再谈温血的起源
2022-12-29刘俊
刘俊
现生脊椎动物中内恒温主要出现在鸟类及哺乳动物中。在本刊2016年4期《温血的起源》一文中初步总结了当时的各种证据,讨论了内恒温在两个支系出现的时间。近年来又有了一些新发现,推进了我们的认知。
远古生物的体温没有办法直接测量,我们只有根据一些与内温性密切相关的性状来推测,尤其是可能在化石中保持下来的性状。在现生动物中有隔膜、心脏为四腔、次生腭、鼻腔冷却、直立的姿势及步态、夜行性、体型小型化、高生长速率、特定的骨骼结构(强烈微脉管化、纤维板层型微观结构、较大个体中的哈氏重建)、脑大、亲代抚育等等。
这些性状在过去都曾经用来讨论内温性的起源,尤其是动物的静息代谢率与骨骼生长速度相关,能够反映在骨组织学上。有很多定性推断化石种生长速度的工作。骨骼中的微脉管化是最新揭示的一类结构。骨骼中有大量的毛细血管运输钙磷,在现生四足动物中它们的相对大小、间距和排列可以区分外温和内温动物。相比现代两爬,哺乳类和鸟类的管径小而密。管径与红细胞的大小成比例,内温动物红细胞一般较小,能够更快地运输氧气到组织,满足高的新陈代谢所需。高的有氧运动水平与内温性一般相关,但是一些外温的鱼类也有小的红细胞;红细胞大小与有氧代谢能力相关,但是不一定与基础代谢率有关。三叠纪的主龙类骨骼高度脉管化,脉管小、密且不规则排列,表明快速生长。最近研究表明微脉管化可以直接测量并估计红细胞直径的平均值。
哺乳动物支系内温性的起源时间
早期的古生物学家通常认为二叠、三叠纪的兽孔类已经获得了许多类似哺乳动物的特征。1913年Watson研究Diademodon(一种阔齿兽类犬齿兽类)时认为它有哺乳型的脑、鼻腔有筛鼻甲骨。1931年他将三叠纪一些兽头类及犬齿兽类吻端表面的小坑解释为触须的证据,也是毛的证据。1957年Brink据此提出既然特化的毛发在这个时候出现,一般的毛发应该起源更早。1960年Van Valen提出兽孔类已经是内温动物。
2011年在南非二叠纪晚期的粪化石中报道了疑是毛发的结构,2016年从俄罗斯的粪化石中也报道了疑是毛发的结构。不过目前还没有发现确切无疑的毛发特征,如黑素小体。但是根据头部神经系统的研究,Benoit等人2016年提出眶下管的存在才表明上颌触须的存在,他们提出毛发在大约2.46—2.40亿年前出现。目前最早的确切的毛发化石发现于中国中侏罗世。
在现代蜥蜴中,实验发现它们不能同时行走和呼吸。在演化中,横隔膜的出现解决了轴向肌肉功能的冲突。而横隔膜的出现可以从肋骨的形态推测,一般认为与背椎分化为胸椎与腰椎一致。有研究者认为横隔膜出现很早,而最近研究认为最关键阶段发生在犬齿兽类中,动物脊柱的侧向运动转换为上下的弯曲,取得了直立的步态。这大约发生在二叠纪晚期到三叠纪早期。
次生腭的出现隔开了口和鼻,动物在进食的同时可以呼吸。在三叠纪早期的三尖叉齿兽Thrinaxodon已经出现完整的次生腭,计算机断层(CT)扫描发现它上颌骨的内侧具有鼻甲脊,表明有鼻甲软骨。而后期的犬齿兽类则有骨化的筛鼻甲。Crompton认为早期鼻甲的功能可能是散热而不是保持热量和水。其他人的研究也肯定鼻甲对哺乳型的内温的重要性。
许多兽孔类具有微脉管化,哺乳类特征的红细胞大小在三叠纪早期已经出现,落在现代袋鼠和有胎盘类的区间。这支持有氧代谢效率在这一时期大大提高。
亲代抚育可能与内温起源密切相关,不过这种行为在四足动物中广泛分布,它们可能是后者的条件。夜行性也通常认为与内温性相关,也被认为与哺乳类起源有关。不过根据对下孔类巩膜环及眼眶大小的研究,暗视觉在盘龙类中就已经出现,比哺乳起源早了近1亿年。根据氧同位素的方法,发现三叠纪时某些二齿兽类以及真犬齿兽类的体温可能显著高于环境温度,是内温动物。
近年来的CT扫描,增进了对兽孔类的脑部演化的认识。研究发现颅内模大小及形态在早期兽孔类中很多样化。生物学家早已经注意到演化中脑的相对大小增加的趋势,而且脑相对容量与新陈代谢率密切相关。2014年Gillooly和McCoy根据现生动物建立了脑的相对容量与体温(T)的回归方程:
ln(体重校正后的相对脑容量)=-0.96(1/kT)+ ln(37.2),k为玻尔兹曼常数(=8.62×10-5 eV/ K)
2016年Benoit等人依据这个关系,使用不同化石属种的体重、脑大小的估算计算出兽孔类的体温。在Probainognathia一支中巴西兽体温达到28°C,而哺乳型类体温均高于30°C。同时有的二齿兽类体温高达34°C。不过最近有研究表明脑容量与体重没有稳定的异速生长关系,可能脑的相对容量需要更精细的方法来表达。所以目前的结果可能存在问题,脑容量在Probainognathia一支中的增长模式还需要仔细研究。
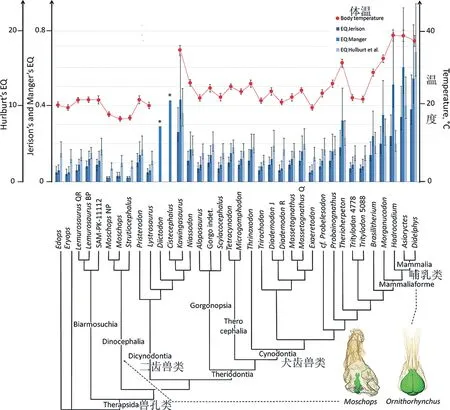
兽孔类中脑量指数(EQ,脑容积相对于体重)以及体温(红色)的演化
Benton(2021)综合了犬齿兽类的骨组织学信息、上颌神经、鼻甲、红细胞大小等建立一个指数推测内温程度,支持在晚三叠世哺乳型类出现前后可能已经是内温性了。
最近研究人员在内耳形态找到了与体温的联系。内耳的半规管是平衡器官,半规管内的内淋巴液的粘性对于其功能至关重要。由于内淋巴液的粘度随着体温的变化而变化,半规管要正常运行,需要改变几何形态适应体温变化所带来的新粘度。于是半规管不仅用来预测运动能力,还用来推断体温。
他们构建了一个热运动性指数(TMI),源自于半规管系统的测量参数,并进行异速生长修正。通过对现生200多个脊椎动物物种的分析,他们建立了TMI与体温之间关系的数学模型。虽然对于物种级别没有较好的相关性,但是对于支系来说相关性很好,能够解释80%的变化。从化石中无法得知内耳的膜迷路结构,内淋巴液的粘性、化学成分等信息。这一研究假设内淋巴在哺乳类支系演化中没有发生变化。他们重建了64个化石种的TMI,其中下孔类的各支系的TMI表示在23页图中,并利用这一关系来推测其体温。在物种级别其结果是三列齿兽为内温动物,巴西兽内温概率是0.46,而大多数早期哺乳型类可能是内温动物,但巨颅兽(Hadrocodium)是外温动物。不过单个物种的推测可信度不高。哺乳形类为内温动物的概率高达0.7,这一支系的起源时间大约是2.33亿年前,卡尼期洪积事件期间。推测向高体温转换主要发生在Probainognathia这一支,而且是在不到一百万年时间内完成的。不过文章中这个阶段的化石数据太少(3个),没有清楚阐述这一重要过程。
不过也有观点认为哺乳动物支系的内温性出现得很晚,至少在早期的哺乳型类还没有获得与现生哺乳动物一样高的新陈代谢率。根据2020年Newham等人用同步辐射扫描摩根齿兽及许氏兽牙齿,得到牙骨质部分的生长轮,发现摩根齿兽可以活14年,而许氏兽可以活9年。这一寿命远比相似大小的现生哺乳动物长得多,而与爬行动物的相似,估计其基础代谢率较低。摩根齿兽股骨中营养孔面积表明其血液流动速率位于现生爬行动物与哺乳动物之间,暗示其最大代谢率已经增大。他们认为基础代谢率增加发生在中侏罗世,多瘤齿兽与哺乳动物冠群的共同祖先出现时,也是最早确切毛皮出现的时间。
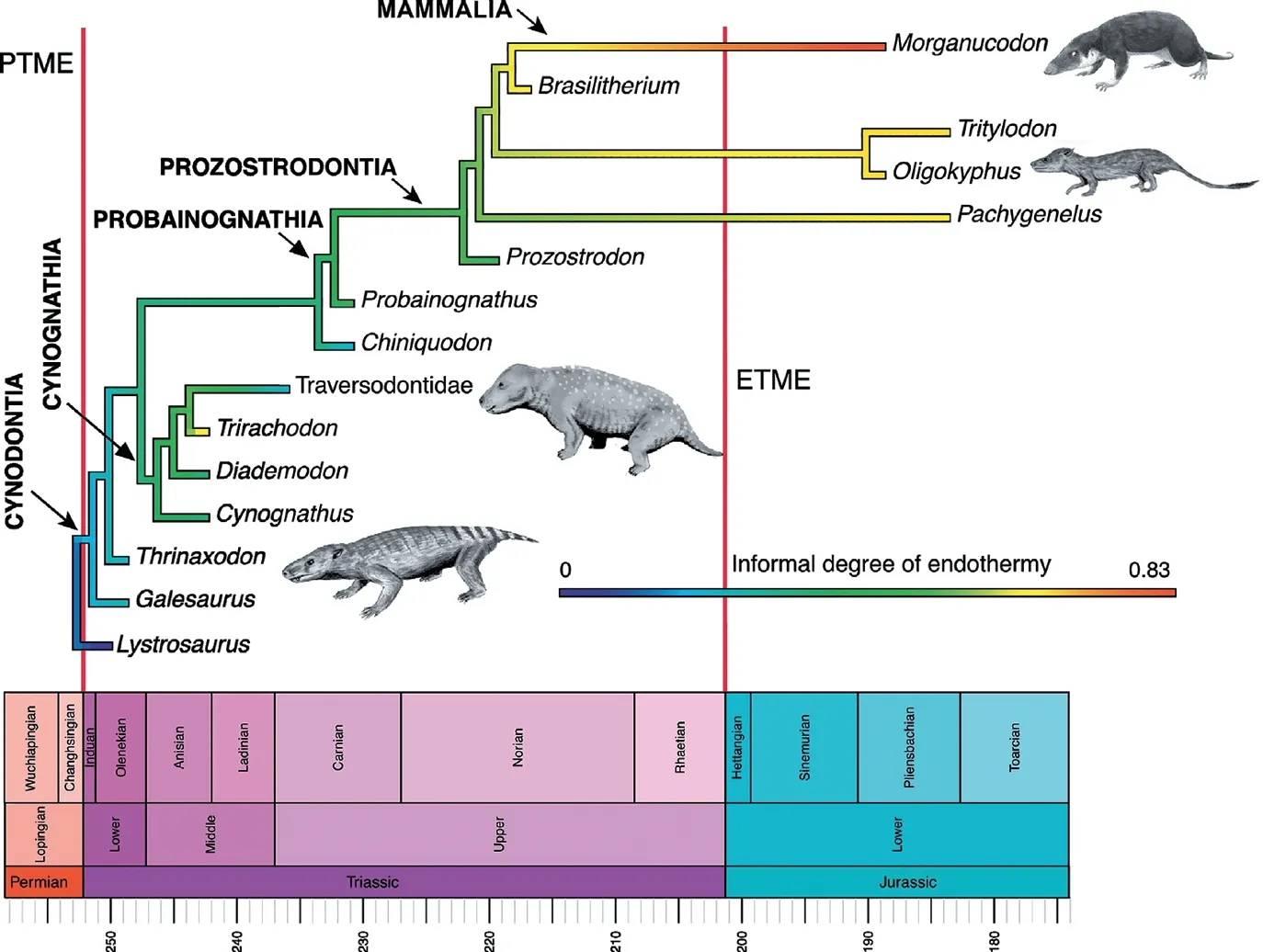
下孔类内温性的演化
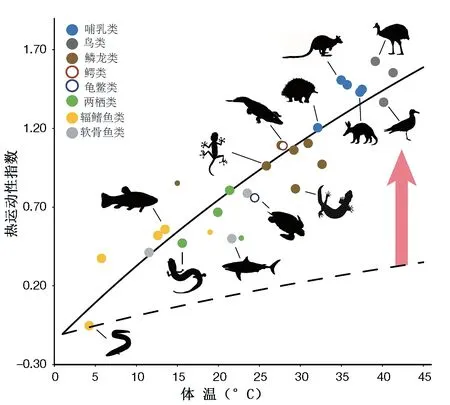
热运动性指数与体温的关系图,虚线表示体温仅影响内淋巴液的粘性而不是行为活动
鸟类支系内温性的起源时间
与哺乳类相反,鸟类祖先恐龙传统上被认为是外温动物。过去认为可能有些恐龙是内温动物,但是最近研究暗示这一支系内温性起源可能很早,在非恐龙的主龙型类就出现了,或者是多次独立演化。
Legendre等人(2016)使用了一套统计模型根据骨组织学特征重建化石种的新陈代谢率。他们的结果表明中生代兽脚类恐龙新陈代谢率与现代鸟类相当,在早期主龙类中新陈代谢率就已经升高。
鸟臀类恐龙大多具有鳞片,但是在天宇龙、鹦鹉嘴龙中发现单丝状的皮肤衍生物;故有的祖先状态重建结果认为恐龙的最近共同祖先有丝状的皮肤衍生物。从近两百年前被发现开始,翼龙就被认为是内温动物,发现了许多带“毛”的物种。不过直到20世纪80年代才意识到翼龙是恐龙的姊妹群,这样翼龙与恐龙的共同祖先很可能带“毛”。
2020年通过对恐龙的蛋壳进行团簇同位素分析,报道了兽脚类(伤齿龙)体温为27±4°C,28±3°C以及38±4°C,鸟臀类(慈母龙)为44±2°C,而另外一个罗马尼亚的蛋壳(蜥脚类或者鸟臀类)得出36±1°C。这些温度在现生内温动物体温范围内,远超出从同一地点的软体动物壳得出的环境温度(25±1°C和28±2°C)。这揭示恐龙具有高的体温是这一类群的祖先状态。
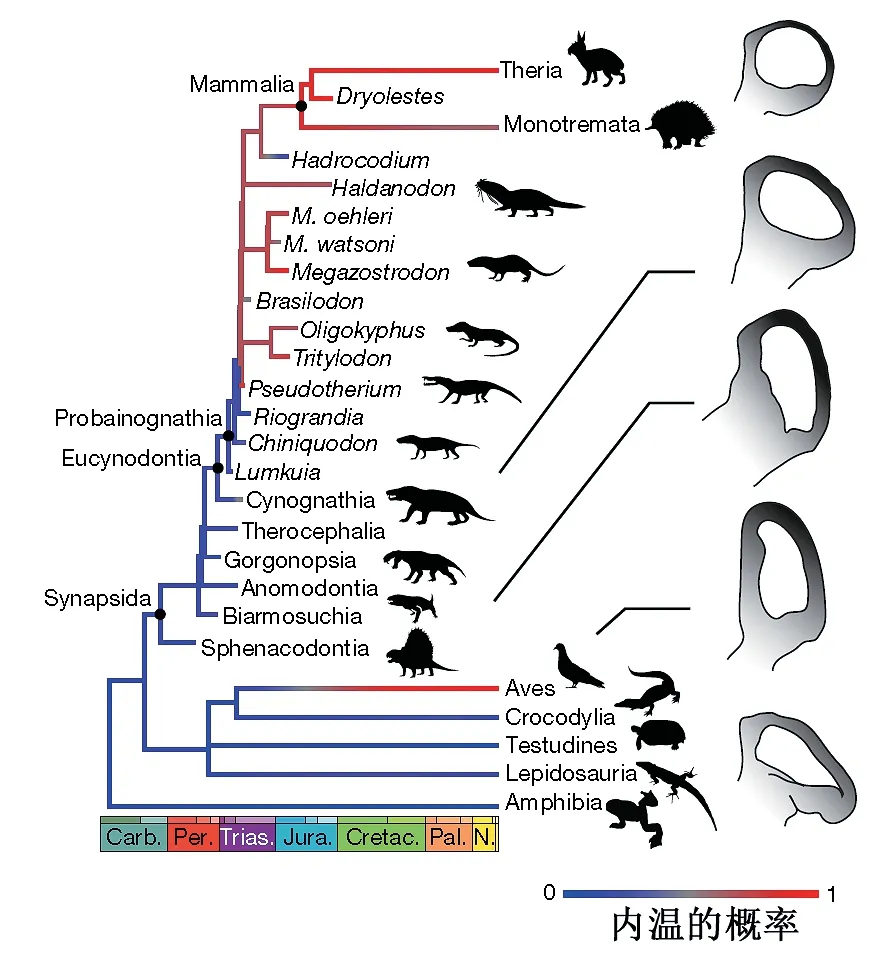
四足动物热运动性指数的系统发育分布
鳄鱼心脏也有四腔,但是具有两条主动脉,它能够控制心脏血液的流向。研究者提出三叠纪的鳄鱼祖先是内温动物,只是因为适应水生的需要后来才转回外温性。骨组织学研究也支持这一观点。如果接受鳄鱼三叠纪祖先早期也是内温性,那么可以推测恐龙和鳄鱼的共同祖先,三叠纪早期的主龙类已经是内温性。
由于下孔类支系及主龙类支系内温起源都可能是三叠纪早期,有人提出当时的大气氧含量降低可能是驱动因素,因为有氧代谢率取决于心血管系统传输氧气到身体各组织的效率。低的大气氧含量将促使动物改变肺的形态和容量、产生四腔的心脏、获得低肺血压和高体循环血压、高血流速度、增加毛细血管表面积、减小红细胞以增大表面积利于气体交换。低的大气氧含量可能还促进四足动物的直立姿态,因为它们能够在运动时呼吸。早三叠世大气氧含量确实从二叠纪晚期的33%降低到19%,到中三叠世又回升到22%。不过这一含量与今天地球(21%)相当,可能并不会有如此强烈的选择压力。到三叠纪晚期大气氧含量可能只有11%-15%,这可能是恐龙中气腔化增多的原因。
根据研究,大型侏罗纪、白垩纪的鱼龙和鳍龙也独立获得了高而恒定的体温。现代鱼类也有内恒温。这可能都表明内恒温起源没有那么难,应该是多次独立起源事件。
目前关于四足动物内温性起源还有很多有待解决的问题。首先是能否用单一的特征,例如新陈代谢率,将内温动物与外温动物截然区分开来?如果二者间是渐变的,中间包括哪些阶段,这些阶段能否用可以从化石和地质记录中获取的特征来识别?从外温性到内温性,演化中下列事件的发生先后究竟如何:近矢面的步态,呼吸系统的改善,心血管系统加强,生理产热,脑量增加以及亲代抚育?主龙类和下孔类内温性的起源是特定的外界变化促发的吗,它们演化同时吗,关键驱动因素究竟是什么?
