论杂种优势机理的逻辑体系
2022-12-28潘春桂
潘春桂
(安徽省东至县农业农村局,安徽 池州 247210)
揭示杂种优势的产生机理,无论是对于遗传学本身学科的发展,还是对于在实践中提高杂种优势利用效率,增加经济效益都具有极其重要的意义。到目前为止,关于杂种优势机理学说和研究方法的多样性并未澄清其发生机理[1],研究者均寄希望于遗传学等学科的进一步发展。很少看到对杂种优势必然性的哲学思考。目前主要静态地研究杂种一代与双亲的差异,对外部条件的作用机制几乎很少研究。如果假设杂种优势初始值为零,它能够和外部条件相互作用发生转化吗?影响杂种优势的外部条件很多,环境养分供给可能是最重要及最易受控的条件。潘春桂于2020年提出了一种植物养分获取总量杂种优势模型[2],本文在这个模型的基础上,对不同的养分环境供给量之下杂种优势的变化进行分析,以期获得有益的启示,希望引发对杂种优势机理逻辑体系的思考。
1 模型与分析
假设环境中各种养分供给量是一定的,植株个体存在这样一种养分吸收过程,吸收过程可分为两个阶段:固定速度吸收期(饱和期)及停止吸收期(停止期),各种养分停止吸收时刻或转折点由最先进入自身临界点的养分所决定,将这种相对亏缺的养分称为主导养分。这里认为不同养分临界环境消耗率在同一遗传类型中相同,杂种一代为加性遗传,饱和期各种养分吸收速度也为加性遗传。在某一时刻任一遗传类型养分获取总量由各种养分吸收速度大小及它们进入自身临界点同步性所决定。
先简单介绍养分的获取过程,首先设环境三种养分A、B、C总供给(vt,质量单位)分别为5、4、3,当任一种养分临界消耗率为K时,三种养分临界消耗量(vt)分别为5 K、4 K、3 K。对于P1,三种养分A、B、C的吸收速度(v,质量时间比)分别为3、2、1,临界消耗率为100%时,三种养分临界消耗量(vt)分别为5、4、3,经过时间1.66(t,时间单位)时,A、B、C三种养分消耗量(vt)分别为5.00、3.32、1.66,可看出A养分已达到临界消耗量,1.66(t)就是转折点,可称A养分为主导养分。这时个体停止对各种养分的吸收,即由饱和期进入停止期,养分吸收总量停止在5.00+3.32+1.66=10(vt)上。对于P2,养分临界消耗率为50%,则三种养分临界消耗量(vt)分别为2.5、2、1.5,它们吸收速度(v)分别为2、3、1,经过0.66(t)时,三种养分消耗量(vt)分别为1.32、2、0.66,养分B达到临界消耗量,总吸收量停止在(2+3+1)×0.66=4.00(vt),0.66(t)就是转折点(时刻),可称B养分为主导养分,当然转折点之前养分吸收总量为三种养分吸收速度之和与时间乘积。对于杂种一代P1×P2,养分临界消耗率为加性遗传,为75%,三种养分临界消耗量(vt)分别为3.75、3.00、2.25,各种养分饱和期吸收速度也为加性遗传,则三种养分A、B、C吸收速度(v)分别为2.5、2.5、1,在1.2(t)时,养分B达到临界点,1.2(t)就是转折点,可称B养分为主导养分,总吸收量停止在(2.5+2.5+1)×1.2=7.2(vt)。如果在2(t)时进行(取样)计算,以上组合的总养分获取量离亲值为0.20(vt),中亲优势为2.85%。先将各种养分吸收速度的衰减定义为各种养分速度相对于其饱和速度的减小比例,再从养分吸收速度衰减角度看看2(t)时杂种优势是如何形成的。给出两种定义,对于每种遗传类型,(1)固有衰减:每种养分的吸收速度按照本身所处状态(由饱和吸收速度、自身临界消耗量等决定)的衰减;(2)实际衰减:每种养分被迫与固有衰减最大的养分(主导养分)同步衰减。例如,在2(t)时,P1对A养分吸收速度的固有衰减为(1-1.666/2)×100%= 16.7%,对于B、C两种养分,仍处于饱和期,固有衰减均为0.0。因此P1对三种养分实际衰减均为16.7%。再看对A养分吸收速度中亲值固有衰减为{1-(3×1.666/2+2×1.25/2)×0.5/[(3+2)× 0.5]}× 100%=25.0%,对A养分吸收速度中亲值实际衰减为{1-(3×1.666/2+2×0.666/2)×0.5/[(3+2)× 0.5]}×100%=36.7%。2(t)时各种养分吸收速度固有及实际衰减,详见表1。
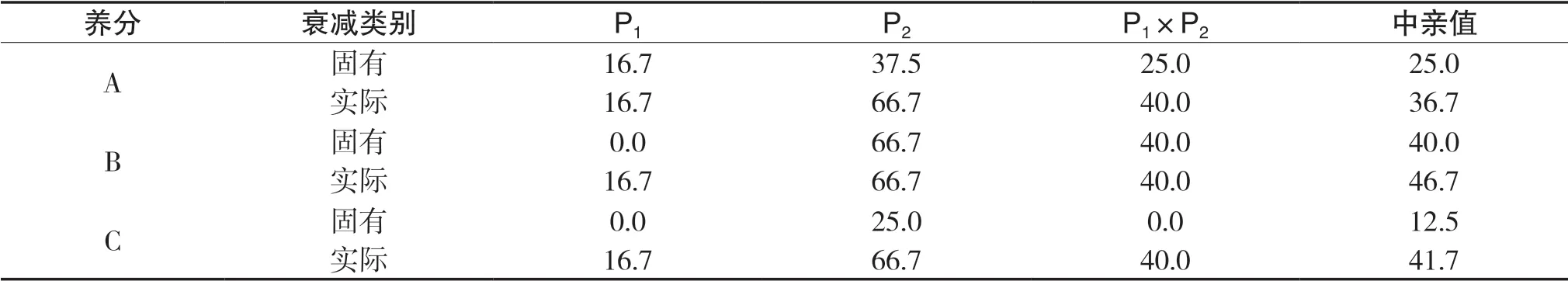
表1 2(t)时各种养分吸收速度固有及实际衰减 单位:%
由以上分析可以推论,当双亲主导养分不同时,则可能导致每种养分吸收速度中亲值的实际衰减程度大于自身固有衰减,而中亲值固有衰减可认为大致等于杂种一代的固有衰减;如果杂种一代各种养分进入自身临界点同步性好或完全同步,各种养分吸收速度的固有衰减程度相近或相同,则杂种一代每种养分固有衰减等同于实际衰减。即可得出结论:每种养分吸收速度中亲值的实际衰减程度大于杂种一代实际衰减。这有利于获得较大的杂种优势。当双亲的主导养分为同一养分时,这使杂种一代各种养分进入自身临界点难以或不能达到同步,各种养分中亲值与杂种一代的实际衰减相近,自然不利于杂种优势的形成。给予简单证明,设三种养分供给量为M、N、G,P1三种养分饱和吸收速度为a1、b1、c1,P2三种养分饱和吸收速度为a2、b2、c2,根据 条 件,M/a1<N/b1、M/a1<G/c1及M/a2<N/b2、 M/a2<G/c2,则可得:2 M/(a1+a2)<2 N/(b1+b2)、2 M/(a1+a2)<2 G/(c1+c2),即杂种一代不能达到同步性:2 M/(a1+a2)=2 N/(b1+b2)=2 G/(c1+c2)。可看出同双亲一样,杂种一代第一种养分也为主导养分,其吸收速度固有衰减介于双亲之间,则杂种一代每种养分实际衰减也介于双亲之间,在很多情况下,这不利于形成较大的总养分获得量离亲值。当双亲临界消耗率不相等时,也存在同以上相似的结论,只需初等数学就能证明(此处省略)。容易看出,对中亲优势的负效应的不确定性有所增加。2(t)时P1×P2在不同养分供给条件下的总养分获得量的杂种优势详见表2。从下文可以看到,当某一种养分严重短缺时可形成这种现象,这直接证明本文结论的正确性。将主导养分在双亲中是否相同,杂种优势大小存在差异的现象称为主导养分效应。
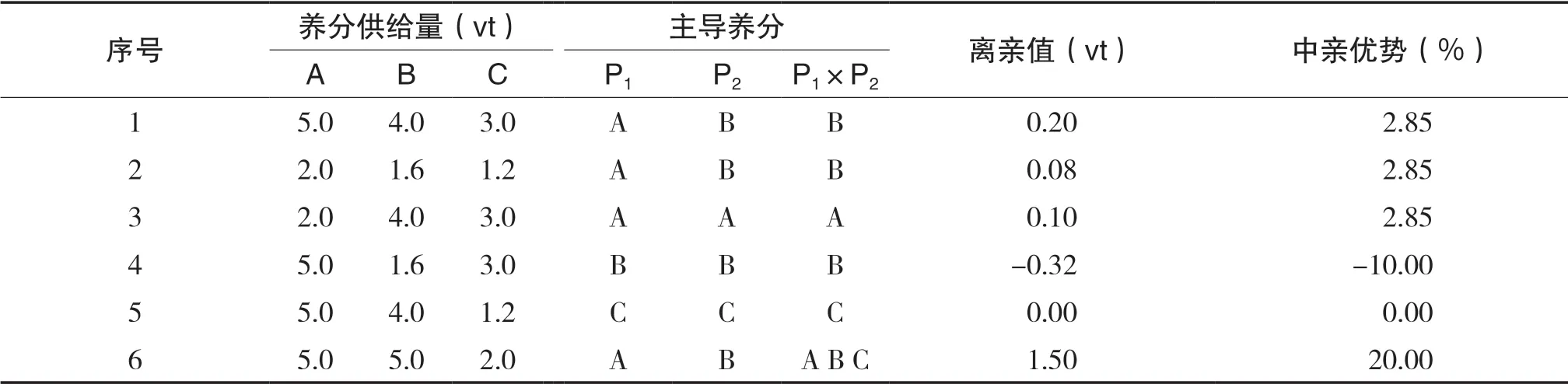
表2 2(t)时P1×P2在不同养分供给条件下的总养分获得量的杂种优势
如果保持双亲各种养分饱和吸收速度及临界消耗率如上文一样,只改变养分供给量,从表2可看出,在第2种养分条件下,各种养分均减少了60%,离亲值亦减小60%,中亲优势没有改变。在第3、4、5养分条件下,分别只有一种不同养分减小了60%,可见这种减少的养分均为双亲的主导养分,这时离亲值均显著减小,中亲优势也多有减小。在第6种养分条件下,养分供给总量未变,只是B、C两种养分量互有增减,杂种一代各种养分进入自身临界点完全同步,这时获得了较大的离亲值与中亲优势。
2 讨论和结论
大量试验表明,单一缺乏某养分如缺氮、缺磷、缺钾、缺水时,中亲优势或离亲值近似为零[3-7]或显著减小[8-13],这与上文分析的结论是一致的。当环境中某种养分严重短缺时,则可能这种养分对于双亲同样属于相对性缺乏或同为主导养分,可导致单株养分获取总量离亲值显著减小,从而引发生物量或产量的离亲值显著减小,这是由于两者之间存在内在紧密的联系,在不同作物的杂交试验中,均存在类似的结论。这表明,主导养分效应在杂种优势机理中占有重要的地位,这必将引发对目前杂种优势机理研究方法的重新审视或反思。可以设想,当环境中所有养分不足时比仅一种养分不足时可能会产生较大的离亲值或中亲优势[14,15],因为后者双亲主导养分可能相同。非常有必要通过试验对这种现象进行进一步验证。
人类认识到杂种优势现象已有近两个世纪,一直以来,科学家提出了各种假设试图解释其形成机制。近年来由于分子遗传学的快速发展,已经在多个层面上获得了进展[16-18]。目前的研究方法是基于这样的一种逻辑体系,由于杂种一代是双亲两套遗传物质的结合,不存在新的遗传物质,因此杂种优势必定来源于它们的相互作用,这导致了杂种一代获得了有别于双亲的分子生物学或遗传学的特异性表现或过程[19,20]。但这种方法已面临了一个窘境,即当外部试验条件改变时,结论可能不一致。另外一点,科学家可能有意无意忽略了这样一个问题,杂种一代这种内在的、微观的特异性是否属于一种本能的或自然的生命过程,也可能如杂种一代外在的、可视的表现一样,如具有较大的营养体或旺盛的生长势,也是杂种优势的一种表现,而不是其形成原因。因为已经有研究表明,环境效应可诱导出基因表达的差异性[21]。从上文模型与分析可看出,表面上杂种一代与双亲是处在相同的外部条件之下,如果这些条件具备某些特征,便能够使杂种优势完成了从无到有的转化。在模型中并没有预设杂种一代具有区别于双亲的特异性,这种特异性可有利于杂种优势的形成。而是仅假设了各种养分饱和吸收速度及临界环境消耗率均为加性遗传。既然最终形成了总养分获得量的杂种优势,必然反过来可能使杂种一代发生内在分子生物学或遗传学的改 变[22],形成某种内在特异性,可见这种内在特异性并不是本能或自然形成的,是与外界条件相互作用的结果,并不能认为它是杂种优势产生的原因,它与杂种一代外在表现特异性处于同等地位。简单地说,当这种外部环境条件对杂种一代相对有利时,便产生的杂种优势。可见目前研究的逻辑体系存在很大的局限性。上文模型与分析重建了一种全新的逻辑体系,包括两个方面:第一,不存在必然的杂种优势,即使存在,也属于偶然性。第二,杂种优势主要是在外界条件作用下转化形成的。因此研究的重点应着眼于转化的条件及过程,而不是像以往那样,主要针对于杂种一代与双亲的差异性。由于试验要素中已包含了外部条件,也避免了像以往那样,不同条件下,结论可能存在差异。虽然植物生物量大多来自于光合产物,但与植株获得的养分存在显著的正相关关系[23]。本文简单地以环境养分供给量作为试验条件,并假定各种养分饱和吸收速度为加性遗传,由于环境中养分消耗或不足,当杂种一代各种养分吸收速度相对于其饱和速度的衰减如果小于中亲值或双亲的相应衰减,便产生了杂种优势。目前形成的共识是,杂种优势机理问题在现阶段科学发展水平之下是不能得到解决[24],只有对未来有所期待。
