谷物花色苷功能活性与开发利用研究进展
2023-01-03刘婷婷彭文婷庞邵杰彭珍珍綦文涛
刘婷婷,彭文婷,庞邵杰,彭珍珍,3,綦文涛, ,王 勇,
(1.国家粮食和物资储备局科学研究院,北京 100037;2.上海理工大学健康科学与工程学院,上海 200093;3.中南林业科技大学食品科学与工程学院,湖南长沙 410004)
谷物是居民膳食的重要基础组成部分,研究表明,摄入全谷物及其产品可降低非传染性慢性病患病风险[1]。“中国居民膳食指南”建议每日全谷物与豆类摄入量为50~150 g,且每日增加摄入30 g全谷物,可降低8%全因死亡风险。谷物含有多种活性物质,其中天然多酚可作为单线态氧猝灭剂和自由基供氢体,发挥抗氧化和抗炎活性,能有效缓解并预防糖尿病、心血管疾病和癌症等慢性病[2]。膳食多酚以花色苷摄入量最高,主要存在于黑色、红色、紫色等深色谷物、蔬菜和水果中,花色苷可有效干预慢性病的发生和发展[3-4]。《中国居民膳食营养素参考摄入量》对花色苷摄入量的特定建议值(specific proposed levels,SPL)为50 mg/d,即每天摄入超过50 mg的花色苷可产生较为明显的健康促进作用[5]。
随着2型糖尿病、心血管疾病、肥胖等慢性病发病率逐年升高,充分挖掘谷物中的功能活性成分、探究作用机理、开发相关功能食品对干预慢性疾病具有重要意义。花色苷作为一种天然食用功能色素,在食品工业已得到广泛应用[6]。当前对于谷物花色苷的研究主要集中在提取纯化、吸收代谢、健康功效等方面,但花色苷发挥健康作用的具体调控机制及其应用尚不清晰。本文主要综述谷物来源花色苷的含量和分布、对慢性疾病的预防作用机制及开发利用现状,为进一步明确花色苷作用靶点、揭示花色苷营养干预机制及开发花色苷类功能产品提供研究思路。
1 花色苷在谷物中的分布与含量
谷物主要包括稻谷、小麦、玉米、燕麦、荞麦、大麦、青稞、高粱等。花色苷作为植物呈色的重要组分,广泛分布于深色谷物中,其化学结构、在植物中的分布和含量不同是导致植物颜色差异的主要原因。例如稻米花色苷主要分布于米糠层,研究发现稻谷籽粒抛光后仅保留总花色苷的3%[7]。小麦、大麦、玉米、高粱等谷物中花色苷分布较为相似,在红色及紫色品种的表皮、蓝色品种的糊粉层、黑色品种的表皮和糊粉层中均含有大量花色苷[8]。
花色苷含量差异受多种因素影响,包括基因型、生长条件、提取及量化方法等。目前检测谷物花色苷含量常用的方法有紫外可见分光光度法、高效液相色谱法、液相色谱质谱联用法等。不同谷物来源花色苷的分布和含量见表1,其中黑米、紫米花色苷含量远高于红米花色苷含量,矢车菊素-3-葡萄糖苷(cyanidin-3-glucoside,C3G)为黑米中最主要的花色苷,约占黑米中总花色苷含量的88%。紫玉米作为食品行业优质安全的天然色素来源,可用于多种焙烤、煎炸及膨化食品的生产。紫色小麦和大麦品种花色苷含量均高于其他有色品种,C3G是玉米、小麦等深色谷物的主要花色苷之一。3-脱氧花青素为高粱特有的花色苷,因其结构特性具有较强的稳定性,作为功能性天然食用色素被广泛应用于食品产业。
除谷物外,豆类、薯类等粮食中也含有大量花色苷,如黑豆花色苷含量可达201.9 mg/100 g[9],且主要花色苷为C3G,其约占黑豆总花色苷含量的95%。紫薯中花色苷含量为558~2477 mg/100 g[10],主要为酰化的芍药素和矢车菊素,糖基主要为槐糖苷和葡糖苷[11-12]。相比较而言,谷物由于水分含量低、易储存、易运输,且谷物作为膳食的重要基础组成部分,谷物花色苷具有来源广泛、摄入量高、稳定性强等特点,从谷物分离制备花色苷具有明显优势。
2 谷物花色苷功能活性研究
花色苷具有抗氧化、抗炎、改善视网膜毛细血管微循环、缓解肥胖、控制血糖、保护神经系统等功能活性。由于谷物花色苷结构特性,其与机体相关靶点受体的特异性和有效性结合作用,使谷物花色苷在促进人体健康方面具有突出表现。诸多研究表明摄入谷物花色苷可通过减轻机体氧化应激和炎症反应,预防或缓解糖尿病、肥胖、心血管疾病、年龄相关性眼病、癌症等多种慢性疾病(表2),相关机制主要涉及Nrf2、NF-κB、MAPK、PI3K/AKT等分子信号通路[30],花色苷对相关通路的调控机制总结见图1。
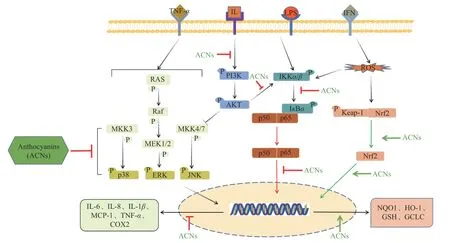
图 1 谷物花色苷对Nrf2、NF-κB、MAPK、PI3K/AKT信号通路的调控Fig.1 Regulation of cereal anthocyanins on Nrf2, NF-κB, MAPK and PI3K/AKT signaling pathways
2.1 抗炎症
炎症反应涉及多种通路,其中核因子-κB(nuclear factor-kappa B,NF-κB)通路是炎症反应经典通路。NF-κB蛋白由p65与p50形成二聚体,并与抑制蛋白IκB结合形成三聚体复合物而处于失活状态。当细胞受到刺激时会诱导相关信号因子磷酸化及核转移,从而激活下游炎症因子转录。多项研究表明花色苷可通过调控NF-κB信号通路发挥抗炎作用。黑米花色苷通过抑制NF-κB p65核转移及炎症因子分泌,缓解大鼠口腔黏膜炎及小鼠结肠炎症状[31-32]。Hathaichanok等[33]研究表明紫玉米花色苷通过降低NF-κB信号通中IKK、IκBα、p65磷酸化水平,减轻晚期糖基化终末产物(advanced glycation end products,AGEs)诱导的人关节软骨细胞炎症反应。
丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)家族蛋白在调节炎症反应中主要涉及p38丝裂原活化蛋白激酶、细胞外信号调节激酶(extracellular-signalregulated protein kinase,ERK)和c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)三条信号通路,磷酸化的p38、JNK和ERK分别激活相关转录因子,参与炎症反应、细胞增殖和凋亡等过程[34]。MAPK可与NF-κB经典途径产生协同作用,促进白介素1β(interleukin-1β,IL-1β)、IL-6、IL-8等基因表达[35]。Zhang等[36]研究表明红玉米花色苷通过调节IκBα和JNK的磷酸化水平,抑制NF-κB和MAPK信号通路,减少炎症因子产生,从而改善脂肪细胞和巨噬细胞之间旁分泌相互作用。紫米中含有丰富的C3G和芍药素-3-葡萄糖苷(peonidin-3-glucoside,Pn3G),Tunchanok等[37]利用紫米花色苷及其酚酸代谢物处理IL-1β诱导的人软骨细胞,可显著抑制IκBα降解,降低p-p65水平,并抑制MAPK信号通路激活,从而具有抗炎效果。在一项人群干预研究中,给予高胆固醇血症患者每日320 mg花色苷,24周后,其血浆C-反应蛋白(C-reactive protein,CRP)、血管细胞粘附分子-1(vascular cell adhesionmolecule-1,VCAM-1)和IL-1β水平分别下降20%、13%和4%[38]。NF-κB和MAPK是炎症反应的主要通路,谷物花色苷通过抑制相关通路降低炎症因子释放,缓解炎症反应。

表 1 不同谷物来源花色苷分布和含量Table 1 Distribution and content of anthocyanins from different cereals

表 2 谷物花色苷对多种疾病的预防作用及相关机制Table 2 The preventive effects of cereal anthocyanins on various diseases and its mechanism
2.2 抗氧化
谷物花色苷可通过清除活性氧(reactive oxygen species,ROS)及激活内源性抗氧化通路,抑制氧化应激。自由基过量产生和抗氧化防御系统失衡是导致氧化应激的主要原因,氧化应激影响细胞基因表达,进一步诱发炎症反应,引发多种疾病。正常情况下核因子E2相关因子2(nuclear factor erythroid 2-related factor 2,Nrf2)与Kelch样环氧氯丙烷相关蛋白1(Kelch-like ECH-associated protein 1,Keap1)在细胞质中结合成二聚体,发生氧化应激时会激活内源性抗氧化通路Keap1/Nrf2,Nrf2与Keap1解偶联并发生核移位,被抗氧化反应元件(antioxidant response element,ARE)识别并结合,启动靶基因转录,释放血红素氧合酶-1(heme oxygenase-1,HO-1)、醌氧化还原酶(NAD(P)H quinone oxidoreductase 1,NQO1)等抗氧化酶,抑制氧化损伤[39-40]。常世敏等[41]研究表明黑米花色苷相比于越橘花色苷具有更好的抗氧化性,是良好的天然抗氧化剂。紫色青稞花色苷提取物可有效降低CoCl2诱导的PC12细胞ROS水平,并提高过氧化氢酶(catalase,CAT)和超氧化物歧化酶(superoxide dismutase,SOD)水平,预防氧化应激诱导的细胞凋亡[24]。Phatchawan等[42]研究发现泰国黑米花色苷提取物(主要含有C3G和Pn3G)可降低肾损伤大鼠丙二醛(malondialdehyde,MDA)含量,提高SOD和谷胱甘肽(glutathione,GSH)等抗氧化酶水平,通过清除ROS、抑制Nrf2核转移减轻氧化应激水平。
2.3 视觉保护作用
视网膜处于高含氧量、高多不饱和脂肪酸、高光照的环境中,光氧化损伤会导致年龄相关性黄斑变性(age-related macular degeneration,AMD)等眼病的发生[43]。花色苷对视网膜的保护机制涉及多种途径,包括抑制N-亚视黄基-N-视黄基-乙醇胺(N-retinylidene-N-retinyl-ethanolamine,A2E)光氧化和裂解、改善4-羟基壬烯酸(4-hydroxynonenal,4-HNE)等脂质过氧化产物损伤、缓解内质网应激、调控Nrf2/HO-1、NF-κB、MAPK等信号通路[44]。C3G可以清除视网膜色素上皮(retinal pigment epithelial,RPE)细胞内ROS、降低炎症因子及凋亡蛋白表达,并抑制光氧化型A2E的形成和裂解[45]。Wang等[46]基于脂质过氧化物4-HNE诱导的RPE细胞损伤模型,发现C3G可抑制血管内皮生长因子(vascular endothelial growth factor,VEGF)过表达、降低炎症因子水平、缓解细胞衰老,提高RPE细胞存活率。基于大鼠视网膜光化学损伤模型,黑米花色苷可有效降低视网膜组织JNK-c-Jun/转录因子(activator protein-1,AP-1)和Caspase-1表达[47]。紫米花色苷可清除大鼠感光细胞ROS,抑制细胞凋亡[48]。一项双盲安慰剂对照的人群交叉研究发现,与安慰剂相比,每天50 mg花青素的摄入量可有效改善暗适应以及视疲劳症状[49]。大量研究表明花色苷可有效保护视力,维持视网膜正常功能。但研究大多集中于浆果来源花色苷,因谷物作为主食而被大量消费,具有极大的研究及应用潜力。
2.4 对肥胖的缓解作用
将富含花色苷的黑色、紫色小麦添加至高脂饲料,可提高小鼠体内与脂肪酸平衡、氧化应激相关的酶活性,显著降低小鼠体重及血清中总胆固醇、甘油三酯和游离脂肪酸水平[50]。Song等[51]研究发现膳食补充黑米花色苷可降低过氧化物酶体增殖物活化受体γ(peroxisome proliferator activated receptorγ,Pparγ)、凋亡相关因子(recombinant factor related apoptosis,Fas)和硬脂酰辅酶A去饱和酶1(stearoyl-CoA desaturase 1,Scd1)基因表达水平,提高Pparα、酰基辅酶A氧化酶1(acyl-coenzyme A oxidase 1,Acox1)和脂联素受体2(adiponectin receptor 2,AdipoR2)基因表达,通过调节Pparγ信号通路减少脂肪生成并促进脂肪酸氧化,调节肠道微生物群和脂质代谢,从而预防肥胖相关的高脂血症、肝脂肪变性和胰岛素抵抗。Wang和Liu等[52-53]研究发现黑米花色苷具有调节胆固醇代谢和肠道菌群的健康功效,降低血脂异常和脂肪肝患病风险。此外,紫玉米花色苷提取物通过调控NF-κB、AMPK信号通路降低IL-12、单核细胞趋化因子-1(monocyte chemoattractant protein-1,MCP-1)等炎症因子表达和胰岛素抵抗,降低高脂饮食诱导的小鼠脂肪生成并促进能量消耗[54]。以上结果表明饮食摄入谷物花色苷可有效预防肥胖,近期研究显示相关机制可能涉及花色苷与肠道菌群的相互作用,花色苷通过改善肠道菌群缓解肥胖的相关机制尚需进一步研究。
2.5 对糖尿病的干预作用
糖尿病是一种慢性内分泌系统疾病,病症直观表现为高血糖、胰岛素抵抗,花色苷可通过调节糖代谢和胰岛素分泌缓解糖尿病症状[55]。Diego等[56]基于细胞共培养技术,研究紫玉米皮花色苷提取物对胰腺细胞胰岛素分泌和肝细胞葡萄糖吸收能力的影响,结果表明紫玉米花色苷可以增强胰腺细胞胰岛素分泌能力,提高游离脂肪酸受体1(free fatty acid receptor 1,FFAR1)和葡萄糖激酶(glucokinase,GK)的活性,降低HepG2细胞中腺苷酸激活蛋白激酶(AMPactivated protein kinase,AMPK)磷酸化和磷酸烯醇丙酮酸羧激酶(phosphoenolpyruvate carboxykinase,PEPCK)表达。
当磷酸肌醇-3-激酶(phosphatidylinositol 3 kinase,PI3K)与生长因子受体结合后,可激活蛋白激酶B(protein kinase B,Akt)并使其发生磷酸化,抑制下游底物如凋亡相关蛋白B淋巴细胞瘤-2基因相关启动子(Bcl-xL/Bcl-2asociated death promoter,Bad)、半胱氨酸蛋白酶-9(cysteinyl aspartate specific proteinase,caspase-9)活性,调节细胞的增殖、分化、凋亡以及迁移。此外,Akt磷酸化同样会激活IKK蛋白,从而参与调控NF-κB信号通路[57]。Huang等[58]研究表明口服黑米花色苷可显著提高抗凋亡蛋白Bcl-2、Bcl-xL水平,激活胰岛素样生长因子结合蛋白(insulin-like growth factor binding protein,IGFIR)/PI3K/Akt信号通路,抑制链脲佐菌素(streptozotocin,STZ)诱导的1型糖尿病大鼠心脏细胞凋亡。Zheng等[59]证明黑米中C3G可以通过抗氧化应激、抗炎症反应和调节转化生长因子-β1(TGF-β1)/phosphor-Smad2/3通路缓解糖尿病肾病。谷物花色苷作为有效缓解糖尿病发生和发展的食品功能因子,可加大其在人群实验中的研究力度,明确花色苷发挥作用的有效剂量及具体机制。
2.6 对心血管疾病的干预作用
Chen等[60]通过构建STZ诱导的1型糖尿病大鼠模型,发现紫米花色苷提取物抑制Toll样受体4(toll like receptor 4,TLR4)、环氧化酶(cyclooxygenase,COX-2)、IL-6蛋白表达,降低IKK、IκBα、NFκB、丝裂原活化蛋白激酶激酶1/2(mitogen-activated protein kinase kinase,MEK1/2)、MEK5和ERK蛋白磷酸化水平以及心肌肥大和纤维化相关分子标志物表达,改善大鼠心脏功能。Petroni等[61]研究发现紫玉米和红玉米花色苷提取物可有效改善小鼠心脏组织病理学变化,降低阿霉素诱导的心肌细胞毒性。Wang等[62]发现连续6个月给予冠心病患者补充黑米花色苷,受试者血浆中可溶性VCAM-1、可溶性CD40配体(soluble CD40 ligand,sCD40L)和超敏C反应蛋白(high sensitive C-reactive protein,hs-CRP)水平均显著降低。
2.7 神经保护作用
多项研究表明ROS是导致多种神经系统疾病的关键介质,花色苷可穿过血脑屏障,通过调节抗氧化通路对神经组织起到保护作用[63]。Zhang等[24]利用提取的青稞花色苷处理PC12细胞,可有效清除ROS,提高CAT和SOD抗氧化酶活性,抑制细胞凋亡,具有高抗氧化活性和潜在神经保护作用。花色苷可以改善认知能力、增强记忆力,如紫玉米花色苷提取物通过提高SOD和GSH-Px抗氧化酶活性,降低ERK1/2磷酸化水平,发挥神经保护和增强记忆的作用[64]。Lee等[65]的研究表明黑米花色苷可有效改善小鼠记忆和认知功能受损,显著抑制大脑脂质过氧化和NO生成。利用阿尔茨海默症(Alzheimer's disease,AD)小鼠模型和帕金森氏病(Parkinson's disease,PD)小鼠模型,Maria等[66]研究发现小麦花色苷可有效延长记忆消退,减少α-突触核蛋白积累,提高M2小胶质细胞的精氨酸酶1表达。因此,富含花色苷的谷物具有保护神经系统、预防神经退行性疾病的潜在功能。
2.8 对癌症的影响
大量研究表明花色苷可以通过抑制细胞生长和诱导DNA损伤,引起细胞周期停滞,从而抑制多种癌细胞生长、转移[67]。例如Aphisit等[68]发现紫米花色苷提取物可通过降低肿瘤坏死因子α(tumor necrosis factor,TNF-α)、一氧化氮合酶(nitric oxide synthase,iNOS)和NF-κB等促炎因子表达,抑制炎症反应和细胞增殖,预防大鼠肝脏肿瘤发生。Candice等[69]研究证明红玉米和紫玉米花色苷提取物通过上调促凋亡蛋白Bcl-2相关X蛋白(Bcl-2-associated X protein,Bax),下调抗凋亡蛋白Bcl-2表达,降低VEGF水平,从而抑制人结肠癌细胞增殖。Chen等[70]利用黑米花色苷处理乳腺癌细胞,可有效抑制细胞中RAF、MEK和JNK表达,下调基质金属蛋白酶2/9(matrix metalloproteinase 2 and 9,MMP-2/9)分泌,通过靶向作用RAS/RAF/MAPK通路抑制乳腺癌细胞的转移。由于花色苷稳定性差、生物利用度较低,通过研究花色苷改性及靶向递送体系,有望提高人体对花色苷的吸收利用率,并精准靶向作用于癌细胞,从而增强花色苷的抗癌活性。
3 谷物花色苷产品开发与发展趋势
3.1 包装指示剂
对于消费者而言,普通包装材料无法得知食品质量和新鲜度,新鲜度指示器通过与食品中微生物或其代谢产物发生化学反应监测食品的质量[71]。化学合成色素指示剂指示效果好、灵敏度高,但由于其可能存在毒性并影响消费者健康,难以应用到食品工业生产中。花色苷作为天然食品着色剂,具有安全无毒、易获得及水溶性强等特点,在不同酸碱度或金属离子作用下呈不同颜色,对pH变化具有广泛的相应范围。例如Yong等[72]利用生物聚合物添加花色苷开发的颜色指示剂薄膜,基于花色苷的pH变色特性,研制了实时监测食品质量状况的包装指示膜。Wu等[73]将黑米花色苷溶于壳聚糖/氧化甲壳素纳米晶体,制备通过颜色变化检测鱼虾腐败程度的新型智能保鲜膜。此外,Qin等[74]研究发现壳聚糖中添加紫玉米花色苷可提高抗氧化、抗菌活性,成为新型食品智能包装指示材料。但由于缺乏腐败特征物质浓度和种类与颜色变化间的关联机制研究,对花色苷等天然色素的挖掘不足,在一定程度上制约了花色苷在包装指示剂领域的应用。
3.2 食品着色剂
因花色苷具有安全、来源广泛、色泽真实等特性,被广泛用于面包、冰激凌、饮料等食品的着色剂,以其开发的健康食品深受消费者欢迎[75]。彩色玉米富含多种花色苷,可用于不同颜色饮料的开发,其中紫色玉米含有大量酰化和甲基化花色苷,具有较好的稳定性[76-77]。Akogou等[78]的研究表明高粱花色苷在碱性条件下具有较好的色泽和热稳定性,是饮料或其他食品着色剂的优质来源。随着人们健康意识的提高,消费者更加倾向选择采用天然着色剂加工的食品,天然着色剂替代人工合成着色剂也是食品行业的一个重要发展方向。因此,应大力开发挖掘天然、营养、多功能的谷物花色苷着色剂,其在食品产业中应用前景广阔。
3.3 膳食补充剂
花色苷常作为天然色素类功能因子应用于膳食补充剂的开发,如花色苷片剂、胶囊、口服液等[79]。谢洁雯和熊艳珍等[80-81]研究证明黑米花色苷的摄入可以有效改善血脂异常患者的血脂水平,且存在明显的剂量效应关系,膳食花色苷提取物有望成为预防心血管疾病的一个重要措施。此外,花色苷补充剂有效降低肥胖和超重人群体内促炎因子水平,可作为膳食补充剂改善相关人群的慢性炎症[82]。当前市场上主要有越橘花色苷护眼胶囊、富含花青素的护肤品、蓝莓等浆果复合饮料等花色苷产品。对于谷物而言,花色苷主要富含于特定谷物品种(紫玉米、黑米、蓝小麦等),但目前对于谷物来源花色苷产品的开发利用不足,谷物花色苷用于膳食补充剂和功能食品原料的潜力巨大,今后应加强相关功能产品开发的研究。
由于花色苷稳定性较差,且在人体内的生物利用度较低,限制其功能活性及临床应用。提高花色苷稳定性的方法主要包括:a.通过化学修饰提高花色苷对温度和pH条件的耐受性,如酰基化、甲基化等[12];b.利用酚酸等小分子化合物作为辅色剂,提高花色苷稳定性,如咖啡酸、迷迭香酸等[83];c.利用蛋白质、多糖、脂质等天然生物大分子对花色苷进行微胶囊化包埋,如海藻酸钠、乳清蛋白等[84]。制备微胶囊、纳米颗粒、乳液等的递送载体,可大幅提高花色苷稳定性及生物利用度,同时具有较好的生物相容性、安全、无毒等优势,是开发利用花色苷类食品功能因子的主要研究方向之一。例如以海藻酸钠为壁材,以黑米花色苷为芯材制备花色苷微胶囊,实现花色苷在肠道内释放,利于发挥花色苷的功能活性[85-86]。
4 结语
谷物花色苷在食品和医药工业中具有广阔的应用前景,本文综述了不同谷物花色苷的含量及分布,讨论了花色苷的营养干预机制及相关产品的开发利用现状。目前研究表明谷物花色苷通过调控Nrf2、NF-κB、MAPK等信号通路,显著改善糖尿病、心血管疾病、视网膜疾病和癌症等慢性病的发生和发展。基于花色苷物理特性开发的新型智能包装材料、着色剂等,及基于花色苷生物功能活性开发的膳食补充剂已在食品、医药领域广泛应用。因多数花色苷稳定性较差、生物利用率较低,其含量和功能活性极易受加工影响,在一定程度上限制了花色苷在健康食品产业中的应用。未来应进一步研究加工过程中提高花色苷稳定性的方法,通过优化递送系统提高花色苷在体内的稳定性和生物利用度,明确花色苷在体内的关键作用靶点及与肠道菌群的互作机制。
