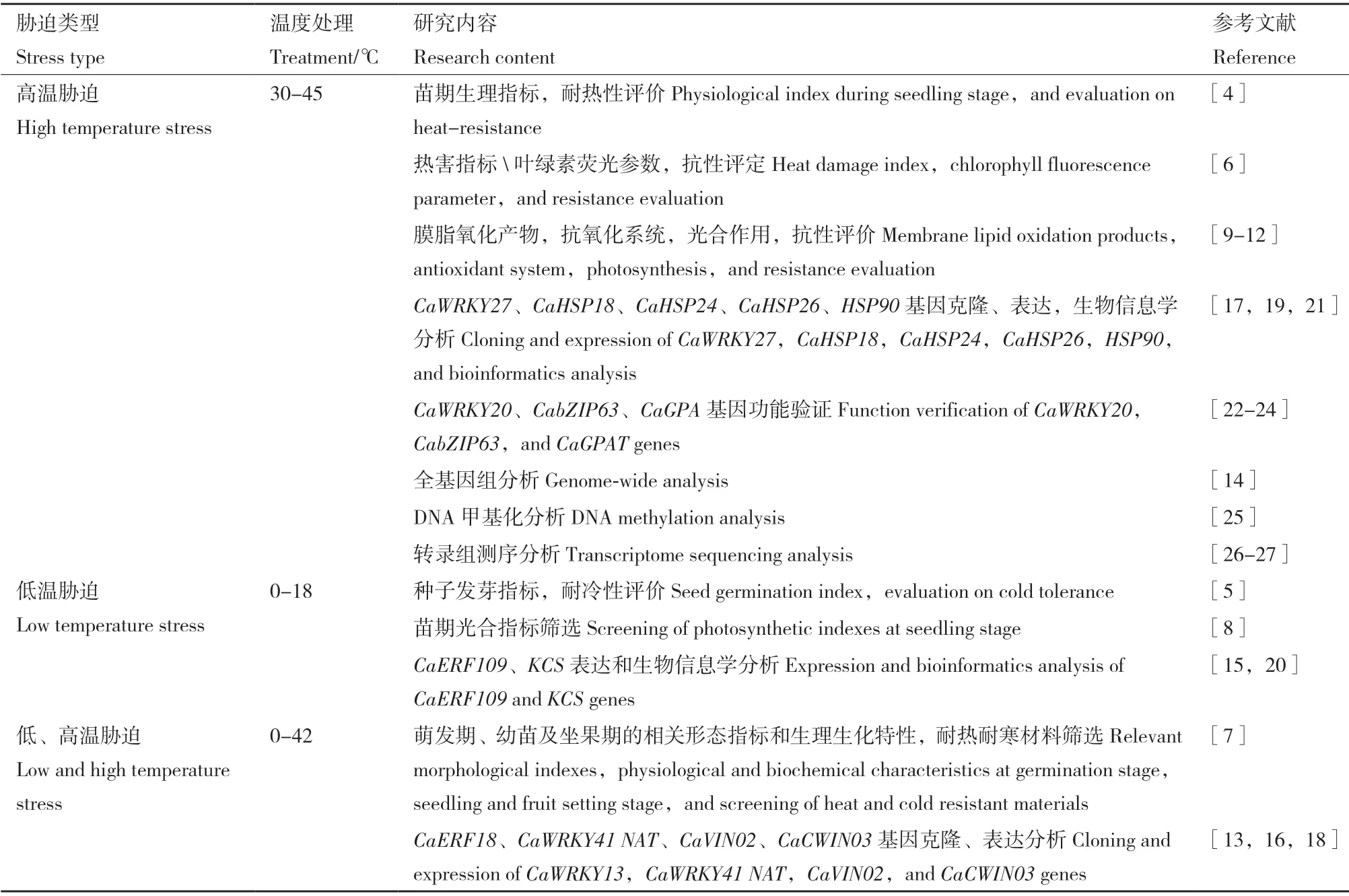
表1 辣椒温度胁迫研究进展Table 1 Recent studies of temperature stress in hot pepper
2 盐碱胁迫
种子萌发期和苗期是植物对盐分最敏感的时期,因而辣椒耐盐性研究主要集中在这两个时期。研究表明,盐、碱胁迫使辣椒的发芽势、发芽率、发芽指数、活力指数及相对盐害率等指标降低,抑制胚芽及胚根的生长,主根长、侧根长与侧根分级数以及子叶展开百分比与盐碱胁迫呈明显负相关[28-29],幼苗株高、干重和根长显著下降[30],导致盐碱胁迫降低了辣椒的产量,影响辣椒果实品质,且碱性盐的抑制效应大于中性盐,强碱(Na2CO3)比弱碱(NaHCO3)的抑制作用更强[31-32]。此外,低浓度的中性盐、碱性盐对辣椒种子发芽势均有促进作用。还有学者发现土壤盐分影响辣椒果实的辣椒素含量[33]。
2.1 辣椒耐盐性评价研究
植物的耐盐性有不同程度的表现,且不同辣椒品种在不同指标上表现各有优劣[34-35],目前,大多研究以形态生理指标鉴定品种间或不同地区辣椒材料间的耐盐性为主[36]。逯明辉等[37]发现,相对发芽势、相对发芽率和相对发芽指数等3 个指标均能区分不同辣椒品种的耐盐性,耐盐性由强到弱依次为尖椒类型>朝天椒类型和线椒类型>甜椒类型。郭春蕊等[38]研究发现,相对发芽指数和相对盐害率也宜作辣椒发芽耐盐筛选指标,而株高、地上部鲜重和干重,叶绿素、MDA、相对电导率、酶活性(CAT、SOD、POD)、渗透调节物质SS、Pro、可溶性蛋白(soluble protein,SP)等指标可用来筛选辣椒幼苗的耐盐性[39-41]。
2.2 生理指标响应
在生理水平上,盐碱胁迫使渗透调节物质(Pro、甜菜碱、有机酸)含量增加,抗氧化物质抗坏血酸(ascorbic acid,ASA)、还原型谷胱甘肽(reduced glutathione,GSH)、维他命E(vitamin E,VE)增加,保护酶活性(POD、SOD、CAT)增加,K+、Na+含量的变化与膜透性(MDA)的变化一致增加[42]。张弢[43]、吉雪花等[44]研究结果与以上研究基本一致。而郑佳秋等[45]发现在盐胁迫下所有供试辣椒品种在幼苗期的SS 和SP 含量总体呈现下降趋势,不同品种SOD 表现也不同。光合性能指数(如实际光化学效率、表观光合电子传递速率、最大光化学效率和光化学猝灭系数等)均能反应盐胁迫对辣椒叶片光合性能的影响[46-47]。李汉钊等[48]、刘会芳等[49]发现较高的盐浓度对辣椒的光合作用及荧光参数有明显的抑制作用。
2.3 辣椒应对盐碱胁迫的分子机制探讨
目前,有学者对辣椒疑似耐盐基因CaCBF1A、CabZIP1、CaNAC61、CaCBF1A进行克隆与分析,推测其在辣椒抗逆机制中起着重要的作用[53-55]。冯冬林[56]、尹延旭[57]通过转基因分别获得CaERF14、CaPIP1-1的转基因株系,其对盐胁迫的耐受力均显著提高。也有学者通过转录组学探索了线辣椒的有关耐盐机理[58]。由于土壤盐化和碱化经常同时发生,因此,也有学者从生理和转录水平阐明辣椒幼苗对中性盐、碱性盐、混合盐胁迫的差异响应机理。李慧姬[59]对盐、碱胁迫不同时间点的辣椒幼苗叶片进行转录组测序发现,盐碱差异基因主要富集在光合作用、渗透调节物质合成、氧化还原过程、氮代谢及有机酸代谢等过程。辣椒盐胁迫相关研究如表2所示。
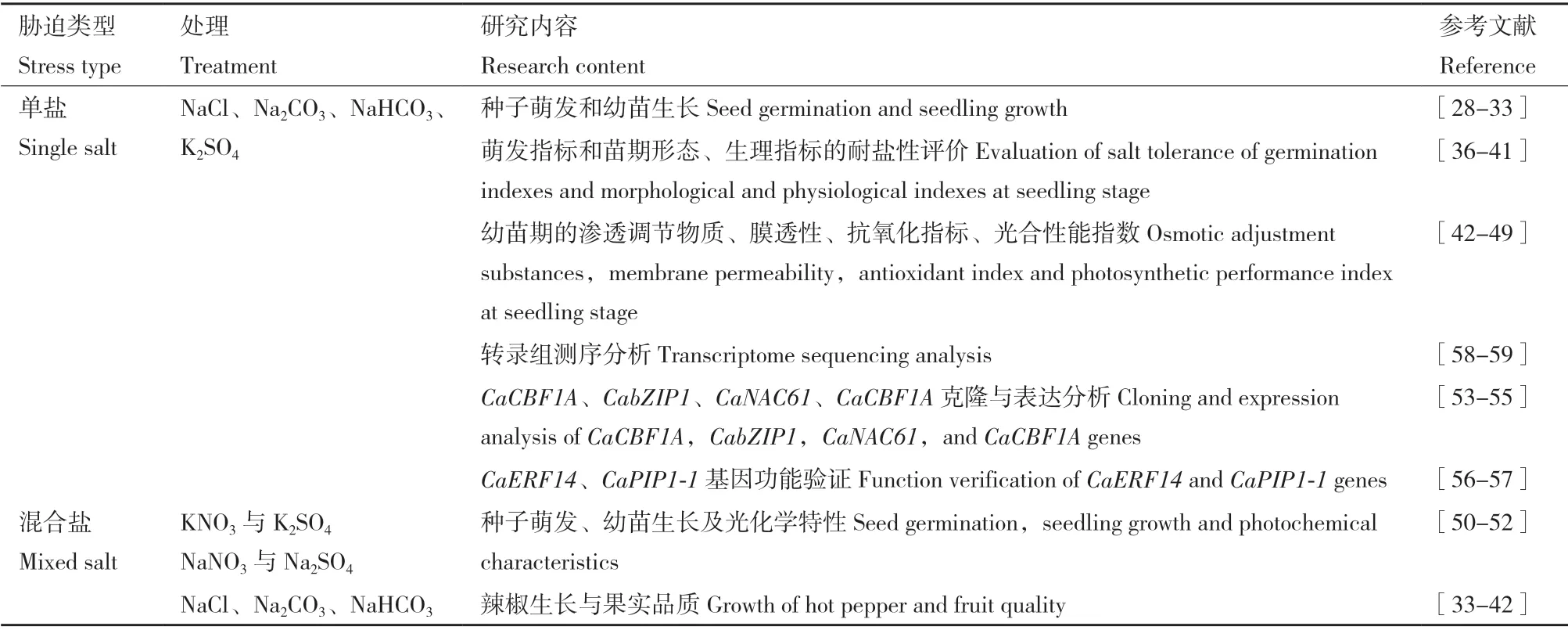
表2 辣椒盐碱胁迫研究进展Table 2 Recent studies of saline-alkali stress to hot pepper
3 水分胁迫
3.1 干旱胁迫
干旱影响辣椒各阶段的生长发育和生理代谢过程,进而影响辣椒的产量和质量[60]。近年来,有关辣椒抗旱胁迫的研究较少,主要集中在辣椒响应干旱胁迫的生理生化特性的研究。有学者研究发现当辣椒植株缺水时,细胞膨压降低导致细胞伸长生长受到抑制,因而叶片较小,辣椒茎叶形态、根系、根冠比均会受到影响[61-63]。随着胁迫程度的增加,水势明显降低,且细胞内脱落酸含量增高,使净光合率随之下降[64]。刘颖等[65]进一步研究发现辣椒地下部分对干旱胁迫的敏感度高于地上部分。朱冉冉[66]研究结果表明辣椒通过提升叶片抗氧化酶活性、Pro 含量、氧化还原酶活性、SP 含量使辣椒抗旱性得到进一步增强,此结果与娄喜艳等[67]、Okunlola 等[68]研究结果一致。总体上抗旱性强的品种组织相对含水量、叶绿素含量和Pro含量相对较高,而质膜透性和MDA 含量相对较低。此外,干旱影响辣椒的果实品质,不仅降低辣椒的果长、果粗、茎粗、株高、单果重以及干物质含量,而且其代谢物质如粗脂肪、粗纤维含量及辣椒素含量较CK 处理显著下降[69],Ricardez-Miranda 等[70]在水分胁迫对辣椒生长、次生代谢产物和果实产量的研究中也得到类似的结果,但适度干旱可促进光合以及使α-胡萝卜素、β-胡萝卜素等上游物质含量显著增加[66]。
3.2 耐涝胁迫
在淹水条件下,植株有氧呼吸受到抑制,水分难以吸收,蒸腾作用降低,细胞缺水失去膨压,最终萎蔫。目前,集中于辣椒耐涝胁迫的研究甚少,有学者对耐涝胁迫下辣椒的生理响应进行了研究。Martínez-Acosta 等[71]研究发现淹水处理对植株的气孔导度、光合和蒸腾速率均产生一定的抑制作用,导致植株的生物量显著下降。付秋实等[72]研究了水分胁迫对辣椒光合作用及相关生理特性的影响,结果表明,在水分胁迫下辣椒植株总干物质含量、叶片水势、叶片相对含水量、叶绿素以及类胡萝卜素含量显著下降。植物在淹水条件下,保护酶系统会被破坏,自由基就会大量产生,造成膜脂过氧化,MDA 含量大幅增加。刘佳等[73]研究结果表明,随着处理时间的延长,辣椒叶片中MDA 的含量呈上升趋势,SOD 和POD 均呈现先升高后降低的趋势。
有关建立辣椒耐涝性评价体系的研究也有所报道,宋钊等[74]从辣椒受到涝渍胁迫后的植株死亡率、叶片颜色、茎基部淹水处的颜色与形态4 个方面建立辣椒耐涝性鉴定评价体系,并通过筛选高耐涝品种和涝渍敏感品种,证实了这个评价体系有效、可行。辣椒水分胁迫相关研究如表3所示。

表3 辣椒水分胁迫研究进展Table 3 Recent studies of water stress in hot pepper
4 重金属胁迫
4.1 重金属对辣椒种子萌发和幼苗生长的影响
目前,由于环境污染,植物受到重金属毒害的现象极为普遍,国内外学者就植物耐受重金属胁迫方面进行了大量研究,但对辣椒重金属耐受性的研究较少,目前的研究主要集中在重金属镉(Cd)、铅(Pb)、铬(Cr)离子对辣椒种子萌发和幼苗生长的影响。高浓度Cd2+(≥20 mg/L)不仅会降低辣椒种子发芽率,抑制辣椒胚根、胚轴生长,还会造成辣椒幼苗植株矮小,抑制植株生长和根系长度,苗鲜重减少[75];同时Cd2+毒害使辣椒幼苗叶片MDA 含量增加,SOD、CAT 活性下降[76]。有学者还发现辣椒的其他指标,如株高、茎粗、展叶数、叶面积、辣椒叶片的光合指标也会在重金属处理下降低[77]。此外,低浓度Cd2+、Pb 可促进辣椒植株的生长[78-79]。
4.2 辣椒对重金属的富集作用
辣椒具有较强的吸收和富集重金属的能力,因此,有学者研究辣椒不同品种、不同部位、以及不同部位中亚细胞对重金属的吸收富集作用,多数学者发现辣椒对重金属(Cd、Pb、As)的富集表现为根部最强[79-81]。杨晓磊等[82]通过植株中Cd 的富集系数研究发现,重金属的迁移是通过根部吸收后由茎叶转运,分布浓度依次为根系>茎叶>果实,苏园[83]研究结果与此一致。此外,品种和土壤不同,辣椒重金属转移、富集能力均有所改变。有学者发现辣椒各部位镉的分布在黄壤上表现为根>茎、叶>果实,石灰(岩)土中为根、叶>果实>茎;不同辣椒类型的镉含量也存在差异,根部为线椒>朝天椒、杂交椒,叶和果实为朝天椒>线椒、杂交椒[84]。谈敏等[85]通过对镉在辣椒品种间以及辣椒不同器官的富集差异、镉对辣椒理化性状的影响、外源物质对辣椒吸收富集镉的调控4 个方面进行了探讨和概述。
4.3 重金属胁迫相关基因的研究
目前,有关辣椒重金属胁迫在分子方面的报道较少。刘海波等[86]从辣椒中分离到一个能够响应热胁迫的金属伴侣蛋白基因CaHPP7,其表达受Cu2+、Cd2+等重金属的诱导;利用基因沉默技术发现辣椒对铜胁迫的抗性都降低;而基因过表达后,铜胁迫下表现为种子发芽率高于对照,幼苗生长的受抑制程度和叶圆片叶绿素含量下降程度均低于对照。故推测CaHPP7在植物应对铜胁迫的过程中起正调控作用。此外,李桃[87]不仅探究了3 个品种辣椒营养器官和果实的生长及生理效应、Cd 吸收、迁移富集及积累情况,也进一步研究了耐性相关基因表达量的变化,初步探明了辣椒低积累Cd 的分子机理,并获得Cd 积累/耐性过程的关键基因。辣椒重金属胁迫研究进展如表4所示。

表4 辣椒重金属胁迫研究进展Table 4 Recent studies of heavy-metal stress in hot pepper
5 改良措施
多年来,我国学者不仅在生理和分子水平上对辣椒各种胁迫(温度、盐碱、水分、干旱、重金属)指标进行了系统研究,同时也对如何缓解这些逆境进行了探索。其中,浸种、叶面喷施是常用的方式。
结合自己的“下水”心得和做题情境,我倾向于第三类同学的做法,因为有的放矢确实可以让阅读的目光变得更敏锐,提笔标画更有方向性和侧重点(其实关于不动笔墨不读书的道理不难懂,问题是——非知之艰,惟行之艰),不会出现要么没重点,要么处处是重点而变成没重点,耗时耗力,答题效率低下。
5.1 降低温度胁迫的改良方法
通过叶面喷施不同浓度的5-氨基乙酰丙酸(ALA)、亚精胺(Spd)、氯化钙(CaCl2)、壳聚糖(CS)、聚乙二醇(PEG)、蔗糖及2,4-表油菜素内酯(2,4-epibrassinolide,EBR),均可以降低辣椒低温胁迫[88-91]。叶面喷施沼液肥20 倍液可以显著降低热害指数[92]。还有学者发现,外源施钙以及喷施适宜浓度的1-甲基环丙烯或者经过6-BA 处理均可以减少高温胁迫对辣椒作物带来的伤害[93-95]。此外,通过嫁接的方式也可以使嫁接苗分别对高温和低温胁迫具有较好的适应性[96]。
5.2 降低盐碱胁迫的改良方法
为缓解盐胁迫对辣椒的损害,许多学者通过施加外源物质如水杨酸、硝普钠、油菜素内酯以及通过赤霉素浸种来降低盐害。有研究发现水杨酸不仅能够缓解盐胁迫对辣椒幼苗的生长抑制,促进叶片叶绿素的合成,保护其光合作用,维持植物的正常生长[97-98],还可以增加辣椒幼苗渗透物质的含量和抗氧化酶活性,从而提高其抗盐能力[99-100]。但超过一定浓度则会抑制辣椒种子的萌发,使种子浸出液电导率和MDA 含量升高[101]。一定浓度赤霉素浸种和硝普钠处理可以缓解辣椒种子盐胁迫,提高种子的发芽率、发芽势和发芽指数,低浓度时有促进作用,高浓度时抑制[102-103]。此外,采用腐植酸和硝酸钙、生物炭基Rs-198 菌剂也可以减轻盐胁迫对辣椒造成的危害[104]。
5.3 降低旱涝胁迫的改良方法
已有研究发现通过添加化学试剂水杨酸、一氧化氮、5-氨基乙酰丙酸、茉莉酸甲酯等,均可以提高辣椒抗旱能力[105-107]。而Matmarurat 等[108]通过对辣椒种子进行200 Gy 的γ 射线辐照,以提高辣椒的耐旱性。Neto 等[109]通过使用牛生物肥料、沟槽侧覆聚乙烯薄膜、直接土壤覆盖的方式,来缓解干旱地区的水分蒸发问题。结果表明,在甜椒栽培中,使用沟槽侧覆可以提高产量,提高生产率;但牛生物肥料×侧覆×土壤覆盖的相互作用会抑制植物中叶绿素的产生。另外,通过外源Ca2+浸种处理,调节辣椒幼苗根系内呼吸代谢可以缓解淹水胁迫对植株的伤害,改善根系(根系总长度、总表面积、体积、根尖数)的生长状况,提高根系活力[110-111]。
5.4 降低重金属胁迫的改良方法
目前,土壤重金属污染的治理包括工程法、植物修复法和农艺调控法等。部分学者提出施加组配改良剂来改善辣椒对重金属的吸收。例如,施加不同添加量的组配改良剂海泡石+石灰石后,与对照相比,辣椒可食部位和根部重金属Pb、Cd、Cu、Zn 含量明显降低[112]。叶面喷施不同浓度的Si、P、Zn,会缓解镉对辣椒的毒害,促进其生长[113]。此外,在镉或铝环境下,施用一定浓度的有机肥、双氰胺和石灰、外源钙,可以缓解辣椒植株的金属胁迫,并提高辣椒的果实品质[114-115]。
通过以上文献分析发现,缓解辣椒非生物胁迫的措施存在一个共同点,在非生物胁迫下,外源物质处理的辣椒幼苗抗逆性与叶绿素含量、保护酶活性和渗透调节物质的含量均有密切关系,可以通过测定生长指标(苗高、茎粗、根鲜重、根干重、地上部鲜重、地上部干重等)和生理指标(SS、SP、Pro、MDA、SOD 和POD 等)来鉴定外源物质对提高辣椒幼苗抗逆的作用效果,获得其生理改良机制。辣椒非生物胁迫的改良方法如表5所示。

表5 辣椒非生物胁迫的改良方法Table 5 Improved methods of abiotic stress in pepper
6 展望
我国是世界第一大辣椒(含甜椒)生产国与消费国,近年来全球气候变化对辣椒生产带来的影响不容忽视,因此,加强辣椒抗逆性方面的研究具有重要的意义。截至目前,关于辣椒非生物胁迫的研究虽然已经取得了一定的进展,但与其他重要的农作物相比,仍然差距较大,今后可从以下几个方面开展研究工作。
在提高辣椒对逆境的耐受能力方面,我国研究相对薄弱,尤其是通过探索植物的抗逆机理并利用现代生物技术,改变其遗传基础来提高耐受性方面仍处于初级阶段。目前,已经利用现代分子生物技术等手段,从辣椒中发掘到一些抗逆相关基因,下一步则需着重阐明其在植物体内行使的功能并对此基因进行克隆并导入其他高产辣椒,从而使辣椒间的优良基因结合,最终培育出品质好、产量高、抗性强的辣椒新品种。但受品种遗传性、环境因素、激素配比、操作方法等因素的影响,辣椒组培体系并未十分成熟,难以建立完善的转化体系,因此,抗逆基因的导入任务有一定困难,需要加强辣椒抗性基因的挖掘与利用。
辣椒的生长受多因素的影响,其响应变化是相互协同或者相对独立仍需进一步思考。有报道称,辣椒属于干旱敏感、适度耐盐的蔬菜作物,其生长发育容易受到干旱以及由过度施肥造成的盐分胁迫影响。有关盐碱、温度及水分胁迫两两互作或三者互作对辣椒生长发育在生理水平的影响已有相关研究,但在分子层面上对其进行深入研究的报道并不多,对辣椒抗逆性机理的研究可以作为后续的一个研究方向。同时在机理研究的基础上,筛选与辣椒抗逆性有关的分子标记或基因,从而加快辣椒抗逆性品种的选育。
不同生育期的辣椒对非生物胁迫的响应程度不同,目前大部分的研究都集中在对苗期的影响上,而对成株期影响的研究较少,特别是对采收期辣椒果实产量、品质如何响应非生物胁迫缺少全面的认识,所以还需要进一步的研究。
通过前面综述可知,关于辣椒逆境胁迫条件下激素水平变化规律及其对耐涝、重金属胁迫的响应的研究不仅数量少,而且多数停留在生理水平。因此,需要进行调控机理的深入研究,进一步为辣椒抗性育种的亲本选择提供理论依据。
